
Abstract
Background:
Osteosarcoma (OS) is a common malignant bone tumor. Circular RNAs (circRNAs) exert important roles in the pathogenesis of human cancers, including OS. In this study, the authors focused on the role and mechanism of circRNA signal-induced proliferation-associated 1 like 1 (circ_SIPA1L1) in OS.
Methods:
The enrichment of SIPA1L1, circ_SIPA1L1, microRNA-379-5p (miR-379-5p), and mitogen-activated protein kinase kinase kinase 9 (MAP3K9) was assessed by quantitative real-time polymerase chain reaction (qRT-PCR). Cell proliferation was analyzed by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The colony formation capacity was assessed through colony formation assay. Transwell assays were used to detect the migration and invasion abilities. Western blot assay was used to measure the expression of metastasis-related proteins and MAP3K9. The target interactions between the genes in circ_SIPA1L1/miR-379-5p/MAP3K9 axis were predicted by StarBase and confirmed by dual-luciferase reporter assay. The in vivo role of circ_SIPA1L1 was verified by murine xenograft assay.
Results:
Circ_SIPA1L1 abundance was aberrantly elevated in OS tissues and cell lines. Circ_SIPA1L1 accelerated the proliferation and metastasis abilities of OS cells. Circ_SIPA1L1 promoted the malignant behaviors of OS cells through elevating MAP3K9 level. MiR-379-5p directly bound to circ_SIPA1L1 and MAP3K9. MiR-379-5p interference rescued the abilities of proliferation and metastasis in OS cells, which were suppressed by the silencing of circ_SIPA1L1. Circ_SIPA1L1 promoted the development of OS via miR-379-5p/MAP3K9 in vivo.
Conclusion:
Circ_SIPA1L1 promoted the progression of OS via miR-379-5p/MAP3K9 axis.
Introduction
Osteosarcoma (OS) is a common malignancy generated from mesenchymal cells of bone. 1,2 The conventional treatment method for OS is surgery combined with presurgical or postsurgical chemotherapy. 3,4 The pathogenesis for OS is complex, and radiation and exposure to alkylating agents are risk factors for OS. 5,6 The outcome of metastatic OS patients remains dismal. 7 Therefore, finding effective therapeutic targets is crucial for OS treatment.
Circular RNAs (circRNAs) are noncoding RNAs (ncRNAs) that possess continuous loop structure. 8 CircRNAs are suitable to be biomarkers for cancers due to their stable circular structure. 9 CircRNAs are implicated in the oncogenesis and development of OS. 10 For instance, circ_UBAP2 accelerated OS progression through sponging microRNA-143 (miR-143). 11 Liu et al. claimed that circ_NT5C2 facilitated the development of OS via sponging miR-448. 12 Circ_SIPA1L1 (circ_0032462) has been predicted to accelerate the development of OS. 13 Here, the authors focused on the expression pattern and underlying working mechanism of circ_SIPA1L1 in OS.
MicroRNAs (miRNAs) also belong to ncRNAs, and they possess 18–24 nucleotides. 14 MiRNAs could regulate the progression of cancers through degrading or restraining the translation of target genes. 15 For instance, Jiang et al. reported that miR-329 repressed the progression of OS through downregulating RAB10. 16 Xie et al. found that miR-302b impeded the metastasis of OS cells through suppressing Runx2. 17 MiR-379-5p played an antitumor role in bladder cancer, hepatocellular carcinoma, and OS. 18 –20 For instance, Xie et al. found that miR-379 prevented the malignancy of OS through inhibiting EIF4G2. 20 In the current study, they found another regulatory pathway of miR-379-5p in OS.
Mitogen-activated protein kinase kinase kinase 9 (MAP3K9), also named mixed-lineage kinase 1 (MLK1), was identified as an oncogene in cancers. 21 For example, Xia et al. found that MAP3K9 acted as the target of miR-7 to promote the progression of pancreatic cancer. 22 Zhang et al. found that MAP3K9 functioned as the target of miR-148a to accelerate the progression of esophagus squamous cell carcinoma. 23 Zhao et al. claimed that MAP3K9 facilitated the proliferation of OS cells. 24 However, the working mechanism of MAP3K9 in OS was not fully illustrated.
The clinical data indicated that circ_SIPA1L1 was aberrantly upregulated in OS tissues, implying that circ_SIPA1L1 might play pivotal role in OS progression. Gain-of-function experiments and loss-of-function experiments were carried out to disclose the functions of circ_SIPA1L1 in OS. The regulatory mechanism by which circ_SIPA1L1 mediated the malignancy of OS cells was then explored in the following experiments.
Materials and Methods
Tissue samples
OS tissues (n = 50) and adjacent normal tissues (n = 50) were resected from OS patients during surgery in Weifang People's Hospital. The diagnosis of OS tissues and normal tissues was conducted by two pathologists. All subjects had signed written informed consents, and these OS patients had not received chemotherapy or radiotherapy before surgery. The procedures were authorized by the Ethics Committee of Weifang People's Hospital. Pathological characteristics and information of OS patients, including age, gender, multifocality, family history, and TNM stage, are listed in Table 1.
Pathological Characteristics and Information of Osteosarcoma Patients
Cell line
MG63, HOS, U2OS, and SaOS-2 and human osteoblasts hFOB 1.19 were obtained from BeNa Culture Collection (Beijing, China). All cell lines were grown with high glucose Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA added with 10% fetal bovine serum (FBS; Gibco) and 10% penicillin (100 U/mL)/streptomycin (100 μg/mL). hFOB 1.19 cell line was cultured in the humidified incubator with 5% CO2 at 33.5°C, while OS cell lines were maintained in the humidified incubator with 5% CO2 at 37°C.
Quantitative real-time polymerase chain reaction
Quantitative real-time polymerase chain reaction (qRT-PCR) was carried out to amplify circ_SIPA1L1, SIPA1L1, miR-379-5p, and MAP3K9 with SYBR Green qPCR Master Mix (Thermo Fisher Scientific, Waltham, MA). The expression was analyzed using 2−ΔΔCt formula and was normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or U6. All primers are listed in Table 2.
Primers Used in Quantitative Real-Time Polymerase Chain Reaction Assay
Cell transient transfection
Circ_SIPA1L1 overexpression vector (circ_SIPA1L1), control vector (Vector), circ_SIPA1L1-specific small interfering RNA (si-circ_SIPA1L1), siRNA negative control (si-NC), miR-379-5p, miR-NC, anti-miR-379-5p, anti-miR-NC, MAP3K9 overexpression vector (MAP3K9), and empty vector (pcDNA) were obtained from Genepharma (Shanghai, China).
Two μg plasmids or 50 nM oligonucleotides were transiently transfected into OS cells using Opti-MEM (Invitrogen, Carlsbad, CA) and Lipofectamine 3000 (Invitrogen).
