
Abstract
Targeted therapy is a fast evolving treatment strategy to reduce unwanted damage to healthy tissues, while increasing efficacy and specificity. Driven by state-of-the-art technology, this therapeutic approach is especially true of cancer. Antibodies with their remarkable specificity have revolutionized therapeutic strategies for autoimmune conditions and cancer, particularly blood-borne cancers, but have severe limitations in treating solid tumors. This is mainly due to their large molecular size, low stability, tumor-tissue penetration difficulties, and pharmacokinetic properties. The tumor microenvironment, rich in immune-suppressing molecules is also a major barrier in targeting solid tumors by antibody-based drugs. Nanobodies have recently emerged as an alternative therapeutic agent to overcome some of the drawbacks of traditional antibody treatment. Nanobodies are the VHH domains found on the heavy-chain only antibodies of camelids and are the smallest naturally available antibody fragments with excellent antigen-binding specificity and affinity, equivalent to conventional antibodies but with molecular weights as low as 15 kDa. The compact size, high stability, enhanced hydrophilicity, particularly in framework regions, excellent epitope interactions with protruding CDR3 regions, and improved tissue penetration make nanobodies the next-generation therapeutics (Nano-BioDrugs). In this review, the authors discuss the interesting properties of nanobodies and their advantages over their conventional counterparts and provide insight into how nanobodies are being utilized as agonists and antagonists, bispecific constructs, and drug and enzyme-conjugates to combat the tumor microenvironment and treat disease.
Introduction
