
Abstract
Background:
The procancer effect of TEA domain transcription factor 4 (TEAD4) has been gradually discovered. However, its expression in esophageal cancer (EC) cells and its effect on proliferation and apoptosis have not been reported. In this study, we investigated the possible role of TEAD4 in EC cells.
Materials and Methods:
TEAD4 messenger RNA and protein expression were assessed in EC cell lines by real-time quantitative-PCR and Western blot. Gene silencing approach was employed to investigate the potential role of TEAD4 in cellular growth, proliferation, migration, and invasion in EC cells. The interaction between TEAD4 and transcription factor 7 (TCF7) was verified by co-immunoprecipitation reaction. The cell apoptosis rates of KYSE-30 cells were detected by flow cytometry. Meanwhile, the expression of apoptosis-related proteins in KYSE-30 cells was detected by Western blot analysis.
Results:
TEAD4 was significantly increased in EC cell lines, interference of TEAD4 inhibited EC cell viability, invasion, and migration, and promotes apoptosis. TCF7 was found when using STRING website to interact with TEAD4 proteins and TCF7 was significantly increased in EC and knockdown expression of TEAD4 hindered biological function of KYSE-30 cells and this effect was reversed by overexpression of TCF7.
Conclusions:
The findings concluded that TEAD4 is highly expressed in EC cells and gene silencing of TEAD4 inhibits proliferation and promotes apoptosis of EC cells by regulating TCF7. These findings suggested that TEAD4 might be a novel therapeutic target for the prevention of EC.
Introduction
Esophageal cancer (EC) is one of the most common malignant tumors worldwide. 1 At present, the rate of EC is declining, but in many economic developing countries, the mortality rate of EC ranks second in tumor mortality, which seriously affects human health. 2 In recent years, with the continuous improvement of new chemotherapy drugs and other treatment levels, the survival rate of EC has improved. However, due to the strong metastatic and invasive ability of EC and the high recurrence rate, the treatment effect of EC is not very satisfactory, and the 5-year survival rate is only 15% to 35%. 3,4 Therefore, the research on EC-related transcription factors and expression genes is of great significance to elucidate the therapeutic targets of EC and provide theoretical basis for the treatment of EC.
Members of the Hippo pathway effector protein-TEAD protein family are highly conserved in mammals, with four members, including TEAD1, TEAD2, TEAD3, and TEAD4. 5 Common features of the family members are the inclusion of the TEA domain (DNA-binding domain) and transcriptional activation domain capable of binding to transcriptional activators. 6 The TEAD protein family, with the participation of cotranscription activators, can transmit signals downstream, affecting cell differentiation, proliferation, antiapoptosis, and stem cell maintenance. 7 Among them, TEAD4 can participate in the process of tumorigenesis and development by activating the transcriptional expression of proto-oncogenes such as CTGF, AXL, and Myc. 8 Recent studies have shown that TEAD4 plays an extremely important role in the development of many malignant tumors, such as ovarian cancer, breast cancer, and prostate cancer. 8,9 In this study, the expression of TEAD4 in EC was analyzed and the role of TEAD4 in proliferation, migration, invasion, and apoptosis was assessed in EC.
Materials and Methods
Cell culture and cell transfection
Human esophageal cancer cells (KYSE-30, KYSE-70, EC-9706, ECA109, and TE-1), as well as human esophageal epithelial cells (HEECs), were purchased from the Stem Cell Bank, Chinese Academy of Sciences in Shanghai, China. All cell lines were cultured in RPMI-1640 medium (Thermo Fisher Scientific, Inc.), which contained 10% fetal bovine serum (FBS) and cultured at 37°C with 5% CO2. Appropriate short hairpin RNA negative control (shRNA-NC) and shRNA targeting TEAD4 were transfected into KYSE-30 cells. The KYSE-30 cells were plated in 24-well plates and transfection was conducted when the cells reached 70%–80% confluence. Transfection was carried out using Opti-MEM, serum-free RMPI, and Lipofectamine 2000 according to the manufacturer's instruction (Invitrogen, Carlsbad, CA). The cells were cultured for 24–72 h at 37°C after transfection.