Establishment of stably transfected MG63 cell line
Stably transfected MG63 cell line was used for murine xenograft assay. Circ_SIPA1L1-specific short hairpin RNA (sh-circ_SIPA1L1) and control (sh-NC) were purchased from Genepharma. Lentiviruses were infected into MG63 cells with the presence of 8 μg/mL polybrene (Genepharma). After infection for 72 h, 2 μg/mL puromycin was used to select stably transfected MG63 cell line.
Three-(4,5-Dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide (MTT) assay
After transfection for 0, 1, 2, or 3 d, 0.5% MTT (5 mg/mL; Sigma) was added to incubate with the OS cells, and cells were continued to cultivate for a further 4 h. The absorbance (490 nm) was examined to analyze the number of viable OS cells.
Colony formation assay
Single-cell suspension was prepared, and OS cells were seeded into six-well plates at the concentration of 200 cells/well. OS cells were incubated for 2 weeks to form the visible colonies. After 24-h cultivation, nonadherent OS cells were discarded. Culture medium was refreshed every 3 d. The colonies were stained, and the number of colonies was counted.
Transwell assays
In transwell migration assay, 100 μL cell suspension was added into the upper chambers of 24-well transwell plates (Corning Costar, Corning, NY) in triplicate. Ten percent FBS (600 μL DMEM) acted as chemotactic factor in the lower chambers. After incubation for 24 h, the OS cells were immobilized using 4% paraformaldehyde for 30 min, and OS cells were dyed with 0.05% Gentian Violet for 5 min. The number of migrated OS cells in five random fields was counted using an optical microscope. In transwell invasion assay, OS cell suspension was added into the upper chambers after precoating BD Matrigel matrix (BD Biosciences, San Jose, CA). The other procedures were similar as above.
Western blot assay
The protein samples were quantified, and then separated using separation gel, and then the proteins were blotted to the PVDF membrane (Millipore, Billerica, MA). The membrane was incubated with the primary antibodies, including anti-snail (ab229701; Abcam, Cambridge, MA), anti-twist family bHLH transcription factor 1 (anti-Twist1; ab49254; Abcam), anti-E-cadherin (ab40772; Abcam), anti-MAP3K9 (ab228752; Abcam), and anti-GAPDH (ab8245; Abcam). The membrane was then probed with secondary antibody. The protein bands were measured with the enhanced chemiluminescent (ECL) system (Beyotime, Shanghai, China).
Dual-luciferase reporter assay
StarBase was utilized to seek the targets of circ_SIPA1L1 and miR-379-5p, and dual-luciferase reporter assay was used to confirm these predictions. Circ_SIPA1L1-wt, circ_SIPA1L1-mut, MAP3K9-wt, and MAP3K9-mut were constructed through inserting the wild-type or the mutant type of the sequences of circ_SIPA1L1 and MAP3K9 into PYr-MirTarget luciferase vectors (Ambion, Austin, TX). After cotransfecting, the luciferase activities in different transfection groups were examined.
Murine xenograft assay
Murine xenograft assay was applied with the permission of the Animal Care and Use Committee of Weifang People's Hospital. Mice were randomly divided into two groups, including sh-NC group (n = 5; inoculation with MG63 cells stably expressing sh-NC) and sh-circ_SIPA1L1 group (n = 5; inoculation with MG63 cells stably expressing sh-circ_SIPA1L1). Two × 106 MG63 cells suspended in 200 μL phosphate-buffered saline (PBS) were inoculated into the flank of the mice on week 0. Tumor size was monitored every week with the method of volume = π/6 × length × width × height. The mice were euthanized, and the dissected tumors were weighted at the end point (4 weeks after inoculation).
Statistical analysis
GraphPad Prism 7.0 was used to conduct the statistical analysis. The differences of groups were evaluated with Student's t-test and one-way analysis of variance (ANOVA). The linear relationship among circ_SIPA1L1, miR-379-5p, and MAP3K9 was analyzed using Spearman's correlation coefficient. The data are displayed as the form of mean ± standard deviation (SD). p-Value <0.05 indicated a significant difference.
Results
The level of circ_SIPA1L1 is enhanced in OS
A conspicuous elevation in the abundance of circ_SIPA1L1 was found in OS tissues (n = 50) compared with adjacent nontumor tissues (n = 50) (Fig. 1A), implied that circ_SIPA1L1 might exert important role in OS. To disclose the functions of circ_SIPA1L1 in cellular level, the authors first detected the enrichment of circ_SIPA1L1 in OS cells and normal osteoblasts hFOB 1.19. Consistent with the tissue-level data, the expression of circ_SIPA1L1 was extremely enhanced in OS cells than that in hFOB 1.19 (Fig. 1B), especially in MG63 and U2OS cells.
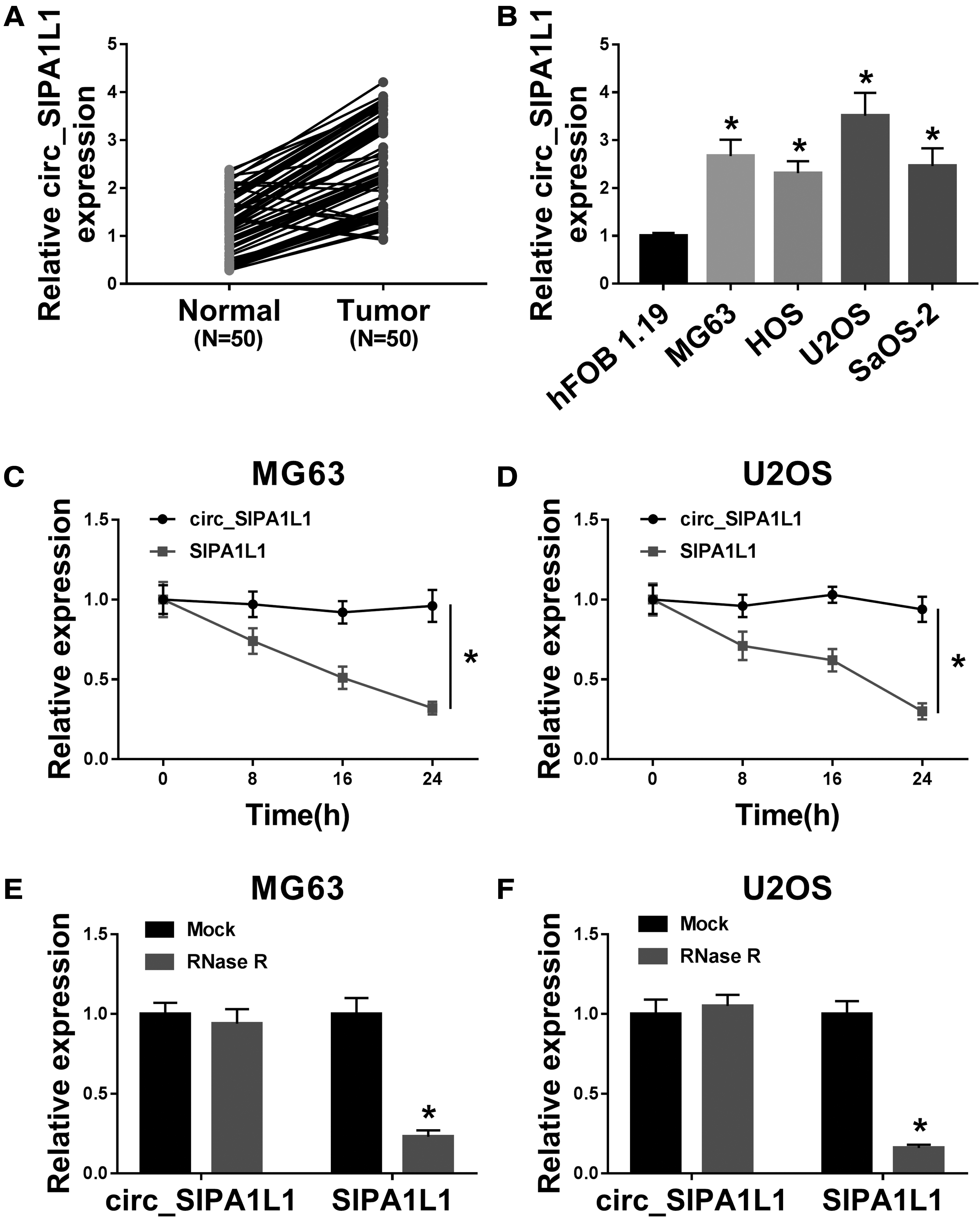
The level of circ_SIPA1L1 is enhanced in OS.
