
Abstract
Background:
The occurrence of cisplatin (DDP) resistance in oral squamous cell carcinoma (OSCC) is a major challenge for OSCC treatment. Circular RNAs (circRNAs) have been associated with the development of cancer resistance, but the role of circ_0109291 in DDP resistance of OSCC is unclear.
Methods:
The expression of circ_0109291 and microRNA-188-3p (miR-188-3p) was determined by quantitative real-time polymerase chain reaction (qRT-PCR). Cell counting kit 8 (CCK8) assay, colony formation assay, and flow cytometry were performed to measure the DDP resistance, proliferation, and apoptosis of cells. The levels of apoptosis-related proteins and ATP-binding cassette sub-family B member 1 (ABCB1) protein were assessed via Western blot (WB) analysis. In addition, dual-luciferase reporter assay, RNA immunoprecipitation (RIP), and RNA pull-down assay were used to illuminate the mechanism of circ_0109291. Animal experiments were employed to confirm the effect of circ_0109291 on OSCC tumor growth in vivo.
Results:
Circ_0109291 was higher expressed in DDP-resistant OSCC tissues and cells, and its knockdown suppressed proliferation and resistance and enhanced the apoptosis of OSCC cells. MiR-188-3p could be sponged by circ_0109291, and its overexpression had an inhibition effect on the DDP resistance of OSCC cells. ABCB1 was a target of miR-188-3p. Further experiments confirmed that both miR-188-3p inhibitor and ABCB1 overexpression also could invert the suppression effect of circ_0109291 silencing on the DDP resistance of OSCC cells. In vivo experiments revealed that silenced circ_0109291 could improve the sensitivity of the tumor to DDP.
Conclusion:
Circ_0109291 could promote the DDP resistance of OSCC, suggesting that silenced circ_0109291 might be a key step to inhibit OSCC resistance.
Introduction
Oral squamous cell carcinoma (OSCC) is a common malignancy of the oral cavity. 1 OSCC usually causes oral dysfunction and can be cured with surgery or radiation in the early stages of the disease. However, advanced OS is usually accompanied by lymph node metastasis, so it is difficult to treat. 2 Surgery combined with radiotherapy and chemotherapy is the main treatment for OSCC at present. 3,4 Cisplatin (DDP) is a common chemotherapy drug, but the occurrence of DDP resistance is a huge obstacle for OSCC treatment. 5,6 Therefore, it is urgent to find the potential mechanisms affecting resistance and provide new ideas for the treatment of cancer resistance.
The progression of OSCC involves a variety of biological processes, including the regulation of circular RNAs (circRNAs), which have been extensively studied in recent years. 7 CircRNAs are a class of stable transcripts with closed loops, whose role in cancer progression has been elucidated. 8,9 The role of different circRNAs in OSCC progression is also different. For example, circ_0004491 was lower expressed in OSCC and its silencing could enhance the metastasis of OSCC, 10 and circ_100290 was higher expressed in OSCC and it could promote the proliferation and glycolysis of OSCC. 11 It has been studied that the increased expression of circ_0109291 in OSCC is associated with malignant biological behaviors of OSCC cells and poor prognosis of OSCC patients. 12 But its effect on the resistance of OSCC has not been studied.
MicroRNAs (miRNAs) are a kind of small non-coding RNAs that can regulate the degradation and translation of messenger RNA (mRNA) at the transcriptional level. 13 Currently, the role of miRNA in many diseases and cancers has been widely confirmed. 14,15 MiR-188-3p has decreased expression in many cancers, including hepatocellular carcinoma and breast cancer. 16,17 Besides, studies have shown that miR-188-3p is associated with drug resistance of pancreatic cancer. 18 However, there are few studies on miR-188-3p in OSCC. ATP-binding cassette sub-family B member 1 (ABCB1) is a multidrug-resistant protein that has been reported to be significantly increased in DDP-resistant OSCC cells. 19,20
Our study explored the role and molecular mechanism of circ_0109291 in the DDP resistance of OSCC. In this, we found that circ_0109291 could promote DDP resistance of OSCC by regulating the miR-188-3p/ABCB1 axis, which provided new ideas for the treatment of OSCC resistance.
Materials and Methods
Samples collection
Sixty OSCC patients were recruited from The Third Hospital of Mianyang, including 31 DDP-sensitive patients and 29 DDP-resistant patients, all of whom signed the informed consent. OSCC tissues and adjacent normal tissues were obtained and stored at −80°C. This study was approved by the Ethics Committee of The Third Hospital of Mianyang and was carried out in accordance with the Ethics of Declaration of Helsinki.
Cell culture and DDP-resistant cell lines construction
OSCC cell lines (SCC-4, SCC-9, CAL-27, UM1, and UM2) and normal human oral keratinocytes (NHOK) were bought from the National Typical Culture Collection (NTCC, Beijing, China). Cells were grown in Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA) containing 10% fetal bovine serum (FBS; Gibco), and 1% antibiotic mixture (Invitrogen, Carlsbad, CA) at 37°C with 5% CO2 incubator.
CAL-27 and UM1 cells were repeatedly cultured in a medium with increasing DDP (Yuanye Bio, Shanghai, China) concentration until all DDP-sensitive cells died. The surviving CAL-27 and UM1 cells were DDP-resistant cells (CAL-27/DDP and UM1/DDP). This process lasted about 5 months. All DDP-resistant cells were cultured in DMEM containing 1 μg/mL of DDP to maintain their resistance. When the experiment started, the culture conditions of CAL-27/DDP and UM1/DDP cells were consistent with normal cells.