Colony formation assay
KYSE-30 cells at logarithmic growth phase were trypsinized to passage for forming a single-cell suspension followed by count on a hemocytometer plate and culture in a six-well plate with 400 cells per well. The culture was continued for 72 h, and the medium was changed every 1–2 d, and the formation of the cell clone was observed at any time. After 72 h, the medium was discarded, washed once with phosphate-buffered saline (PBS), and the filter paper was washed to remove excess liquid and air dried with a ventilated place. After staining with 0.1% crystal violet methanol solution to 20 min, PBS was washed, and after air drying, the number of cell clones of >50 cells per well was recorded under a microscope.
Cell Counting Kit-8 assay
At 48 h post-transfection, the KYSE-30 cells were plated in a 96-well plate at a concentration of 1 × 103 cells/mL per well, respectively. The cells were treated as shRNA-NC group and shRNA-TEAD4 group. For cell proliferation assay, 10 μL of Cell Counting Kit-8 (CCK-8) solution was added to each well for 2 h. The absorbance was detected by a microplate reader at 450 nm.
Wound healing
The cell layer of transfected KYSE-30 cells growing to 80% was scratched. Cell migration was monitored and photographed using a microscope after 24 h. Using 10 μL sterile suction head, two straight lines were drawn perpendicular to the marking line in each hole to form scratches, and the exfoliated cells were washed off with serum-free medium. One milliliter of RPMI1640 medium without 10% FBS was added into each pore. Photographs were taken under the microscope, the recording time was t = 0 h, and the scratch width was recorded as W0. The cells were cultured for 24 h in a 5% CO2 incubator at 37°C and then photographed again to record the cell migration for 24 h. At this time, the scratch width was recorded as W24. The ability to migrate was assessed by measuring changes in the size of the injured area in the six areas.
Migration rate = (W24−W0)/W0 × 100%.
Transwell
Cell invasion was detected by Transwell assay. These cells were cultured with the serum-free medium for 12 h before the formal experiment. Next, the matrixgel (BD, USA) was added into the 8 μm boyden chamber. Then, the chamber was placed into the incubator for 4 h to confirm the solidification of matrixgel. The cells were placed into the upper chamber with 1 × 104 cells per well and cultured in 400 μL serum-free RPMI-1640 medium. To the lower chamber was added 600 mL RPMI-1640 with 10% FBS. After incubation for 24 h, cells on the bottom of the upper chamber were fixed with 90% ethanol solution for 30 min. Then, 0.1% crystal violet was used to stain the cells for 10 min. At last, the invasion of cells was observed by a light microscope.
Flow cytometric analysis of apoptosis
KYSE-30 cells were harvested and fixed in precooled ethanol, then KYSE-30 cells were resuspended in cold buffer with 5 μL of Annexin V-FITC for 10 min at room temperature. The samples were incubated for 5 min after adding 5 μL of PI and 200 μL of binding buffer. Cell apoptosis was analyzed using Annexin FITC/PI flow cytometry assay kit.
Coimmunoprecipitation
Cells were harvested 24–48 h after transfection, and cell lysis buffer was added and lysed on ice for 30 min. Then, 1 μg of the corresponding antibody was added to the cell lysate and incubated at 4°C overnight; 10 μL of protein A agarose beads was taken, washed three times with an appropriate amount of lysis buffer. Antibodies were incubated overnight in cell lysates at 4°C with slow shaking for 2–4 h. After the immunoprecipitation reaction, the agarose beads were centrifuged to the bottom of the tube. The agarose beads were washed 3–4 times with 1 mL of lysis buffer; finally, 15 μL of 2*SDS loading buffer was added and boiled in boiling water for 5 min. Bound proteins were separated with SDS-PAGE, followed by visualization using Western blotting.