To verify the closed loop structure of circ_SIPA1L1, actinomycin D and RNase R were used. As mentioned in Figure 1C and D, actinomycin D (transcription inhibitor) was used to inhibit the synthesis of RNA, and the levels of circ_SIPA1L1 and its linear form SIPA1L1 were monitored after treatment for 0, 8, 16, or 24 h via qRT-PCR. Circ_SIPA1L1 was more stable than its matching linear mRNA. As exhibited in Figure 1E and F, circ_SIPA1L1 was more resistant to RNase R than SIPA1L1 mRNA. Taken together, circ_SIPA1L1 possessed the closed loop structure, and it was aberrantly upregulated in OS tissues and cell lines.
Circ_SIPA1L1 promotes the proliferation, migration, and invasion of OS cells
The biological functions of circ_SIPA1L1 were explored in MG63 and U2OS cells through gain-of-function experiments and loss-of-function experiments. The transfection efficiencies of si-circ_SIPA1L1 and circ_SIPA1L1 were high in OS cells (Fig. 2A). The absorbance in OS cells was declined with the depletion of circ_SIPA1L1 (Fig. 2B, C), and the transfection of si-circ_SIPA1L1 also decreased the number of colonies (Fig. 2D), demonstrated that circ_SIPA1L1 silencing restrained the proliferation of OS cells. The number of migration or invasion OS cells was lesser in si-circ_SIPA1L1 group than that in si-NC group (Fig. 2E, F), suggested that circ_SIPA1L1 knockdown suppressed the migration and invasion of OS cells.

Circ_SIPA1L1 promotes the proliferation, migration, and invasion of OS cells.
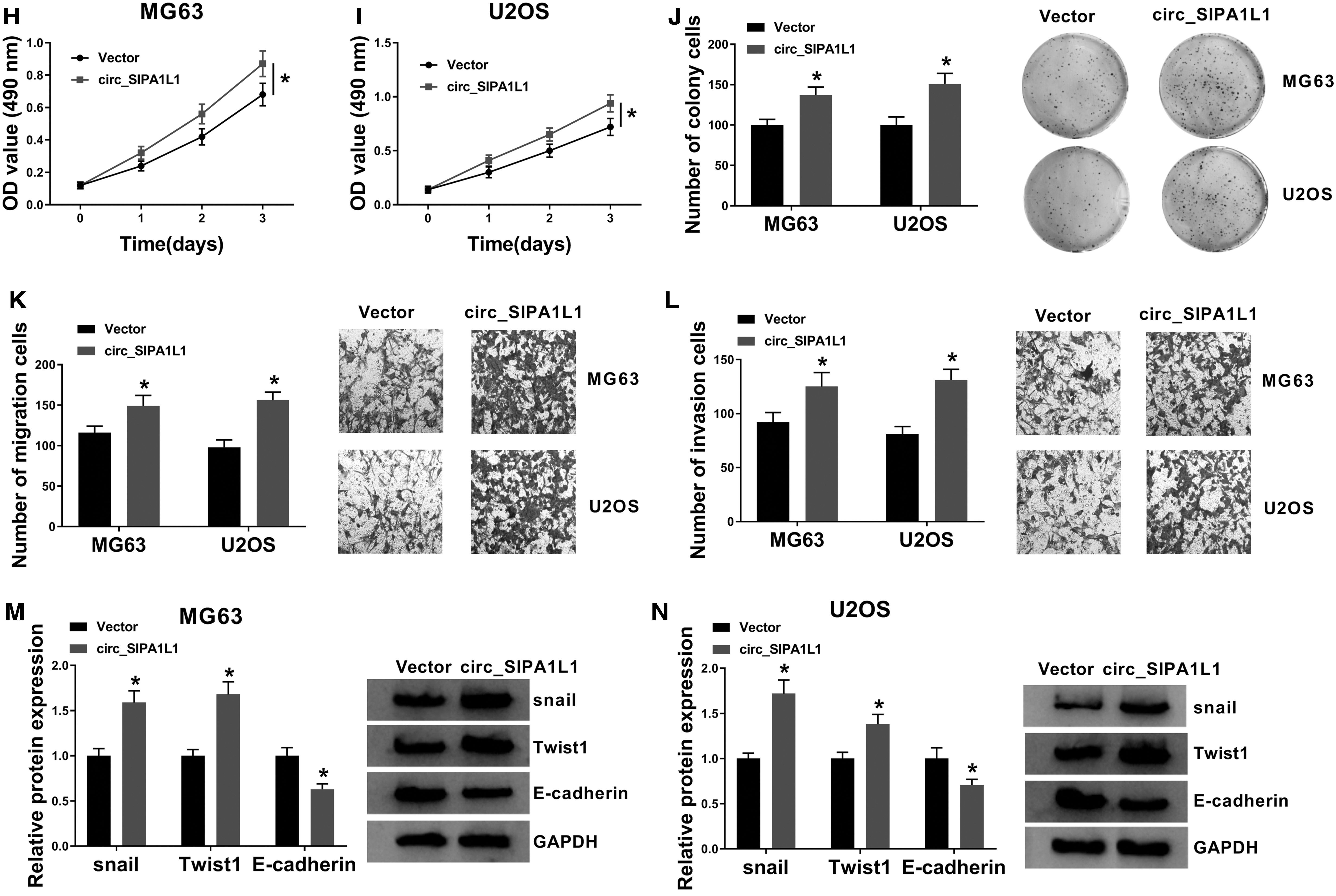
The authors measured the expression of metastasis-related proteins (snail, Twist1, and E-cadherin) to further confirm the role of circ_SIPA1L1 in the metastasis of OS cells. There was a notable reduction in the expression of snail and Twist1, while a significant increase in the abundance of E-cadherin was found in si-circ_SIPA1L1 group in comparison with that in si-NC group (Fig. 2G), demonstrated that circ_SIPA1L1 silencing suppressed the motility of OS cells.