Targeted therapy is a promising strategy to combat cancer without damaging healthy cells and tissues. Monoclonal antibodies (mAbs) are the most widely used and successful targeted therapy against a wide range of cancers, showing great specificity towards tumor-associated antigens (TAA) and neoantigens, but with little impact on solid tumors. 1 Antibodies are heterodimeric globular glycoproteins, secreted by B lymphocytes and are composed of two heavy chains and two light chains with variable and constant domains linked by inter- and intrachain disulfide bonds 2 (Fig. 1A). The variable domains of the light and heavy chains form the antigen-binding regions, whereas the constant domains—CH2 and CH3 of the heavy chains form the Fc region. The Fc region controls effector functions such as antibody-dependent cellular cytotoxicity (ADCC), antibody-dependent cellular phagocytosis, antibody-mediated complement-dependent cytotoxicity (CDC), antibody recycling, and other immunomodulatory functions via interaction with Fc receptors. 3 The unrivalled specificity of antibodies and their high affinity toward specific targets/antigens make them excellent therapeutic molecules (BioDrugs).
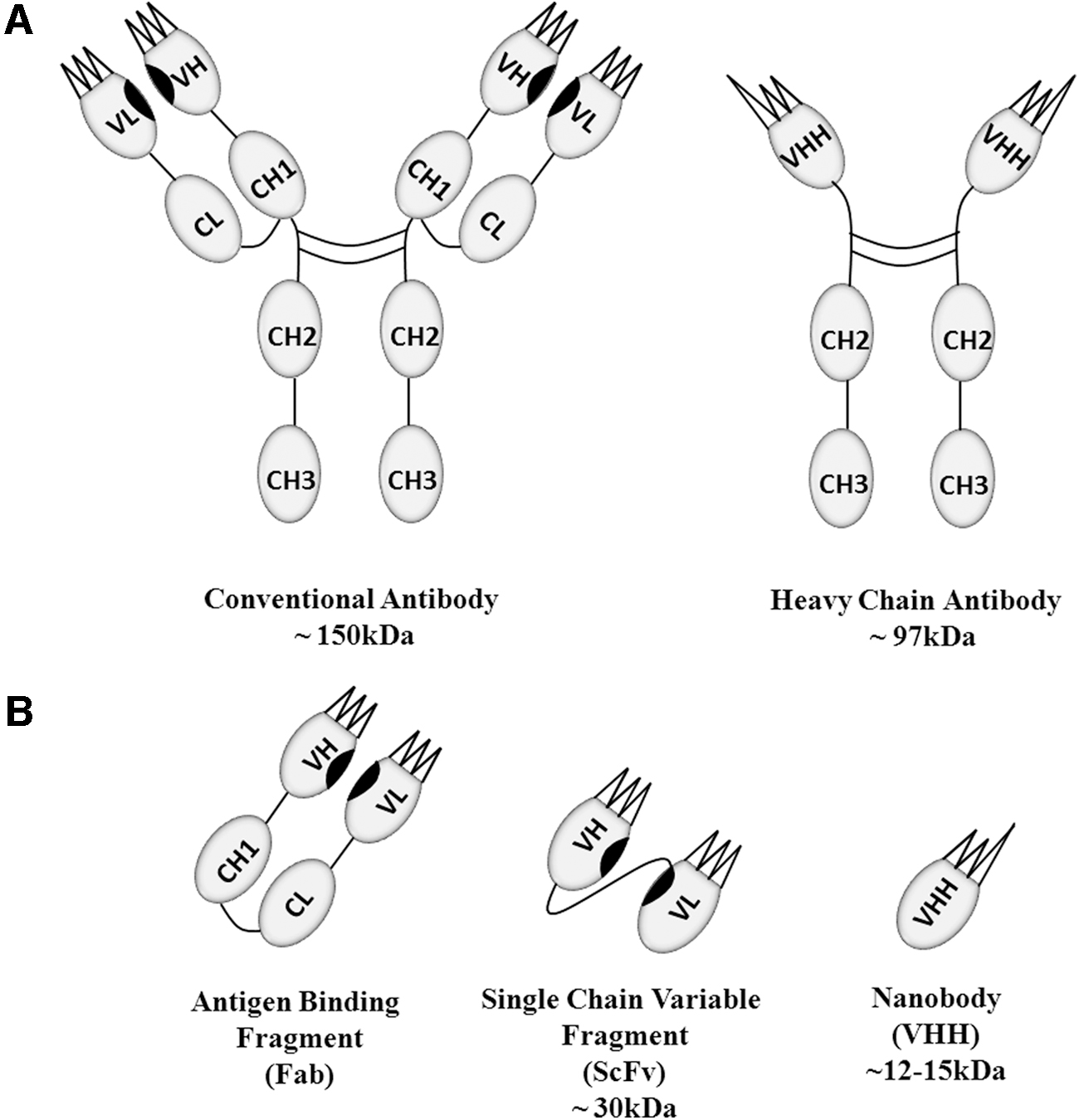
The conventional IgG antibody versus the heavy-chain (camelid) antibody.
Through recombinant technologies and genetic manipulation, the antibody structure has been engineered in many ways to develop different variants and constructs, including bispecifics, triabodies, tetrabodies, scFv, Bis-scFv, and minibodies, with also the addition of chemotherapeutic drugs to form antibody-drug conjugates. 4 Blockbuster antibody-based drugs such as Rituxan (rituximab) and Darzalex (daratumumab) have demonstrated excellent cancer remission and survival rates in patients with blood cancers, thereby revolutionizing cancer treatment strategies for these conditions. 5 However, the large size, low stability (such as frequent denaturation, aggregation), 6 immunogenicity, large doses required, and difficulties with tissue and tumor penetration, 1 together with high manufacturing costs and development of anti-drug antibodies and neutralizing antibodies (Nabs), have limited the effectivity and range of antibody-based treatments for cancer. These limitations have led to the development of novel antibody therapeutics to improve clinical outcome.