Quantitative real-time polymerase chain reaction
The TRIzol reagent (Invitrogen) was used to extract total RNA. The PrimeScript RT Regent Kit (Takara, Dalian, China) was employed to reverse-transcript RNA into cDNA. Quantitative real-time polymerase chain reaction (qRT-PCR) analysis was performed using SYBR Green (Takara). β-actin was used as the reference gene for circ_0109291, and U6 was considered as the internal control for miR-188-3p. All primers were listed as follows: circ_0109291: F, 5′-GGATGAATCCCCAGGAGACG-3′, R 5′-CTGAGCAGGGTTCAGGCATT-3′; β-actin: F, 5′-ATAGCACAGCCTGGATAGCAACGTAC-3′, R, 5′-CACCTTCTACAATGAGCTGCGTGTG-3′; miR-188-3p: F, 5′-CCCTCTCTCACATCCCTTGCAT-3′, R, 5′-ATCCTGCAAACCCTGCATGTG-3′; U6: F, 5′-GCTTCGGCAGCACATATACTAA-3′; R, 5′-AACGCTTCACGAATTTGCGT-3′. The relative expression was calculated with the 2−ΔΔCt method.
Cell counting kit 8 assay
The DDP resistance and proliferation of cells were assessed using this assay. All cells were seeded in 96-well plates. On the one hand, cells were treated with different concentrations of DDP (0.25, 0.5, 1, 2, 4, 8, 16, and 32 μg/mL) for 48 h. After that, cell counting kit 8 (CCK8) solution (MSK Bio, Wuhan, China) was added into cells and cultured for 4 h. Cell viability was measured at 450 nm and half-maximal inhibitory concentration (IC50) value was calculated to assess the cell resistance to DDP. On the other hand, cells were incubated with CCK8 solution for 4 h at a specific time point (24, 48, and 72 h). Absorbance was detected at 450 nm to evaluate cell proliferation.
Cell transfection
Small interference RNA targeting circ_0109291 (si-circ_0109291) and negative control (si-NC), miR-188-3p mimic or inhibitor (miR-188-3p or anti-miR-188-3p) and negative controls (miR-NC or anti-NC), ABCB1 overexpression vector (ABCB1) and scrambled control (vector) were purchased from RiboBio (Guangzhou, China). Lentiviral short hairpin RNA targeting circ_0109291 (sh-circ_0109291) and scrambled shRNA control (sh-NC) were obtained from GenePharma (Shanghai, China). Lipofectamine 3000 reagent (Invitrogen) was used for transfection when the cell confluence rate reached 50%.
Colony formation assay
After transfection for 48 h, CAL-27/DDP and UM1/DDP cells were seeded into six-well plates and cultured for 14 days. Then, cells were fixed with methanol and stained with 0.1% crystal violet. The colony numbers of CAL-27/DDP and UM1/DDP cells more than 50 cells were counted under a microscope.
Flow cytometry
Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) Apoptosis Detection Kit (Yeasen, Shanghai, China) was used for this assay. Briefly, CAL-27/DDP and UM1/DDP cells were harvested after transfection for 48 h. After having stained with Annexin V-FITC and PI, the apoptosis rate of cells was analyzed using a flow cytometer (BD Biosciences, Franklin Lakes, NJ).
Western blot analysis
Total protein was extracted using RIPA buffer (Beyotime, Shanghai, China), separated using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gel, and shifted to polyvinylidene fluoride (PVDF) membranes (Roche, Basel, Switzerland). Then, the membranes were blocked with 5% nonfat milk, incubated overnight at 4°C with primary antibodies, and interacted with secondary antibody (1:2000; Sigma-Aldrich, St. Louis, MO) for 1 h. Finally, protein bands were detected with the BeyoECL star (Beyotime). All primary antibodies were bought from Sigma-Aldrich and were listed as follows: B-cell lymphoma-2 (Bcl-2; 1:1000), Bcl2-associated x (Bax; 1:2000), Cleaved-caspase-3 (Cleaved-cas-3, 1:1000), ABCB1 (1:1,500) or β-actin (1:2000).
Dual-luciferase reporter assay
The sequences of wild-type (WT) and mutant-type (MUT) circ_0109291 or ABCB1 3′UTR were subcloned into the psiCHECK-2 reporter vector (Promega, Madison, WI) to build the circ_0109291-WT/MUT or ABCB1 3′UTR-WT/MUT reporter vector. CAL-27/DDP and UM1/DDP cells were seeded into 24-well plates and transfected with the reporter vectors together with miR-188-3p mimic or miR-NC. After transfection for 48 h, luciferase activities were evaluated using the Dual-Luciferase Reporter Assay System (Promega).
RNA immunoprecipitation assay
EZ-Magna RNA immunoprecipitation (RIP) Kit (Millipore, Billerica, MA) was used for this assay. CAL-27/DDP and UM1/DDP cells were lysed by RIP buffer (Millipore). Magnetic beads were conjugated with immunoglobulin G antibody (anti-IgG) or argonaute2 antibody (anti-Ago2) and incubated with the cell lysates overnight at 4°C. After having resuspended with proteinase K, the immunoprecipitated RNA was isolated and the enrichment of circ_0109291 and miR-188-3p was determined using qRT-PCR.
RNA pull-down assay
CAL-27/DDP and UM1/DDP cells were transfected with biotin-labeled miR-188-3p probe (Bio-miR-188-3p; Sangon, Shanghai, China) or negative control probe (Bio-miR-NC; Sangon). After 48 h, the cells were lysed using RIP buffer (Millipore), and the cell lysates were incubated with magnetic beads. The enrichment of circ_0109291 was detected using qRT-PCR.