Subsequently, circ_SIPA1L1 recombinant plasmid was used to explore the influences of circ_SIPA1L1 overexpression on the behaviors of OS cells. As depicted in Figure 2H–J, OS cells transfected with circ_SIPA1L1 exhibited an increased proliferation ability via MTT assay and colony formation assay. Besides, according to the results of transwell assays and Western blot assay, the metastasis of OS cells was prominently accelerated in circ_SIPA1L1 group in contrast to Vector group (Fig. 2K–N). These findings together demonstrated that circ_SIPA1L1 exerted an oncogenic role to facilitate the proliferation and motility of OS cells.
Circ_SIPA1L1 promotes OS progression through upregulating MAP3K9
The enrichment of MAP3K9 mRNA and protein was also found to be abnormally upregulated in OS tumor tissues than that in adjacent normal tissues (Fig. 3A, C). Given the similar expression trend of circ_SIPA1L1 and MAP3K9 in OS, they analyzed the linear correlation of these two molecules via Spearman's correlation coefficient. A significant positive correlation was found between the expression of circ_SIPA1L1 and MAP3K9 mRNA in OS tissues (Fig. 3B). The abundance of MAP3K9 mRNA and protein was both elevated in OS cells relative to hFOB 1.19 cells (Fig. 3D, E).

Circ_SIPA1L1 promotes OS progression through upregulating MAP3K9.
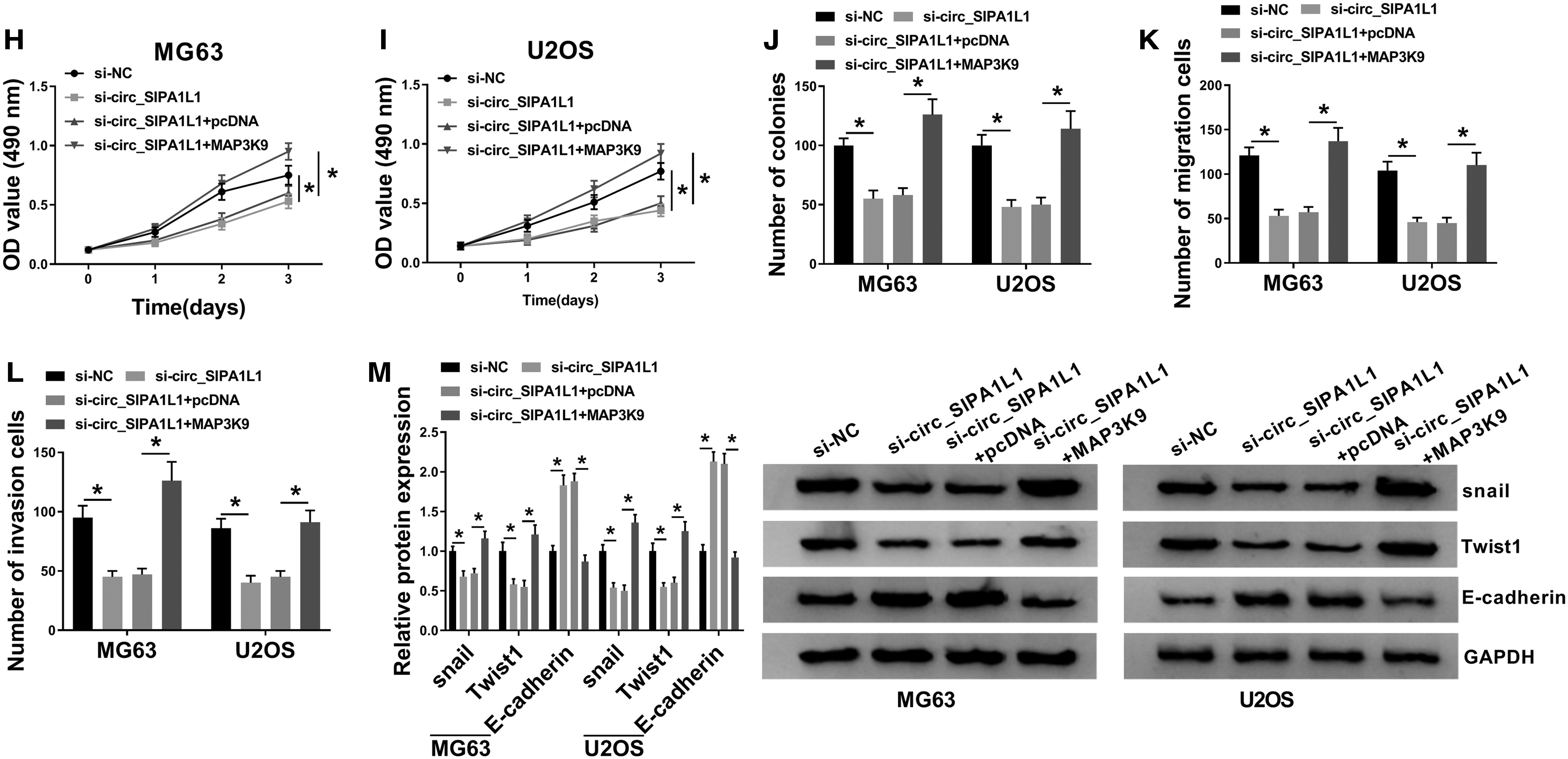
The authors cotransfected si-circ_SIPA1L1 together with pcDNA or MAP3K9 into OS cells to explore the functional correlation between MAP3K9 and circ_SIPA1L1. The mRNA and protein levels of MAP3K9 were rescued in si-circ_SIPA1L1 and MAP3K9 group, which were decreased by the transfection of si-circ_SIPA1L1 (Fig. 3F, G). As mentioned in Figure 3H–J, si-circ_SIPA1L1-mediated proliferation inhibition was reversed by the addition of MAP3K9 in OS cells. The metastasis of OS cells was impeded in si-circ_SIPA1L1 group, and the cotransfection of si-circ_SIPA1L1 and MAP3K9 rescued the motility ability of OS cells (Fig. 3K, L).
The epithelial-mesenchymal transition (EMT) markers were detected to further elucidate the effect of circ_SIPA1L1 and MAP3K9 on the metastasis of OS cells. The abundance of snail and Twist1 was restrained with the depletion of circ_SIPA1L1, and the addition of MAP3K9 regained the abundance of snail and Twist1 in OS cells, and the changes in the expression of E-cadherin exhibited an opposite trend to snail or Twist1 (Fig. 3M). In summary, circ_SIPA1L1 contributed to the progression of OS via enhancing the level of MAP3K9.