The Single-Domain Antibody
The discovery of heavy-chain-only therapeutic antibodies is marked by the seminal studies of Raymond Hamers et al. on the dromedary camel immune system in 1989. 7 The heavy-chain-only antibody and conventional antibody differs in the light chains; the heavy-chain-only antibodies are devoid of the variable and constant light chain along with the CH1 domain 8 (Fig. 1A). A similar type of heavy-chain-only antibody was discovered in cartilaginous fishes such as sharks; however, these heavy-chain-only antibodies known as IgNAR antibodies have five constant (CH) domains 9 and are challenging to engineer and develop due to the difficulty in immunizing sharks. Conventional antibodies contain six complementary determining regions (CDR) on the heavy and light chains, whereas the heavy-chain-only antibodies possess only three CDRs (Fig. 1B). Despite only possessing three CDRs, these heavy-chain-only antibodies show equivalent antigen-binding affinity and other effector functions compared to conventional six CDR antibodies. 10 This property has driven immense interest in the variable region (VHH) of heavy-chain antibodies as therapeutics. 11
The Nanobody (VHH Domain)
The VHH domain is the antigen-binding fragment of heavy-chain-only antibodies and the smallest fragment of an antibody with antigen-binding capability, comparable with conventional heavy- and light-chain antibodies. Due to the lack of the CH1 domain, the VHH is directly connected to the CH2 domain through a hinge in the heavy-chain-only antibody. The diminutive size of a monomeric VHH domain antibody augments its efficacy with improved penetration to target tissues. 12 It is composed of only ∼120 amino acids with a molecular weight of ∼12–15 kDa (1–10th the molecular weight of a conventional antibody) and with dimensions of ∼4 × 2.5 × 3 nm. 13,14 In addition, unlike murine antibodies, most of the VHH domain shows 80% homology with human domains. 15,16 Crystallography studies revealed many dynamic features of the VHH domain, such as it is composed of four framework regions (FR) with nine β-sheets separated by loops, which include three hypervariable regions. 17 In general, three hypervariable regions (CDRs) and four framework regions comprise the variable region, of which the framework regions occupy 85% and act as a scaffold to the CDRs. 18 The VH-VL interacting regions, VH-CH1 contact regions, and the hydrophobic interface of VH domains are replaced with amino acid substitutions in VHH domains, particularly in the framework 2 region (FR2) of VHH. Sequence analysis revealed amino acid substitutions at four places in FR2: at Val37 to Phe or Tyr, Gly44 to Glu, Leu45 to Arg or Cys, and Trp47 to Gly. All of these amino acid substitutions contribute to the enhanced solubility and increased structural variability of nanobodies. 19
The VHH domain antibody, with only three CDRs has notable differences compared to the conventional VH domain, such as the difference in the upper core packing (buried CDR residues and a number of N-terminal residues), 20 due to different CDR1 and CDR2 conformations. 21 This loose packing in the upper core enables the VHH to adopt a significant variety of shapes, thereby enhancing its binding diversity and also aiding in the establishment of a stable VHH-Ag complex. 