Statistical analysis
All data of this study are expressed as the mean ± standard deviation and repeated at least three times. SPSS 17.0 software was used to conduct all statistical analyses (SPSS, Inc.). A one-way analysis of variance followed by a Tukey's or Dunnett's test was performed using GraphPad Prism 5 software (GraphPad Software, Inc.). Differences with p < 0.05 were significant.
Results
TEAD4 was significantly increased in EC cell lines
The protein and mRNA expression of TEAD4 in EC cell lines (KYSE-30, KYSE-70, EC-9706, ECA109, and TE-1) and in HEEC was detected by Western blot and real-time quantitative-PCR. TEAD4 expression level was significantly increased in the EC cell lines compared with the HEEC, especially in KYSE-30 cells (Fig. 1A, B).

Interference of TEAD4 inhibited the cell proliferation in EC cell lines. TEAD4 was significantly upregulated in EC cell lines as assessed by RT-qPCR
Interference of TEAD4 inhibited the biological function
To further investigate the role of TEAD4 in EC cells, the interference efficacy of shRNA-TEAD4 in KYSE-30 cells was first confirmed, RT-qPCR demonstrated that the inhibitory effect was improved (Fig. 1C). For CCK-8 assay, cell viability of KYSE-30 cells transfected with shRNA-TEAD4 was significantly inhibited compared with the negative control (Fig. 1D). Furthermore, the colony formation assay revealed results similar to CCK-8 (Fig. 1E). Then the role of TEAD4 in invasion and migration of EC cells by transwell invasion test and cell scratch test was studied. As shown in Figure 2A–D, the invasiveness and migration ability of KYSE-30 cells transfected with shRNA-TEAD4 were significantly decreased.