Circ_SIPA1L1/miR-379-5p/MAP3K9 axis is identified in OS
MiR-379-5p was a potential target of circ_SIPA1L1 predicted by StarBase software (Fig. 4A). The transfection of miR-379-5p mimic instead of miR-NC caused a dramatic downregulation in the luciferase activity of OS cells transfected with circ_SIPA1L1-wt, while there was no notable change in the luciferase activity of OS cells in circ_SIPA1L1-mut group with the cotransfection of miR-NC or miR-379-5p (Fig. 4B, C), demonstrated that miR-379-5p bound to circ_SIPA1L1 via the putative binding sites.
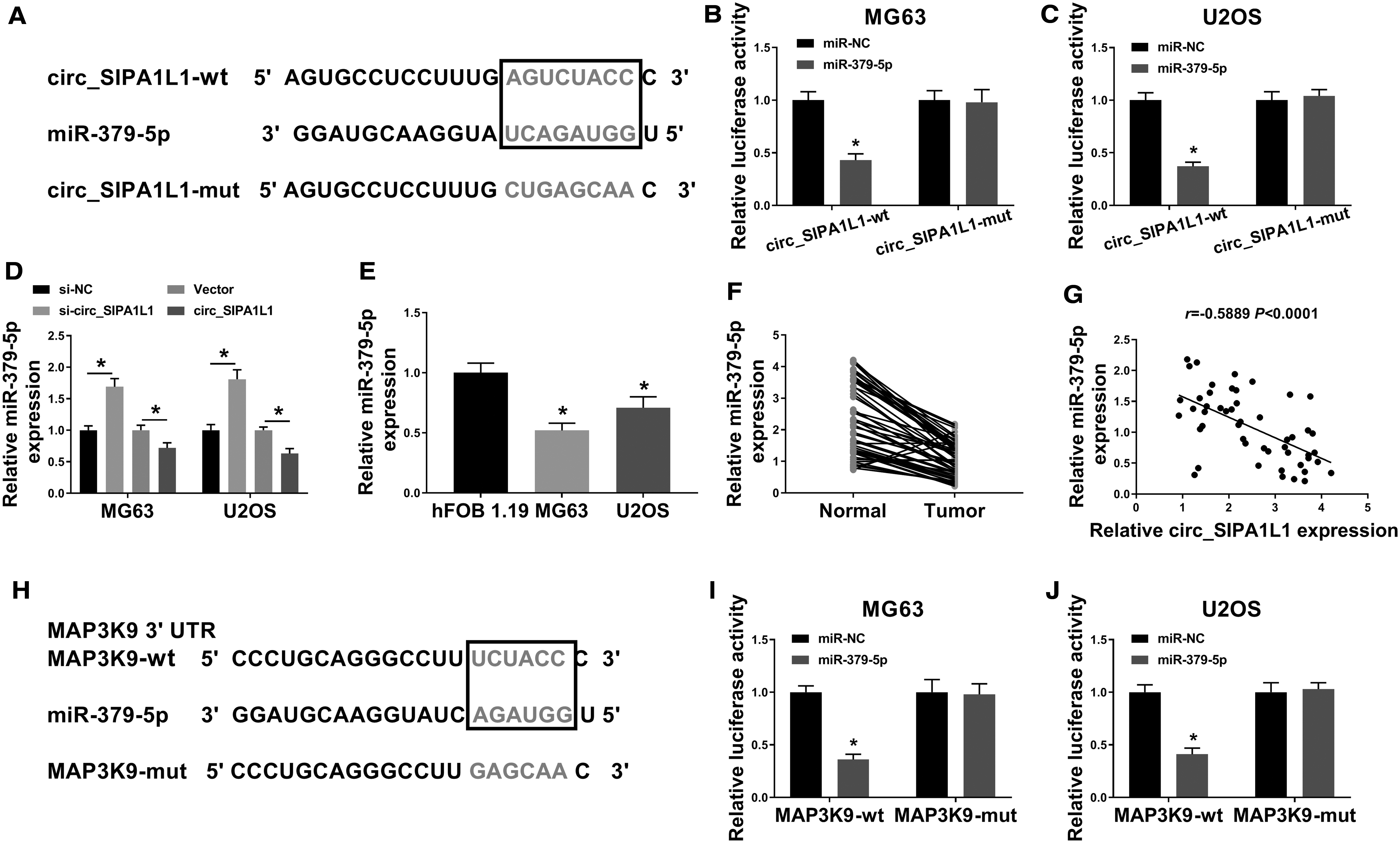
Circ_SIPA1L1/miR-379-5p/MAP3K9 axis is identified in OS.
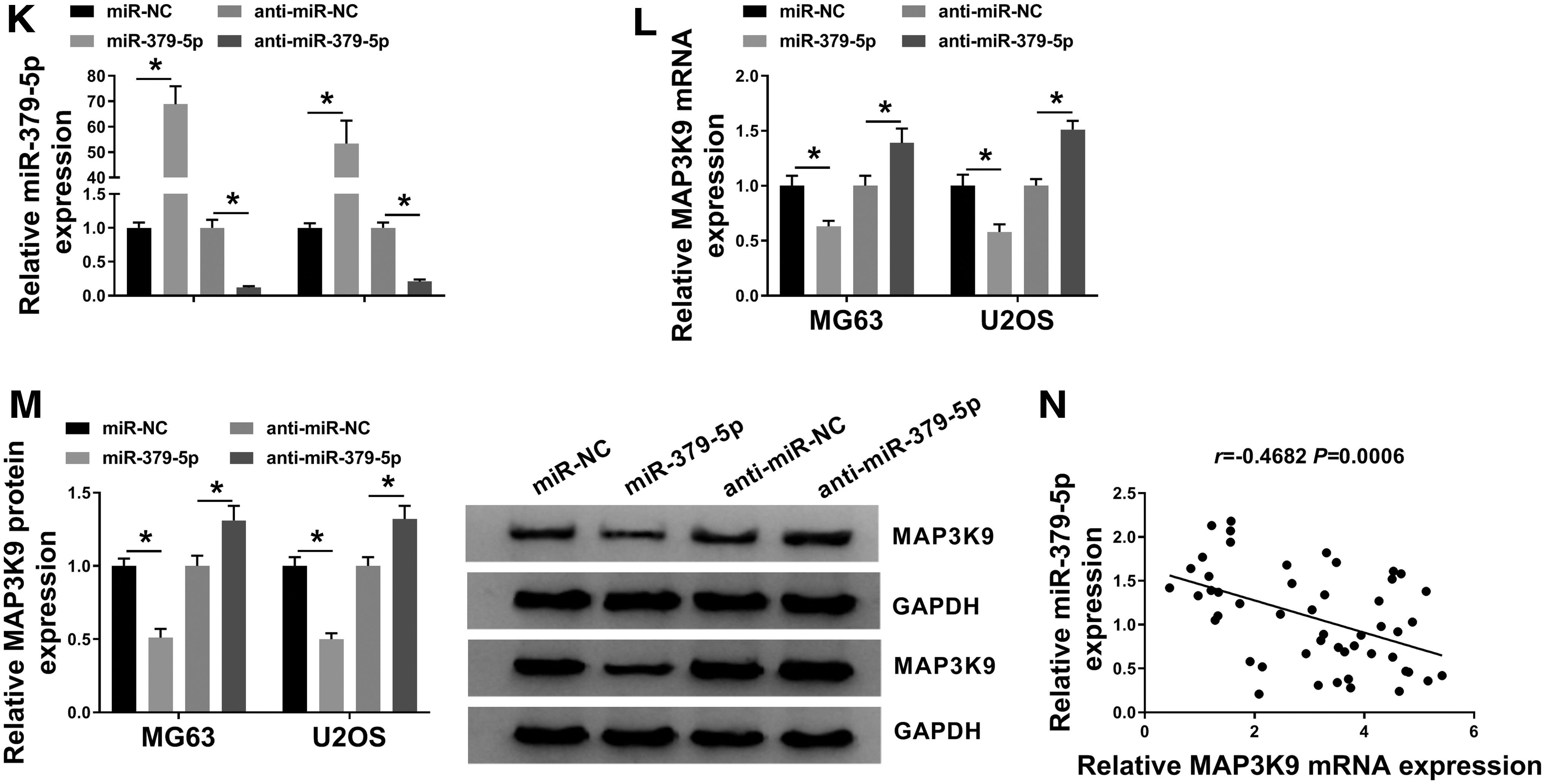
As exhibited in Figure 4D, the transfection of si-circ_SIPA1L1 conspicuously increased the expression of miR-379-5p, and the accumulation of circ_SIPA1L1 downregulated the abundance of miR-379-5p, suggesting the negative regulatory relationship between cir4c_SIPA1L1 and miR-379-5p in OS cells. There was a reduced abundance of miR-379-5p in OS cell lines and tissue samples relative to hFOB 1.19 cell line and corresponding nontumor tissues (Fig. 4E, F). Based on the expression of circ_SIPA1L1 and miR-379-5p in each subject, a negative linear relationship between miR-379-5p and circ_SIPA1L1 was found (Fig. 4G).
The 3′ untranslated region (3′ UTR) of MAP3K9 possessed the complementary sites with miR-379-5p (Fig. 4H). The luciferase activity was notably decreased in miR-379-5p and MAP3K9-wt group relative to miR-NC and MAP3K9-wt group (Fig. 4I, J), suggesting the interaction between miR-379-5p and MAP3K9 in OS cells.