Mice xenograft models
CAL-27/DDP cells (2 × 106) stably transfected with sh-NC or sh-circ_0109291 were subcutaneously injected into the flanks of BALB/c nude mice (Junke Bio, Nanjing, China). After 1 week, mice had received DDP (5 mg/kg) every week. The control group was injected with the same amount of normal saline. Three weeks after treated with DDP, the mice were sacrificed on day 28, and tumor tissues were collected for subsequent experiments. The experiment was approved by the Ethics Committee of The Third Hospital of Mianyang and was carried out in accordance with the National Institutes of Health guide for the care and use of laboratory animals.
Statistical analysis
All data were expressed as mean ± standard deviation and analyzed using the Student's t-test (for two groups) or one-way analysis of variance (for multiple groups). The relationship between circ_0109291 and miR-188-3p was evaluated using the Pearson correlation analysis. GraphPad Prism 5.0 software (GraphPad Software, San Diego, CA) was used to perform statistical analysis. Significance was established as p < 0.05.
Results
Circ_0109291 was highly expressed in DDP-resistant OSCC tissues and cells
Compared with adjacent normal tissues, circ_0109291 was remarkably upregulated in OSCC tissues and was higher in DDP-resistant OSCC tissues than in DDP-sensitive OSCC tissues (Fig. 1A, B). Besides, in OSCC cell lines (SCC-4, SCC-9, CAL-27, UM1, and UM2), we also found that circ_0109291 had elevated expression compared with that in NHOK cells (Fig. 1C). Subsequently, we constructed DDP-resistant cell lines and detected their resistance to DDP by CCK8. The results showed that CAL-27/DDP and UM1/DDP cells showed good DDP resistance with the increase of DDP concentration, and the IC50 value was markedly increased in CAL-27/DDP and UM1/DDP cells, indicating that the DDP-resistant OSCC cell lines were successfully constructed (Fig. 1D, E). Moreover, circ_0109291 was also highly expressed in CAL-27/DDP and UM1/DDP cells compared with that in CAL-27 and UM1 cells (Fig. 1F). Hence, we speculated that circ_0109291 might play a key role in the DDP resistance of OSCC.
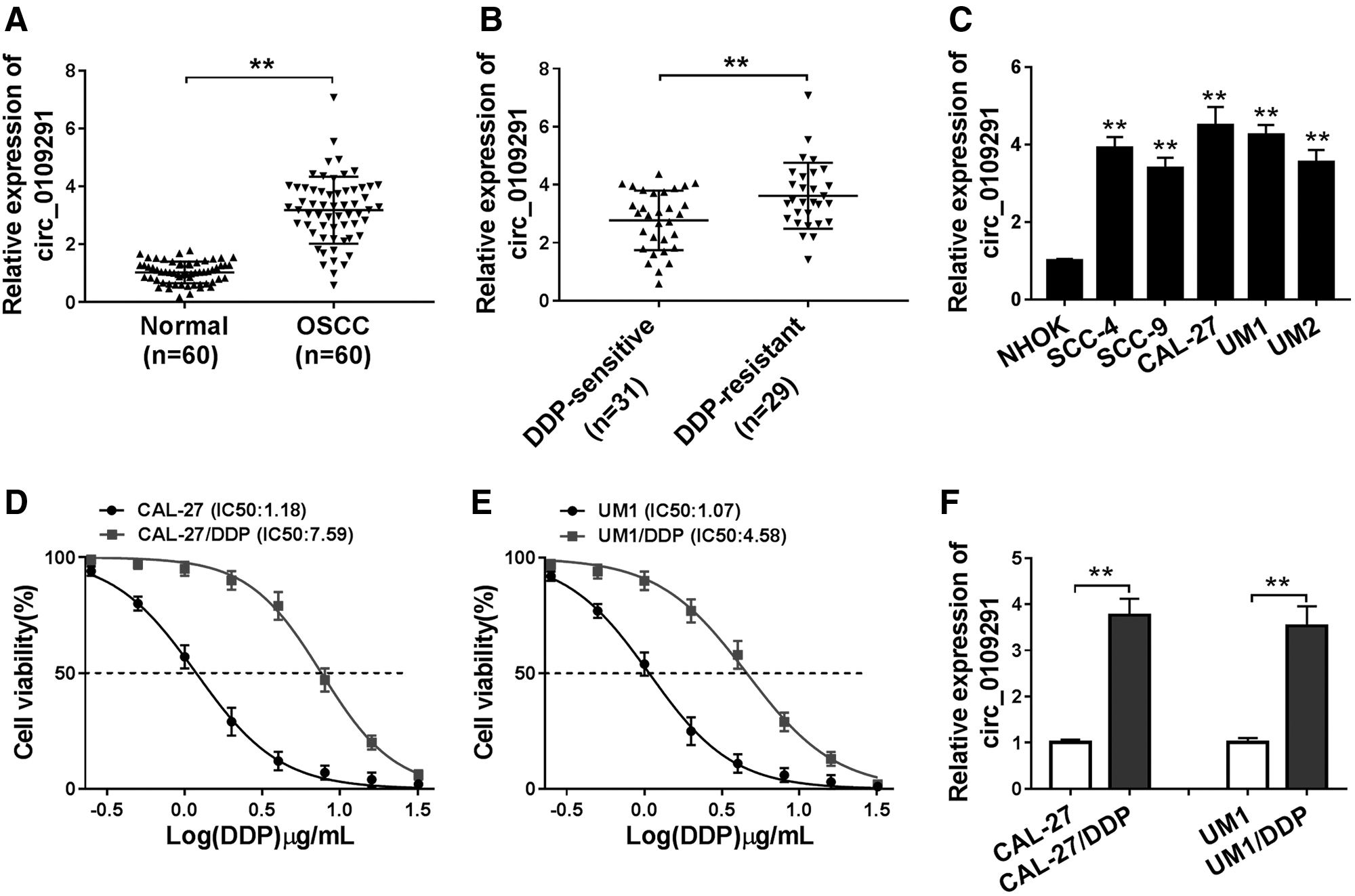
The circ_0109291 expression in OSCC tissues and cells.