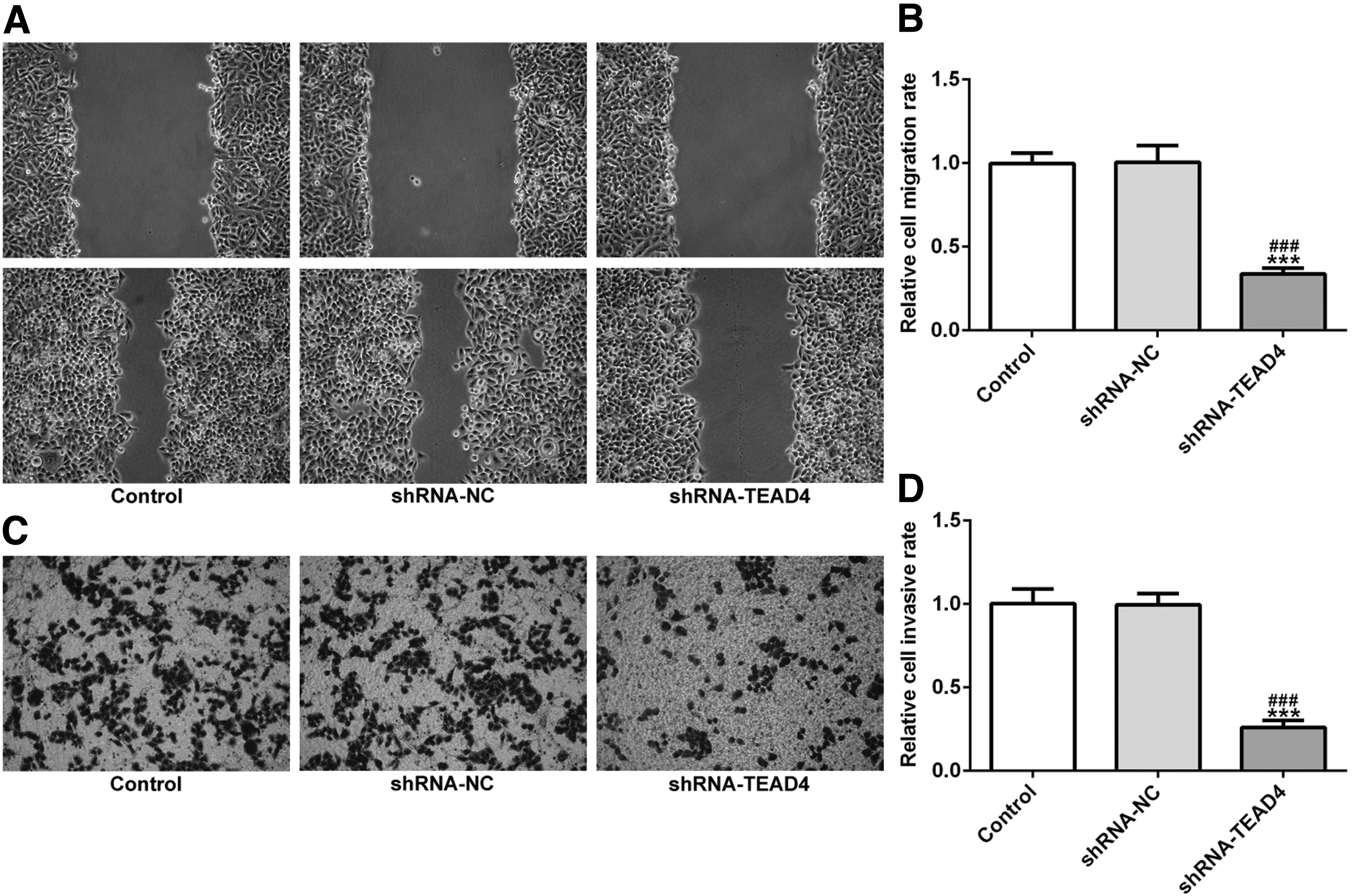
Interference of TEAD4 inhibited the cell invasion and migration in EC cell lines. Wound-healing assays
Knockdown expression of TEAD4 promotes apoptosis in KYSE-30 cells
Flow cytometry was used to detect the apoptosis rate of EC cells and to investigate the mechanism of TEAD4 in KYSE-30 cells. As shown in Figure 3A, the apoptosis rate of KYSE-30 cells was significantly increased after TEAD4 knockout. Western blot results also showed that after TEAD4 knockout, BCL-2 was significantly decreased, and the expression levels of Bax, PARP, and caspase-3 in KYSE-30 cells were increased, which was consistent with the results of apoptosis detection.

Interference of TEAD4 promotes apoptosis in EC cell lines.
Knockdown expression of TEAD4 downregulates the expression of transcription factor 7 in EC cell lines
First, transcription factor 7 (TCF7) was found to significantly increase in EC cell lines (Fig. 4A, B). RT-qPCR was performed to further investigate whether TEAD4 could regulate the expression of TCF7. In cells treated with shRNA-TEAD4, the expression level of TCF7 decreased compared with shRNA-NC (Fig. 4C). Coimmunoprecipitation analysis revealed protein–protein interaction between TEAD4 and TCF7, indicating that TEAD4 was associated with TCF7 (Fig. 4D).