The authors initially assessed the overexpression and interference efficiencies of miR-379-5p and anti-miR-379-5p in OS cells. As shown in Figure 4K, the overexpression and knockdown efficiencies of miR-379-5p and anti-miR-379-5p were both high in OS cells. The overexpression of miR-379-5p reduced the mRNA and protein expression of MAP3K9, while the mRNA and protein levels of MAP3K9 were upregulated with the silencing of miR-379-5p (Fig. 4L, M). As expected, a notable inverse correlation was found between the expression of miR-379-5p and MAP3K9 mRNA (Fig. 4N). In summary, circ_SIPA1L1/miR-379-5p/MAP3K9 was first identified in OS cells.
Circ_SIPA1L1 accelerates the proliferation, migration, invasion, and EMT of OS cells through suppressing miR-379-5p
The enrichment of MAP3K9 mRNA and protein was decreased with the silencing of circ_SIPA1L1, and the addition of anti-miR-379-5p regained the mRNA and protein expression of MAP3K9 in OS cells (Fig. 5A, B). The inhibitory effect mediated by the interference of circ_SIPA1L1 in the proliferation of OS cells was counteracted by the cotransfection of si-circ_SIPA1L1 and anti-miR-379-5p via MTT assay and colony formation assay (Fig. 5C–E). Besides, the transfection of anti-miR-379-5p also attenuated the suppressive influences of si-circ_SIPA1L1 transfection on the migration and invasion of OS cells (Fig. 5F, G).
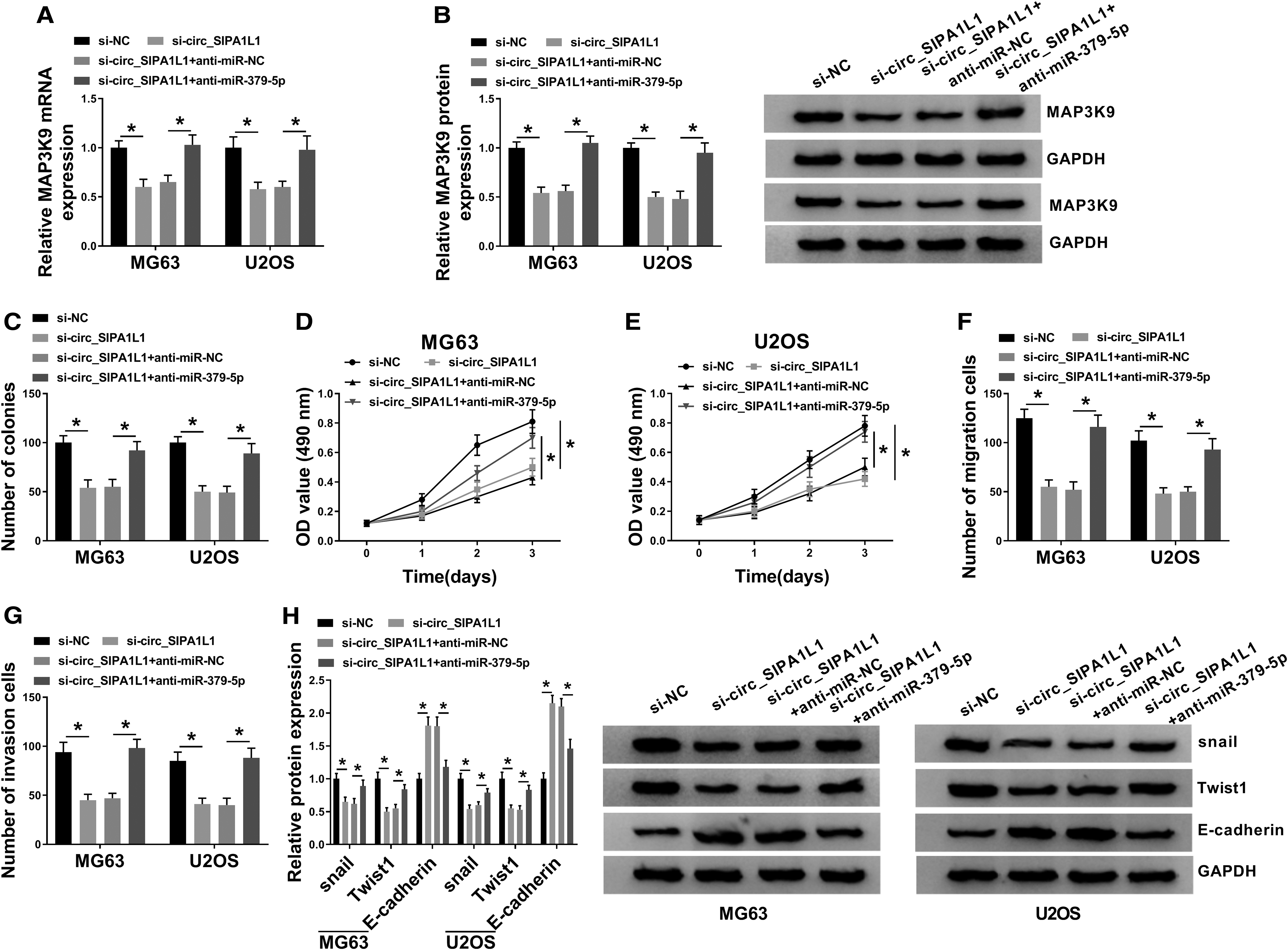
Circ_SIPA1L1 accelerates the proliferation, migration, invasion, and EMT of OS cells through suppressing miR-379-5p. MG63 and U2OS cells were transfected with si-NC, si-circ_SIPA1L1, si-circ_SIPA1L1 + anti-miR-NC, or si-circ_SIPA1L1 + anti-miR-379-5p.