22 The CDR1 located toward the N-terminal region has high variability, 16 but nonetheless contributes to the binding strength of the domain in collaboration with CDR2, particularly by formation of an interloop disulfide bond with CDR3. 23 The CDR3 is mainly engaged in antigen recognition 24,25 and is longer than in the VH domain; it is made up of ∼14–18 amino acids, 26 whereas the human and murine CDR3 is composed of 12 and 9 amino acids, respectively. 27 This longer CDR3 gives increased flexibility for antigen binding and specificity variations. Furthermore, in many nanobodies, CDR3 domains have protruding ends, which improves their ability to reach and interact with epitopes located in protein crevices and enzyme-active domains. 28 All of these properties contribute to the convex surface of VHH, which supplements its ability to interact with the antigen cavities. 29 Therefore, nanobodies with a convex paratope surface most probably bind to concave surfaces. 30 The intriguing properties of nanobodies have further uses such as in helping to solubilize poorly soluble proteins for crystallization and to hinder their domain flexibility. 31 The protruding ends can also make nanobodies excellent inhibitors or modulators of proteins, enzymes, receptors, or ion channels. 32,33
A key feature of VHH domains is their stability and their ability to remain stable in the presence of degrading enzymes and other harsh conditions such as low pH and detergents. They exhibit unchanged activity even after subjecting them to thermal degradation up to 90°C. 34 The conventional VL-VH domain, which comprises the hydrophobic interface with antigen binding sites, becomes denatured when subjected to high temperature and does not refold back to the original active conformation and forms aggregates. In contrast, the VHH domains demonstrate excellent refolding capability and with minimal loss in activity. The melting temperature (refolding) of llama VHH was identified as ∼60°C by Perez et al. (2001) 35 and a number of other studies demonstrated the melting temperature to lie between 60.1°C and 66.7°C. 22
One of the most desirable properties of nanobodies as therapeutics is their dynamic penetrating ability into tissues not normally accessible by conventional antibody therapeutics. In a study conducted by Pierre Lafaye et al. (2012), it was shown that nanobodies are capable of crossing the blood–brain barrier (BBB), 36 where most proteins, including antibodies, cannot and can be used as a shuttle to deliver drugs to targeted cells within the central nervous system. 37 This has intriguing implications for the treatment of neurological conditions, including cancer. The same group also showed the ability of nanobodies to induce general and receptor-mediated transcytosis, which also has important implications for drug delivery. 38 A further important property in the application of nanobodies in drug delivery is their ability to access intracellular protein targets by penetrating cells. It was shown that an E9 nanobody developed against the intracellular glial fibrillary acidic protein (GFAP), effectively penetrated astrocytes and bound to GFAP. 36 Besides many positive features, one of the downsides of nanobodies is their fast pharmacokinetics (quick renal elimination from blood stream). 39 This drawback can be overcome by multiple approaches such as covalent attachment of nanobodies with polyethylene glycol (PEGylation), fusion to the IgG Fc region, and by developing bispecific nanobodies that target serum albumin. 40 –42 Some examples of successful nanobodies currently in clinical trials are listed in Table 1.