Depletion of circ_0109291 inhibited proliferation and resistance and promoted apoptosis in DDP-resistant OSCC cells
To investigate the role of circ_0109291 in the DDP resistance of OSCC, we knocked down its expression using si-circ_0109291. As shown in Figure 2A, si-circ_0109291 showed a good silencing effect on circ_0109291 expression. CCK8 and colony formation assay results indicated that silenced circ_0109291 could repress the viabilities and the colony numbers of CAL-27/DDP and UM1/DDP cells, suggesting that circ_0109291 knockdown suppressed the proliferation of DDP-resistant OSCC cells (Fig. 2B–D). Meanwhile, silenced circ_0109291 also decreased the IC50 value of CAL-27/DDP and UM1/DDP cells, indicating that its knockdown could reduce the resistance of CAL-27/DDP and UM1/DDP cells to DDP (Fig. 2E, F). Besides, we also measured the apoptosis rate of cells and found that the silencing of circ_0109291 could enhance the apoptosis of CAL-27/DDP and UM1/DDP cells, which also could be confirmed by the decreased Bcl-2 protein level and the increased Bax and Cleaved-cas3 protein levels (Fig. 2G, H).

Inhibition of circ_0109291 suppressed the DDP resistance of OSCC cells.
Circ_0109291 interacted with miR-188-3p in OSCC
To explore the mechanism by which circ_0109291 affected the progression and resistance of OSCC, we used the Circinteractome tool (

Circ_0109291 interacted with miR-188-3p in OSCC.
Overexpression of miR-188-3p restrained the DDP resistance of OSCC