Furthermore, circ_SIPA1L1 silencing-mediated downregulation in snail and Twist1 in OS cells was largely rescued with the introduction of anti-miR-379-5p (Fig. 5H). The expression trend of E-cadherin exhibited an opposite phenomenon to snail or Twist1 (Fig. 5H), suggested that circ_SIPA1L1 promoted the metastasis of OS cells partly via sponging miR-379-5p. These results revealed that circ_SIPA1L1 exerted its protumor role in OS through sponging and downregulating miR-379-5p.
Circ_SIPA1L1 depletion prevents the growth of OS tumors in vivo
To verify whether circ_SIPA1L1 exerted a similar role in vivo, the authors monitored the volume and weight of OS tumors in nude mice inoculated with 2 × 106 MG63 cells (200 μL PBS) stably expressing sh-NC or sh-circ_SIPA1L1. As exhibited in Figure 6A and B, OS tumors were smaller in nude mice injected with MG63 cells stably expressing sh-circ_SIPA1L1 other than sh-NC.
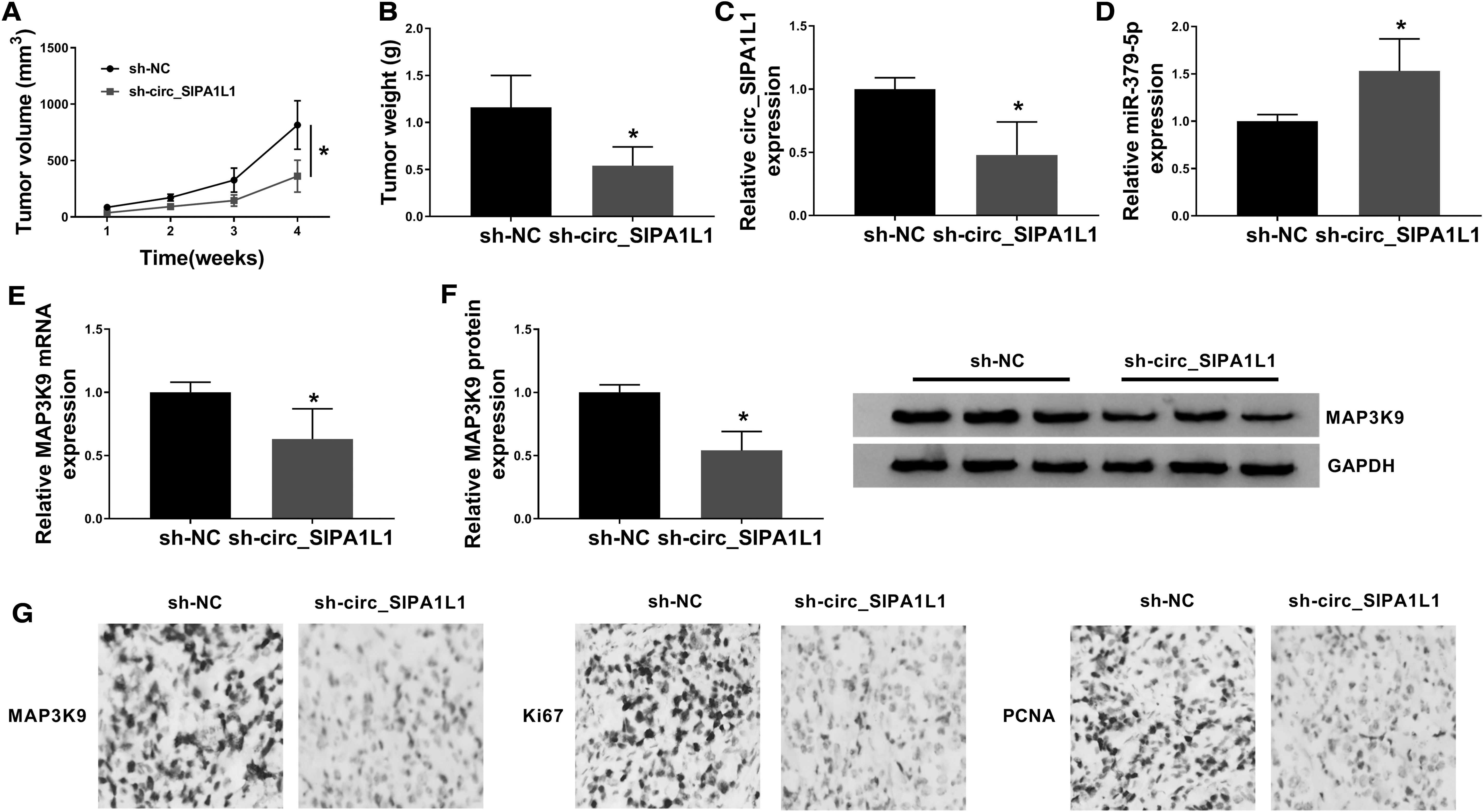
Circ_SIPA1L1 depletion prevents the growth of OS tumors in vivo.
The enrichment of circ_SIPA1L1 was decreased in tumor tissues generated from sh-circ_SIPA1L1 group, while the abundance of miR-379-5p was upregulated in sh-circ_SIPA1L1 group (Fig. 6C, D). Apart from these, a significant upregulation in the mRNA and protein expression of MAP3K9 were observed in sh-circ_SIPA1L1 group compared with that in sh-NC group (Fig. 6E, F). According to the results of IHC, the levels of MAP3K9 and two proliferation markers (Ki67 and PCNA) were all reduced in tumor tissues in sh-circ_SIPA1L1 group compared with sh-NC group (Fig. 6G). In summary, circ_SIPA1L1 accelerated the OS tumor growth in vivo.