Therapeutic Nanobodies in Clinical Trials
MMR, macrophage mannose receptor; TNF, tumor necrosis factor.
Nanobodies as Cancer Therapeutics
Advances in cancer immunology have paved the way for many novel therapeutic strategies, including antibodies and nanobodies and their ability to detect tumorigenic cells and activate effector responses of the immune system. A number of these ground-breaking immunotherapeutic approaches to cancer include identification and therapeutic deactivation of checkpoint inhibition, activation of antigen-presenting cells (APCs), blocking of immune-suppressive cells, and neutralization of immune suppressive molecules, development of chimeric antigen receptor T cells (CAR T cells) and CAR-natural killer (NK) cells, cytokine modulation, and others.
Nanobodies and T Cells in Cancer Immunotherapy
T lymphocytes and their subsets are key cells in the immune system response to cancer. They exhibit antitumor activity directly and indirectly through secretion of granzymes and perforins by cytotoxic T lymphocytes and activation of other cancer targeting immune cells. Tumorigenic cells and the tumor microenvironment therefore target T lymphocytes to inactivate them and suppress the immune response, a major survival mechanism for cancer cells. Many therapeutic approaches involving nanobodies have been used to counteract this inactivation and suppression. One particularly promising approach is the use of bispecific antibodies in combination with checkpoint inhibitors to link T cells to tumor cells and at the same time prevents immune suppression. This strategy has shown promising improvements in tumor mass reduction. 43 Currently, three types of bispecifics exist with specific benefits; the bispecific T cell engager (BiTE), dual-affinity retargeting proteins (DARTs), and tandem diabodies (TandAbs). 44 BiTE antibodies were developed to bridge T cells with tumor cells and are composed of two scFv fragments, one against an antigen/marker of T cells such as CD3 and the other against a TAA, coupled with a peptide linker 45 (Fig. 2A). Blinatumomab is a Food and Drug Administration (FDA)-approved BiTE developed to treat B cell acute lymphoblastic leukemia (B-ALL) with specificity for CD3 of T cells and CD19 of B cells (overexpressed in ALL), connected with a glycine-serine linker. 46 This antibody demonstrated good efficacy, but a short half-life, a major problem with blinatumomab as well as other BiTEs and requires high therapeutic dosage to reach required T cell activation. 47 DARTs have been developed as an alternative to overcome the short half-life of BiTEs and to tackle quick elimination. Unlike BiTEs, DARTs have heterodimerized Fv fragments, with the VL of target A and VH of target B on Fv1 fragment and the VL of target B and VH of target A on the Fv2 fragment 48 (Fig. 2B). These combinations mimic the natural IgG molecule and have increased the half-life/serum retention of these antibody molecules from 1.25 ± 0.63 h (BiTE) to ∼10 h (DARTs). 49,50
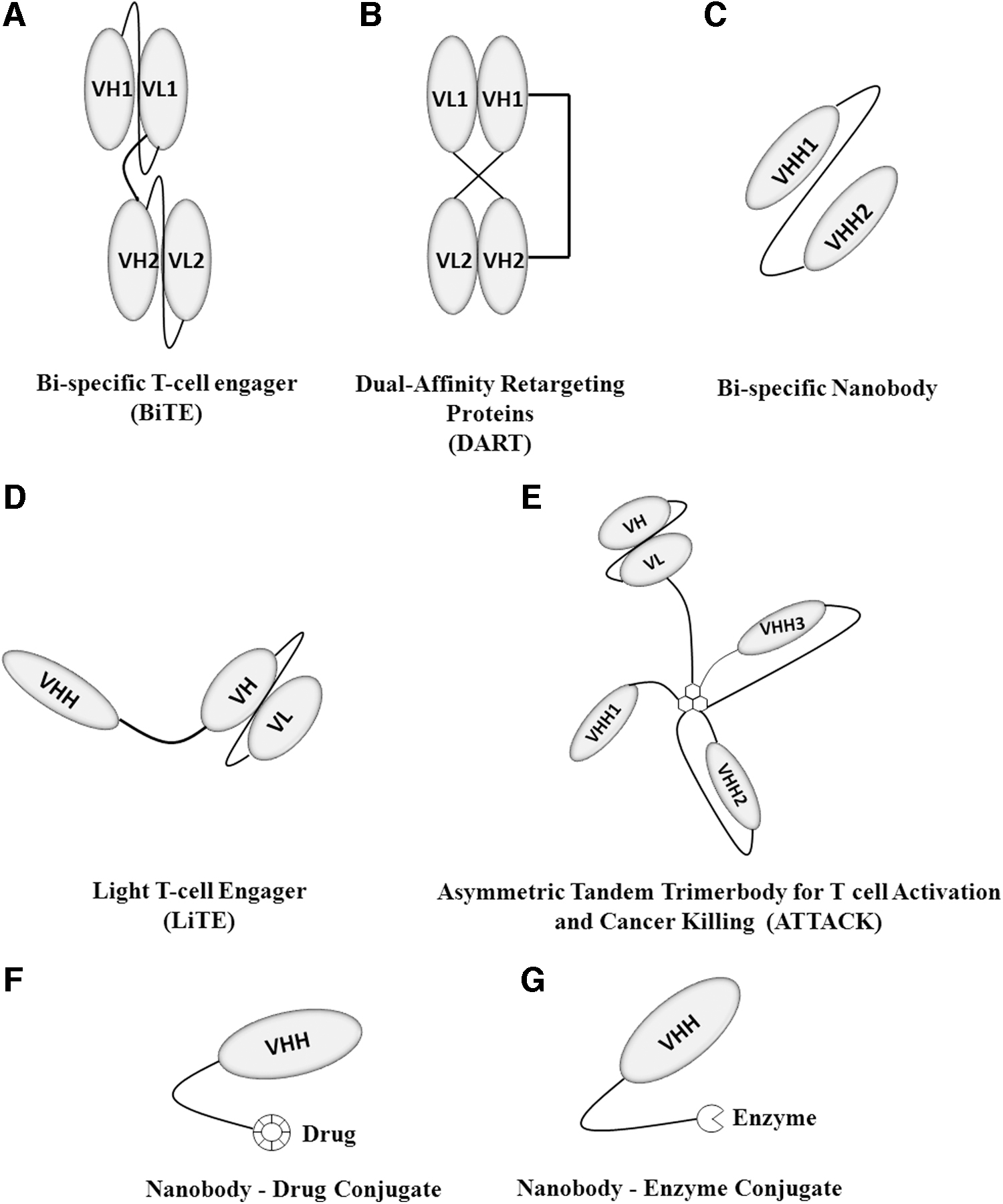
Therapeutic Antibody and Nanobody constructs. These are a wide range of novel therapeutic constructs developed against various cancers, which have proved to be highly successful and are discussed extensively in this review.