For exploring the function of miR-188-3p in OSCC, we performed gain-functional experiments using miR-188-3p mimic. QRT-PCR showed that miR-188-3p mimic could effectively improve the expression of miR-188-3p in CAL-27/DDP and UM1/DDP cells (Fig. 4A). Next, we measured the proliferation of CAL-27/DDP and UM1/DDP cells using CCK8 and colony formation assays. The results showed that the viabilities and colony numbers of CAL-27/DDP and UM1/DDP cells were remarkably suppressed in the miR-188-3p overexpression group, suggesting that elevated miR-188-3p expression could inhibit the proliferation of DDP-resistant OSCC cells (Fig. 4B–D). Further, miR-188-3p overexpression also lowered the resistance of CAL-27/DDP and UM1/DDP cells to DDP, which could be obtained by the reduction of IC50 value (Fig. 4E, F). In addition, the promotion of miR-188-3p overexpression on the apoptosis rate, the reduction of Bcl-2 protein level and the acceleration of Bax and Cleaved-cas-3 protein levels also confirmed that miR-188-3p overexpression could enhance the apoptosis of CAL-27/DDP and UM1/DDP cells (Fig. 4G–I). Based on these results, we found that the effect of miR-188-3p mimic on the DDP resistance of OSCC cells was similar to that of circ_0109291 knockdown.
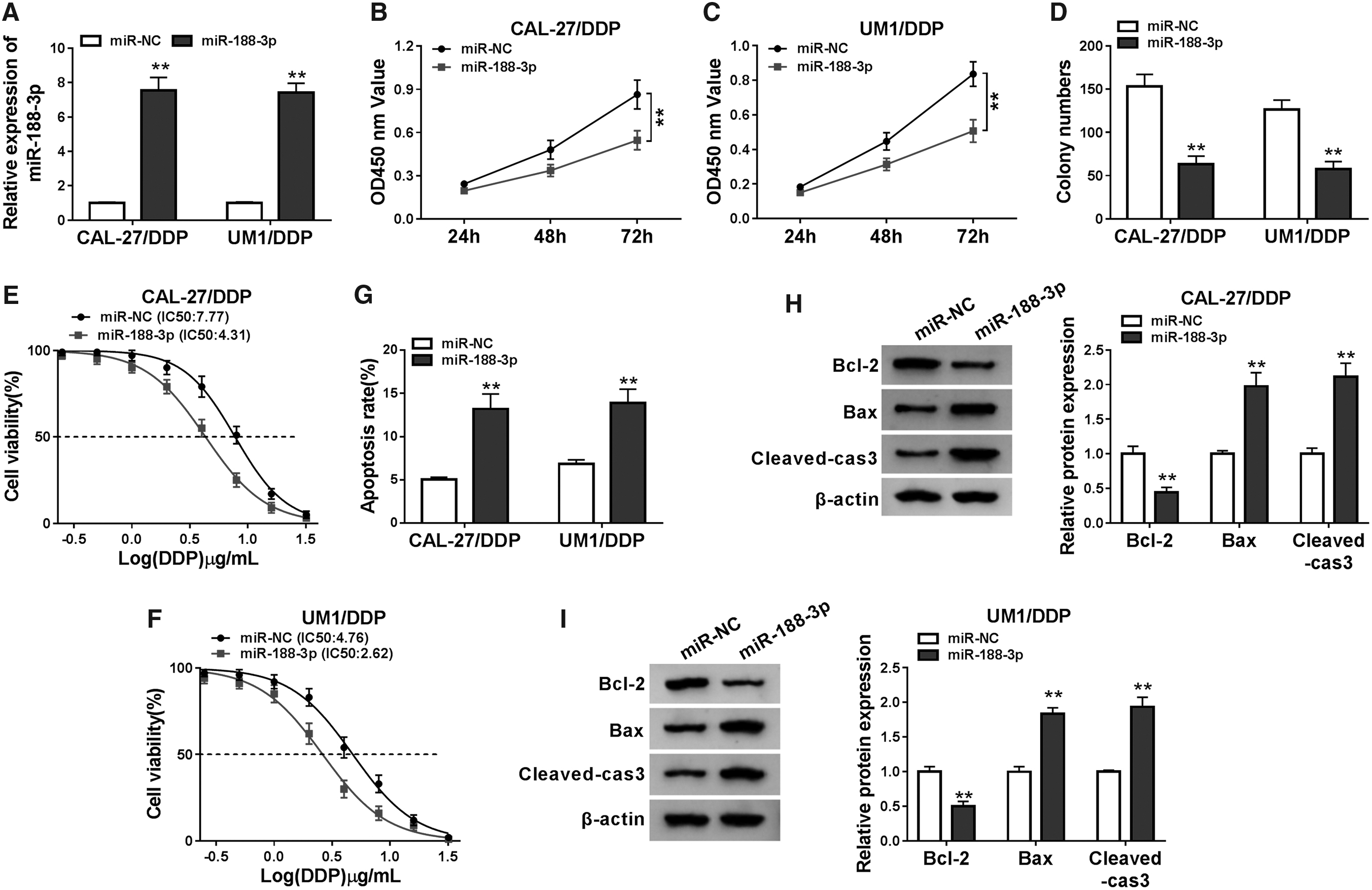
Overexpression of miR-188-3p restrained the DDP resistance of OSCC cells.
ABCB1 served as a target of miR-188-3p in OSCC
To improve the circRNA-miRNA-mRNA regulatory network, we used the TargetScan tool to predict the downstream targets of miR-188-3p and found that ABCB1 3′UTR had binding sites with miR-188-3p (Fig. 5A). And dual-luciferase reporter assay confirmed that the luciferase activity of ABCB1 3′UTR-WT rather than ABCB1 3′UTR-MUT could be hindered by miR-188-3p mimic (Fig. 5B, C). Meanwhile, we discovered that ABCB1 expression was upregulated in DDP-resistant OSCC tissues and cells compared with that in their corresponding controls through Western blot (WB) analysis (Fig. 