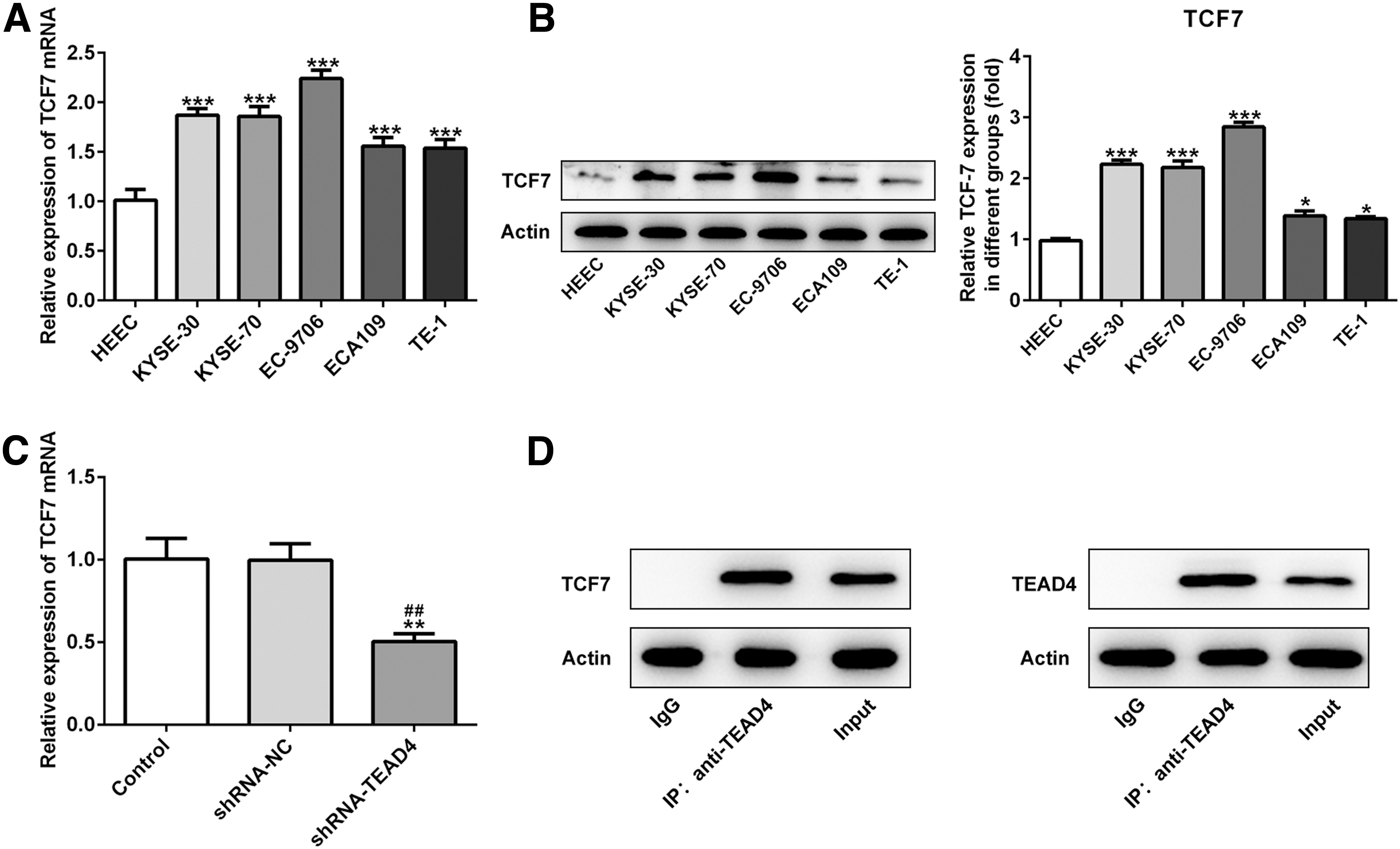
Interference of TEAD4 downregulates the expression of TCF7 in EC cell lines. TCF7 was significantly upregulated in EC cell lines as assessed by RT-qPCR
Knockdown expression of TEAD4 hindered biological function of KYSE-30 cells and this effect was reversed by overexpression of TCF7
To evaluate the effect of TCF7 on TEAD4 expression, overexpression-TCF7 was transfected into KYSE-30 cells. The transfection efficacy of overexpression-TCF7 in KYSE-30 cells were detected by qRT-PCR and Western blot. (Fig. 5A, B). It elucidated that overexpression-TCF7 reversed the facilitation of TEAD4 in cell proliferation (Fig. 5C, D), migration (Fig. 6A–D), and apoptosis (Fig. 7A, B). The mentioned results indicated that interference of TEAD4 hindered biological function of KYSE-30 cells and was reversed by overexpression of TCF7.