Circ_SIPA1L1 exacerbates OS through miR-379-5p/MAP3K9 axis
Through the in vitro and in vivo experiments using qRT-PCR, MTT assay, colony formation assay, transwell assays, Western blot assay, dual-luciferase reporter assay and murine xenograft model, it was concluded that circ_SIPA1L1 accelerated the progression of OS through upregulating MAP3K9 via sponging miR-379-5p (Fig. 7).
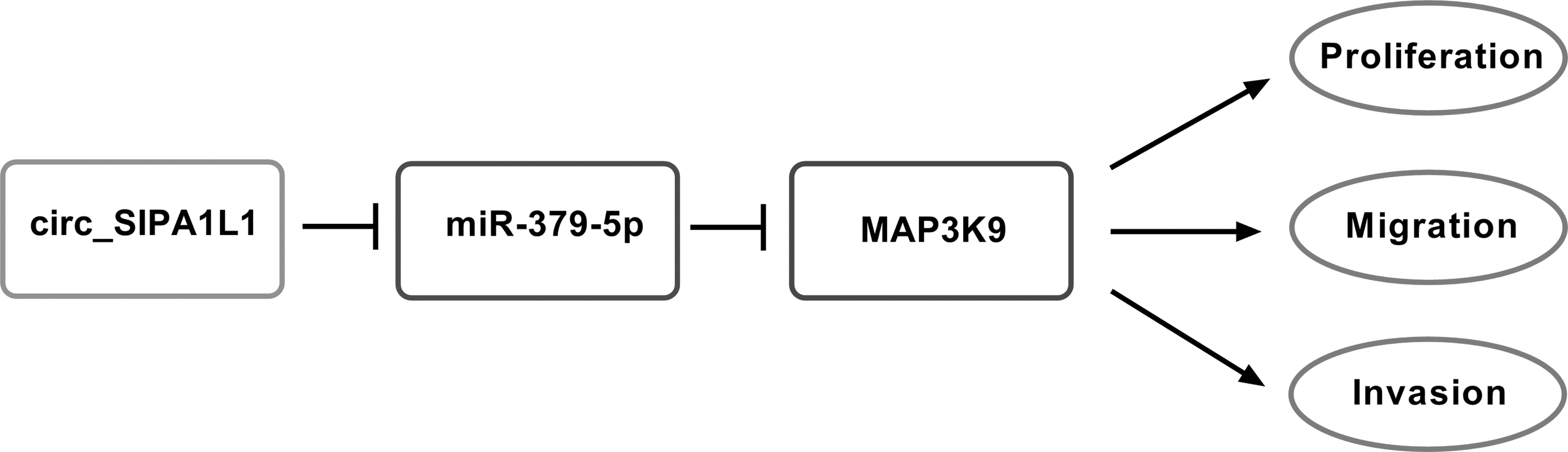
Circ_SIPA1L1 exacerbates OS through miR-379-5p/MAP3K9 axis. The mechanism diagram revealed that circ_SIPA1L1 accelerated the proliferation, migration, and invasion of OS cells through sequestering miR-379-5p from MAP3K9.
Discussion
OS is a primary malignant bone tumor that occurs in adolescents. 25 The survival rate of patients with metastatic OS remains low although improvement has been made in the treatment strategy of OS. 26 Therefore, it is necessary to find crucial molecules that are implicated in the initiation and development of OS.
CircRNAs have emerged as pivotal regulators in OS. Wang et al. claimed that circ_0003998 accelerated proliferation and invasion of OS cells via miR-197-3p. 27 Zhang et al. claimed that circ_0051079 facilitated proliferation and metastasis of OS through miR-26a-5p/TGF-β1 axis. 28 Circ_SIPA1L1 was predicted to promote the development of OS through upregulating CADM1. 13 In the study, a significant upregulation of circ_SIPA1L1 expression was found in OS tissues and cells, suggesting the important role of circ_SIPA1L1 in OS.
EMT is characterized by the decrease of cell-cell adhesion, and it causes the promotion of metastasis in tumor cells. 29,30 Besides, migration and invasion abilities are the crucial features of cancer cells. 31 Functional experiments demonstrated that circ_SIPA1L1 was an oncogene in OS, and it potentiated the proliferation, migration, invasion, and EMT of OS cells.
MiR-379-5p was a tumor suppressor in many cancers. Lv et al. found that circ_PITX1 contributed to the development of glioblastoma through sponging miR-379-5p to upregulate MAP3K2. 32 Wu et al. claimed that miR-379-5p inhibited the proliferation and motility of bladder cancer cells via MDM2. 18 Chen et al. demonstrated that miR-379-5p repressed the metastasis of hepatocellular carcinoma cells through FAK/AKT pathway. 19 Xie et al. claimed that miR-379 restrained the proliferation and metastasis of OS cells via EIF4G2. 20 Li et al. found that miR-379 restrained the progression of OS through targeting PDK1. 33
The authors found that miR-379-5p was a direct target of circ_SIPA1L1 in OS cells. Besides, the depletion of miR-379-5p alleviated si-circ_SIPA1L1-mediated inhibition on the proliferation and motility of OS cells, suggesting the antitumor role of miR-379-5p in OS, which was in accordance with previous articles. 20,33
MAP3K9 is a member of MAPK signaling. MAP3K9 was the target of many miRNAs, containing miR-148a, miR-490-5p, and miR-15a. 23,34,35 Herein, MAP3K9 was validated as a downstream gene of miR-379-5p in OS cells. Zhang et al. reported the oncogenic role of MAP3K9 in esophagus squamous cell carcinoma, and miR-148a prevented the progression of esophagus squamous cell carcinoma through inhibiting MAP3K9. 23 Abdeyrim et al. found that MAP3K9 promoted the proliferation, metastasis, and EMT of pharyngolaryngeal cancer cells. 34
The oncogenic role of MAP3K9 was also found in OS. Zhao et al. demonstrated that miR-1247 played an antitumor role in OS through targeting MAP3K9. 24 Echoing with the former studies, it was found that MAP3K9 acted as the target of circ_SIPA1L1/miR-379-5p to accelerate the proliferation and metastasis of OS cells. Through conducting murine xenograft assay, it was found that circ_SIPA1L1 silencing also suppressed OS tumor growth in vivo.
There are also some limitations in this study. For example, the abundance of circ_SIPA1L1, miR-379-5p, and MAP3K9 was detected in OS by qRT-PCR, and the application of the in situ hybridization will make the results more reliable.
Taken together, circ_SIPA1L1 potentiated the malignancy of OS cells through accelerating the proliferation, migration, and invasion via targeting miR-379-5p/MAP3K9 axis. The identification of circ_SIPA1L1/miR-379-5p/MAP3K9 axis provides a novel entry point in OS treatment.
Footnotes
Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
No funding was received for this article.