5D, E). By detecting the ABCB1 protein level, we also uncovered that its expression could be suppressed by miR-188-3p overexpression while increased by miR-188-3p inhibition in CAL-27/DDP and UM1/DDP cells (Fig. 5F). Furthermore, circ_0109291 could restrain the ABCB1 protein level in CAL-27/DDP and UM1/DDP cells, and miR-188-3p inhibitor could reverse this effect (Fig. 5G). In a word, we confirmed that ABCB1 was a target of miR-188-3p and could be indirectly regulated by circ_0109291 in OSCC.

ABCB1 served as a target of miR-188-3p in OSCC.
MiR-188-3p inhibitor or ABCB1 overexpression could reverse the effect of circ_0109291 silencing on the DDP resistance of OSCC cells
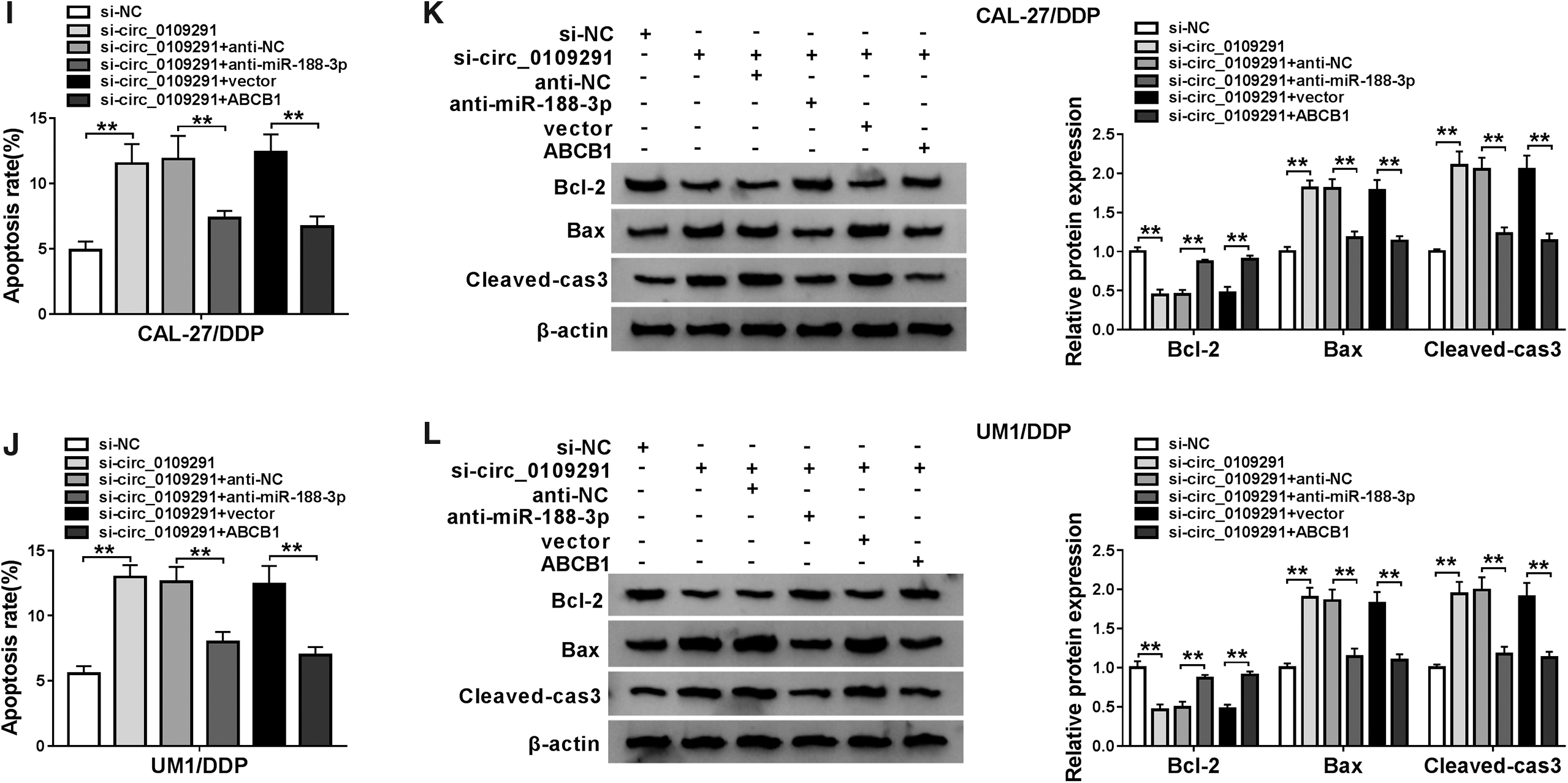
To further confirm our conclusion above, we conducted rescue experiments. The inhibitory effect of anti-miR-188-3p on miR-188-3p expression and the promoting effect of ABCB1 vector on ABCB1 expression confirmed that both had a good transfection effect (Fig. 6A, B). Therefore, we co-transfected with si-circ_0109291 and anti-miR-188-3p or ABCB1 overexpression vector into CAL-27/DDP and UM1/DDP cells. CCK8 and colony formation assay results revealed that the inhibition effect of circ_0109291 silencing on the viabilities and colony numbers of CAL-27/DDP and UM1/DDP cells could be partially inverted by miR-188-3p inhibitor or ABCB1 overexpression (Fig. 6C–F). Besides, the reduction effect of circ_0109291 knockdown on the resistance of CAL-27/DDP and UM1/DDP cells to DDP also could be recovered by miR-188-3p inhibitor or ABCB1 overexpression (Fig. 6G, H). Through the detection of the apoptosis rate and the protein levels of Bcl-2, Bax, and Cleaved-cas-3, we uncovered that miR-188-3p inhibitor or ABCB1 overexpression also could partially reverse the promotion effect of silenced circ_0109291 on the apoptosis of CAL-27/DDP and UM1/DDP cells (Fig. 6I–L). Hence, we proved that circ_0109291 regulated the DDP resistance of OSCC cells by mediating the miR-188-3p/ABCB1 axis.