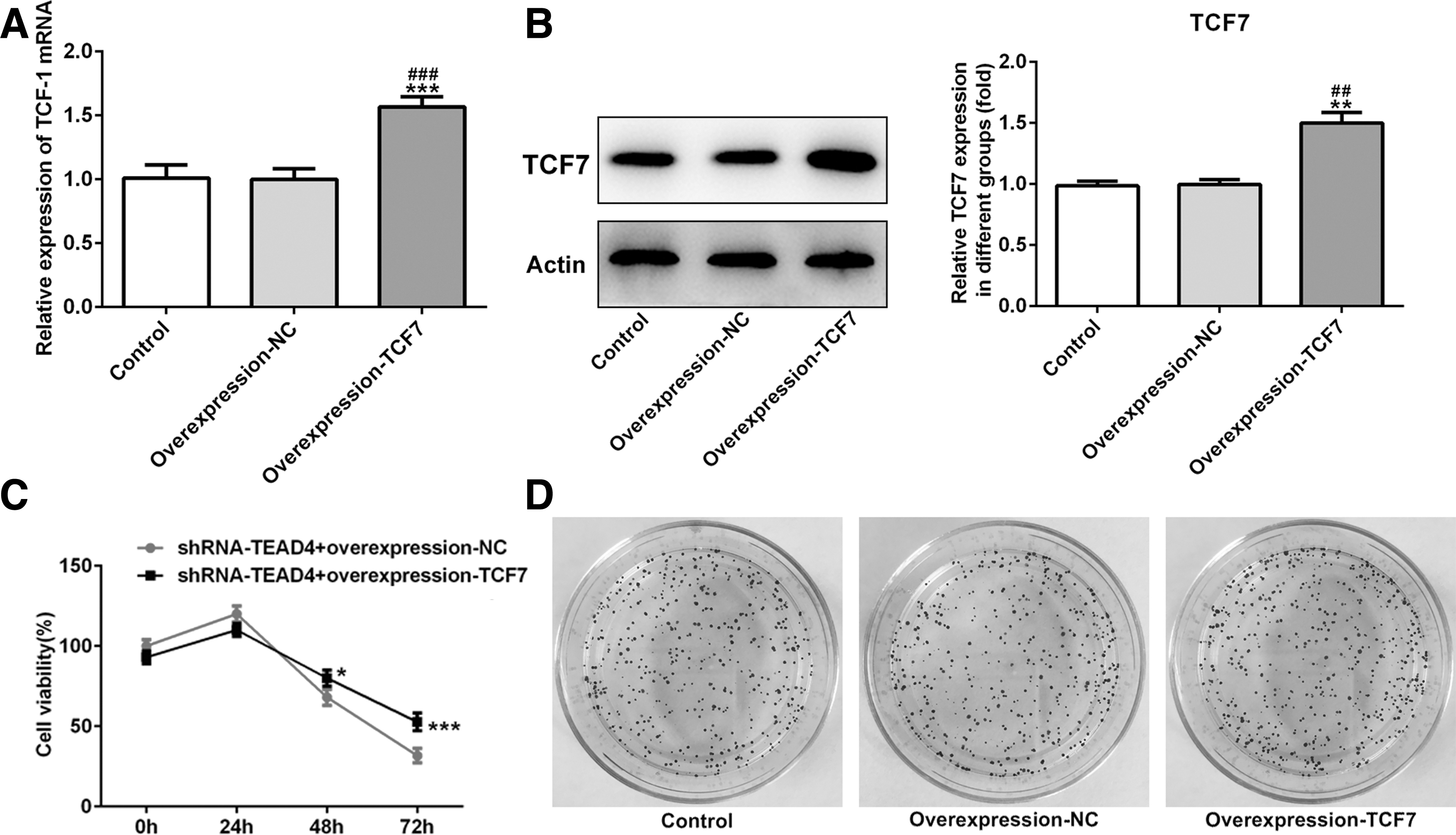
Interference of TEAD4 inhibited the cell proliferation of KYSE-30 cells and this effect was reversed by overexpression of TCF7. The efficiency of transfection was detected by RT-qPCR