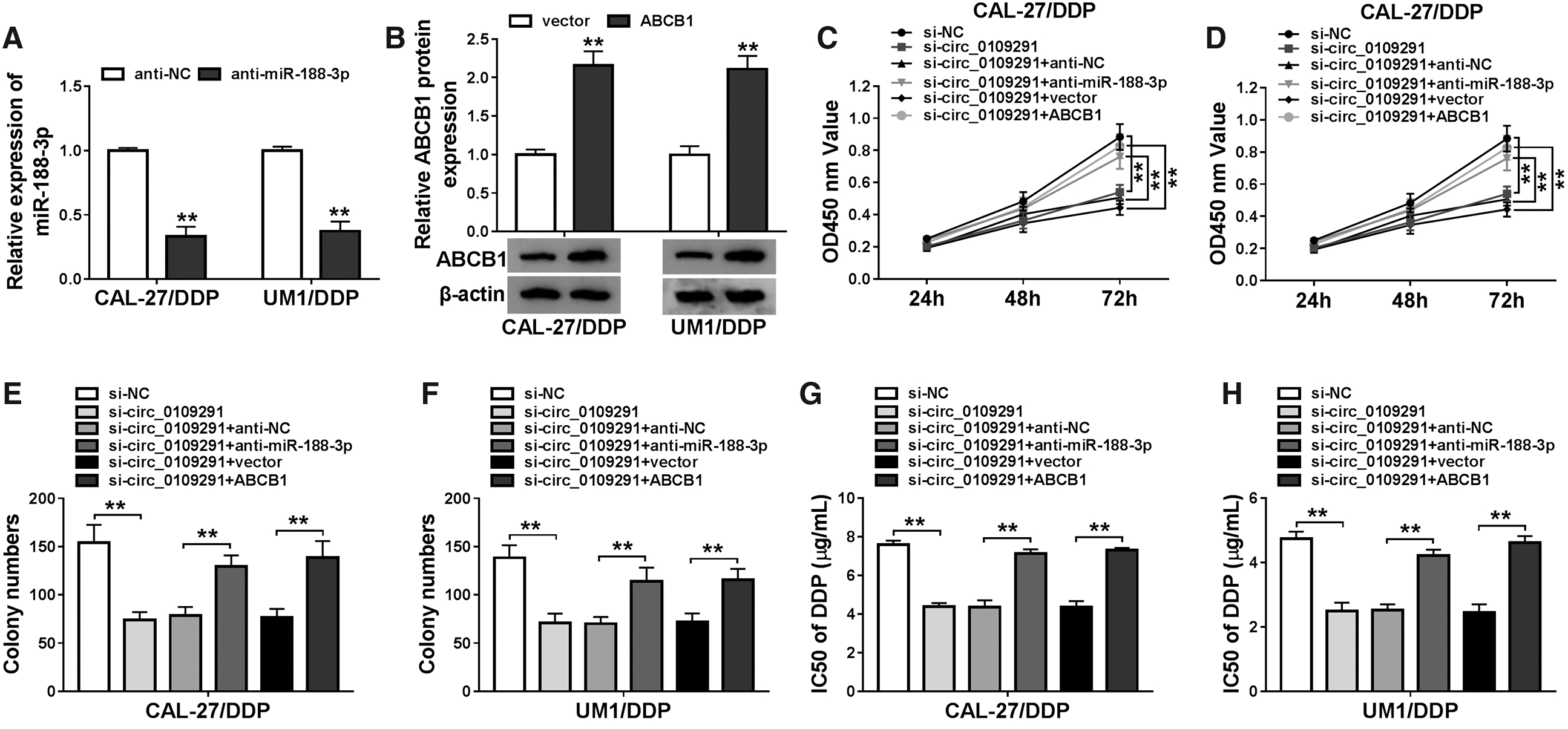
MiR-188-3p inhibitor or ABCB1 overexpression reversed the effects of circ_0109291 knockdown in the DDP resistance of OSCC cells.
Knockdown of circ_0109291 promoted the sensitivity of OSCC tumor to DDP in vivo
In addition, we built the mice xenograft models to confirm our results. As shown in Figure 7A, B, we found that DDP or circ_0109291 knockdown both significantly restrained the tumor volume and weight of OSCC, and circ_0109291 silencing also exacerbated the inhibitory effect of DDP on tumor volume and weight, suggesting that circ_0109291 knockdown might improve the DDP sensitivity of OSCC tumor. By detecting the expression of circ_0109291, we found that DDP could repress the expression of circ_0109291, and sh-circ_0109291 also had a significant inhibitory effect on its expression, confirming that the suppression of tumor growth was caused by decreasing circ_0109291 expression (Fig. 7C). Besides, we also measured the expression levels of miR-188-3p and ABCB1 in tumors and discovered that DDP and silenced circ_0109291 also could increase the miR-188-3p expression and decrease the ABCB1 expression, which was more obvious when DDP and sh-circ_0109291 acted simultaneously (Fig. 7D, E). Collectively, these findings showed that circ_0109291 might be an effective target to improve the sensitivity of OSCC to DDP.
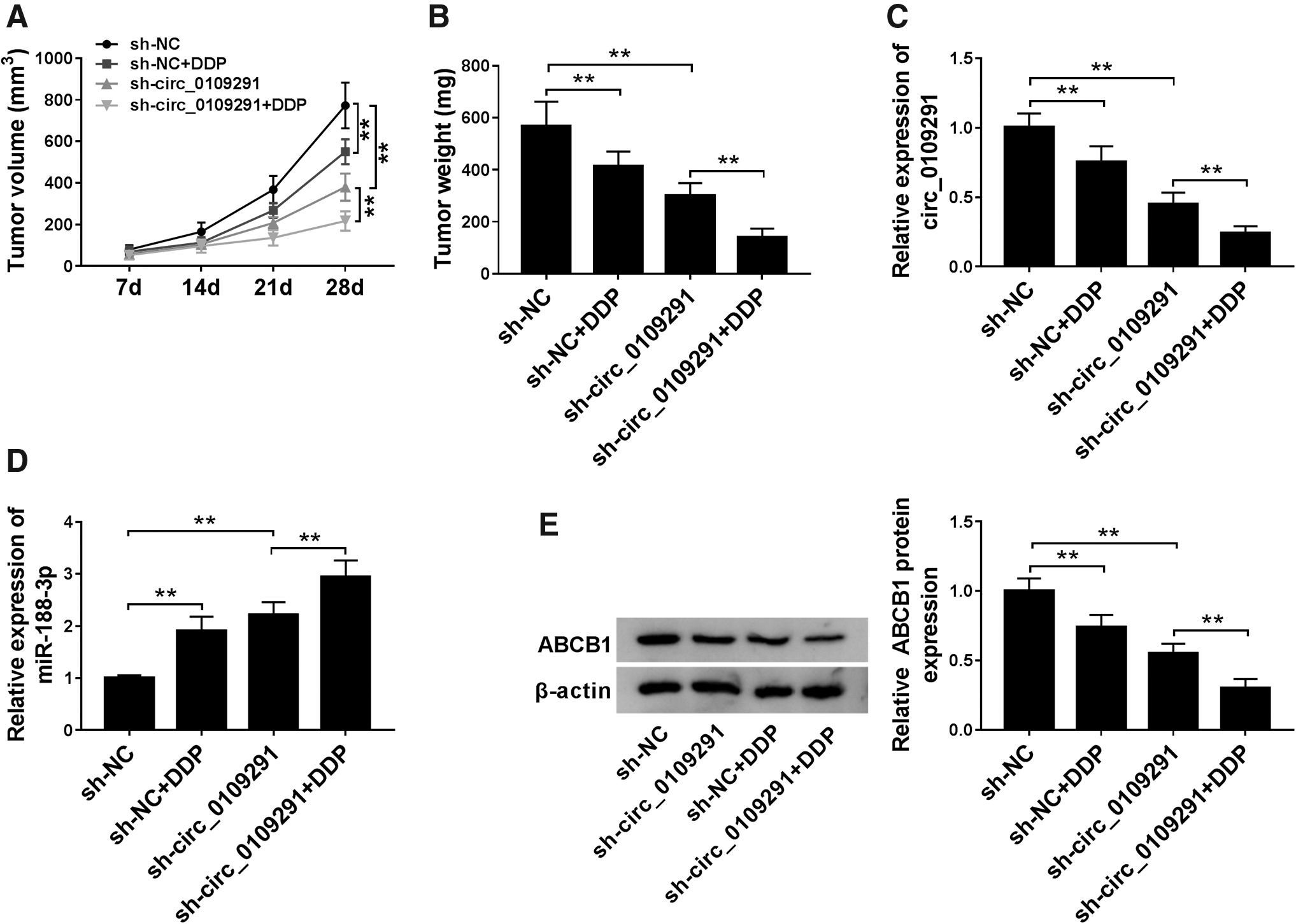
Knockdown of circ_0109291 reduced the OSCC tumor growth in vivo.