Interference of TEAD4 inhibited the cell invasion and migration of KYSE-30 cells and was reversed by overexpression of TCF7. Wound-healing assays
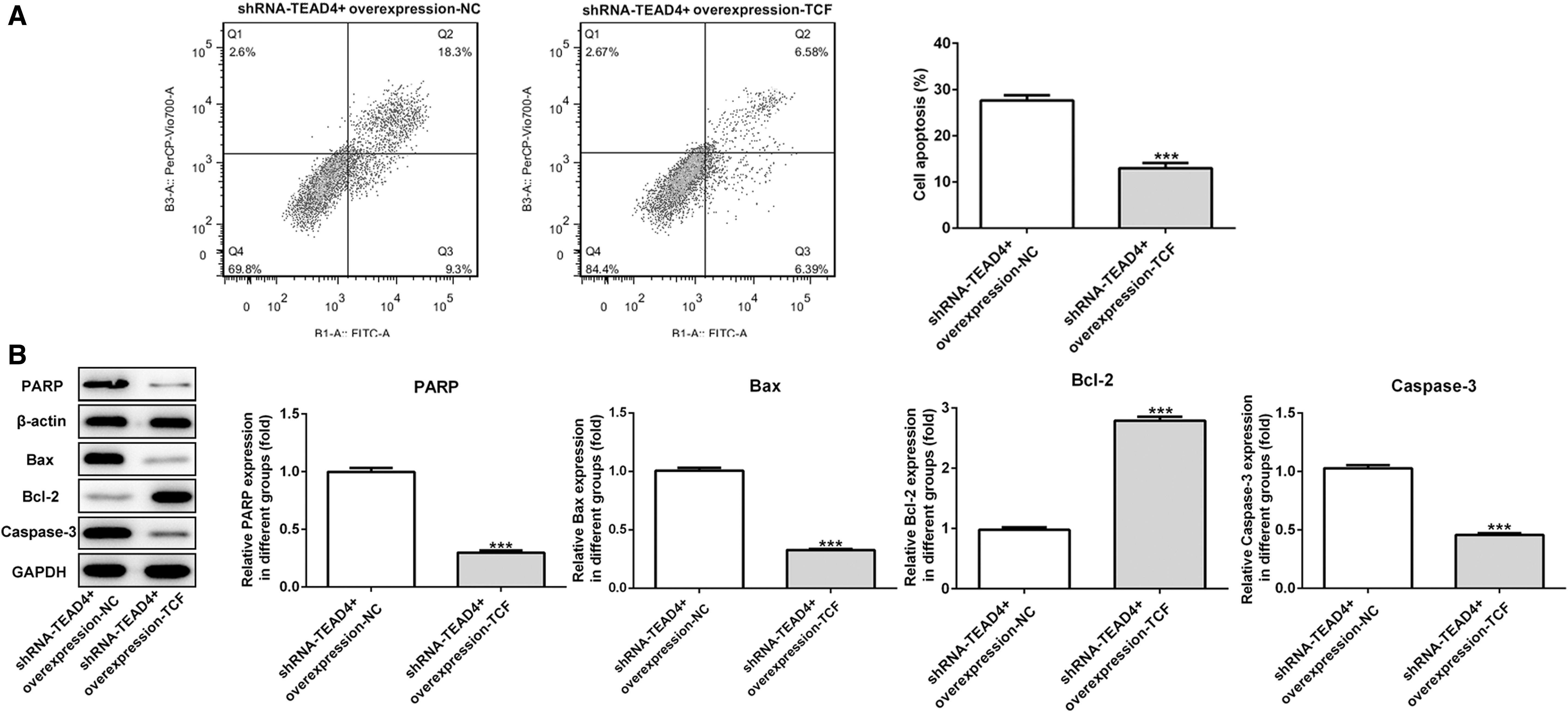
Interference of TEAD4 inhibited the cell apoptosis of KYSE-30 cells and was reversed by overexpression of TCF7.