Discussion
Invasive OSCC with a poor prognosis often exhibits therapeutic resistance. 21 Therefore, it is of great significance to clarify the factors influencing drug resistance for cancer. The application of drug-resistant cells in exploring chemotherapy treatment has been widely concerned, and the function of circRNA in cancer progression has been well documented. However, few circRNA studies on OSCC drug resistance have been reported. Circ_0109291 was a newly discovered circRNA in recent years, and Ouyang et al. confirmed for the first time that it might be related to the proliferation, metastasis, and apoptosis of OSCC cells. 12 Here, we verified that circ_0109291 was upregulated in OSCC tissues and cells, especially in DDP-resistant OSCC tissues and cells. Through the loss-functional experiment, we confirmed the necessity of circ_0109291 expression for maintaining the DDP resistance of OSCC cells. In vivo experiments, we also demonstrated a significant increase in sensitivity of OSCC tumor to DDP with lower circ_0109291 expression. These data suggested that circ_0109291 might play an active role in the DDP resistance of OSCC.
In general, circRNA regulates tumor progression by a variety of molecular mechanisms, including acting as a sponge for miRNAs, regulating gene splicing or transcription, and mediating epigenetic regulation. 22 At present, more studies have been conducted on circRNA as a “miRNA sponge” that affects the expression of miRNA to indirectly regulating gene expression. 23 Zhao et al. showed that miR-188 could act as a tumor suppressor to regulate the tumorigenesis of non-small lung cancer. 24 Also, Li et al. suggested that miR-188 could inhibit the proliferation and cell cycle progression of glioma. 25 Here, we discovered that miR-188-3p had decreased expression in DDP-resistant OSCC tissues and cells, which was consistent with the expression trend in other diseases. 16,17 Besides, we also confirmed that miR-188-3p expression was negatively regulated by circ_0109291 in vitro and in vivo. MiR-188-3p overexpression had an inhibition effect on the DDP resistance of OSCC cells. Considering the opposite effect of circ_0109291 deletion and miR-188-3p overexpression on OSCC cells, we hypothesized that the pro-resistance of circ_0109291 might be related to the interaction of miR-188-3p. Further studies confirmed that the miR-188-3p inhibitor could partially reverse the inhibitory effect of circ_0109291 knockdown on the DDP resistance of OSCC cells. Therefore, our results suggested that miR-188-3p was a target for circ_0109291.
Many studies have confirmed that overexpression of ABCB1 is an important factor leading to multidrug resistance in tumor patients, so inhibition of ABCB1 expression is an effective way in reducing the drug resistance of cancers. 26,27 Li et al. showed that ABCB1 expression was associated with the acquisition of DDP resistance in human lung adenocarcinoma cancer cells. 28 Consistent with the previous study, 19,20 we suggested that ABCB1 was upregulated in DDP-resistant OSCC tissues and cells. Also, ABCB1 expression was regulated by circ_0109291 and miR-188-3p. The reversal effect of ABCB1 overexpression on si-circ_0109291 also verified that ABCB1 was a downstream target gene of circ_0109291. Similarly, the ability of circ_0109291 negatively regulating ABCB1 expression also had been demonstrated in vivo. Hence, our results elucidated the role of ABCB1 in OSCC resistance and perfected the mechanism of circ_0109291 regulating the DDP resistance of OSCC.
Conclusion
In general, our results suggested that circ_0109291 played a role in promoting the DDP resistance of OSCC, mainly by adsorbing miR-188-3p to regulate ABCB1 expression. Our conclusion had enriched the study of circ_0109291 in cancer research, which might provide a theoretical basis for reducing OSCC resistance occurrence.
Authors' Contributions
Conception and design: Fei Gao and Jianjun Han; Development of methodology: Yun Wang; Acquisition of data: Lin Jia; Analysis and interpretation of data: Wenjuan Luo; Writing, review, and revision of article: Yan Zeng; All coauthors have reviewed and approved of the article before submission.
Footnotes
Disclosure Statement
The authors declare that they have no financial conflicts of interest.
Funding Information
No funding was received for this article.