Discussion
TEAD4 is one of the most important transcription factors in the downstream of YAP/TAZ in the Hippo signaling pathway. 10,11 In recent years, the role of TEAD4 in promoting cancer has been gradually discovered. 12,13 It can form transcription complexes with YAP or independently regulate the expression of downstream-related target genes without YAP. 10,14 It plays a role in promoting cancer in gastrointestinal tumors, liver cancer, lung cancer, breast cancer, and other human solid tumors, leading to the occurrence and progress of tumor, and is a sign of adverse prognosis of many tumors. 9,12,15 In addition, targeted TEAD4 and drugs that block YAP-TEAD4 binding have achieved significant therapeutic effects in vitro and animal models of various tumors, suggesting that TEAD4 may be an ideal target for tumor treatment. 16,17 According to the query on the UALCAN website, it was found that TEAD4 expression in EC patients is significantly higher than that in normal patient tissues, and its expression in EC cells and its effect on proliferation and apoptosis have not been reported. We found that the expression level of TEAD4 was elevated in EC cell lines, which was consistent with the previously reported elevated expression of TEAD in other human solid tumors. It suggests that TEAD4 may as a pro-oncogenic factor in EC.
TCF7 gene is a compound gene, in which several different homotypes have been found, including the homotype lacking β-catenin binding domain. It is reported that TCF7 is the target gene of β-catenin-TCF712 and acts as a feedback repressor of β-catenin-TCF712. TCF7 can help APC to inhibit the malignant transformation of epithelial cells. 18,19 TCF7 was found when using STRING website to interact with TEAD4 proteins. It has been reported that overexpression of TCF7 promotes the progression of gastric cancer, or is associated with poor progression of many types of cancers, 18 and is highly expressed in lung cancer patients. 20 TCF7 can also be used as a target for suppressing miRNAs, suggesting that it has a cancer-promoting effect. 21 According to the UALCAN website, TCF7 expression is elevated in tissues of patients with EC. In this study, it was found that interference with TEAD4 hindered proliferation, invasion, and migration, and promotes apoptosis of KYSE-30 cells, and all the effect was reversed by overexpression of TCF7.
The caspase family plays an important and irreplaceable role in the occurrence and development of apoptosis. In the caspase family, caspase-3, caspase-6, and caspase-7 have been found to be involved in apoptosis. Caspase-3, as the ultimate executor of apoptosis, plays an important role in both exogenous and endogenous apoptotic pathways. 22 In the early stage of apoptosis, caspase-3 exists in the cytoplasm as an inactive precursor. The upstream apoptotic factors first activate the precursor of caspase-3 composed of two large subunits (17KD) and two small subunits (12KD) to form activated caspase-3. Activated caspase-3 cleaved multiple functional proteins and structural proteins in cells, eventually causing cell apoptosis. In this study, Western blot results also showed that after TEAD4 knockout, BCL-2 was significantly decreased, and the expression levels of Bax, PARP, and caspase-3 in KYSE-30 cells were increased, the result indicates that knockdown expression of TEAD4 promotes apoptosis in KYSE-30 cells and was reversed by overexpression of TCF7.
Conclusions
In summary, evidence that interference of TEAD4 hindered proliferation, invasion, and migration, and promoted apoptosis of KYSE-30 cells is presented, and this effect was partly mediated by TCF7. These data indicate that TEAD4 is a latent target for treatment of EC.
Footnotes
Acknowledgments
The authors appreciate ShuDong Laboratory for providing a satisfactory experimental platform and experimental materials.
Availability of Data and Materials
The data sets used and/or analyzed during this study are available from the corresponding author on reasonable request.
Authors' Contributions
Z.W.X. and S.L.H. carried out the study design and writing of the article. K.G. confirmed the completeness and validity of the data. L.M. and S.B. participated in data collection and analysis. Z.W.X. conceived of the study, participated in its study design, and revised the article. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.