Bispecific nanobodies, with significantly higher stability, solubility, and improved properties would be expected to also present enhanced efficacy compared to conventional BiTEs. Recently, a bispecific antibody against CD3 of T cells and Her2 of breast cancer cells was developed. The uniqueness of this therapeutic is the combination of the single-domain (VHH) specific to Her2 and scFv-fragment specific to CD3. 51 This construct enabled easy expression in E.coli and purification by affinity chromatography. A similar construct was developed by Mølgaard et al. (2018), who termed it LiTE (Light T cell Engager) where a CD3-specific scFv fragment and two antiepidermal growth factor receptor (EGFR) VHH domains were developed to bind two molecules of the EGFR. Two LiTE constructs were developed by fusing the nanobody at the N- and C-terminal domains of scFv 52 (Fig. 2D). There was no significant difference in the efficacy of these antibody constructs, where both exhibited satisfactory tumor-specific cytolysis with excellent tissue penetration and low cytotoxicity. 52 In addition, ATTACK (Asymmetric Tandem Trimerbody for T cell Activation and Cancer Killing) molecules have been generated into novel bispecifics. Here, T cell CD3-specific scFv was fused with three anti-EGFR nanobodies to create a construct that displayed enhanced specificity toward EGFR with a 15-fold increase in efficacy 53 (Fig. 2E). Similarly, bispecific-trivalent constructs and other different antibody constructs are being developed; all which confirm the potential of nanobodies with improved tumor localization and lower off-target toxicity and side-effects. 54 Table 2 lists a number of bispecific nanobodies in development.
Bispecific Nanobodies in Preclinical Development
EGFR, epidermal growth factor receptor.
γδ T Lymphocyte
The γδ T lymphocyte is present in low abundance, but plays a pivotal role in antitumor activity. This small subset of T cells possesses many unique properties in the immune response; they do not need major histocompatibility complex (MHC) to become activated; are stimulated by stress antigens: DNAM-1 (DNAX accessory molecule-1), CD27 costimulatory receptor, and they exhibit both innate and adaptive immune features. 55 –57 They execute target cell killing by recognizing tumor necrosis factor (TNF)-mediated apoptosis, secretion of granzymes and perforins, ADCC, and other means. 58,59 They also function as APCs, prime CD4+ T cells, and cross-present antigen to stimulate CD8+ T cells. 60,61 There are subsets of γδ T lymphocyte—the regulatory γδT and γδT17 cells that promote tumor growth, whereas the Vγ9Vδ2 T cells inhibit tumor growth and have different activation molecules and pathways, which allow them to be targeted therapeutically. 62,63 Numerous nanobodies have been developed against the γδ T cell; Bruin et al. (2016) raised 20 nanobodies against the T cell receptor (TCR) of Vγ9Vδ2 cells and selected two, which were specific to Vδ2+ and Vγ9+. 64 Random activation of Vγ9Vδ2-T cells results in low efficacy with a systemic response rather than a tumor-focused response. To address this, a bispecific nanobody targeting the EGFR (overexpressed on tumor cells) and Vγ9Vδ2 T cells was engineered, thereby inducing a tumor-specific response (cell lysis) while avoiding the consequences of systemic Vγ9Vδ2 T cell activation. 65,66 In conditions such as metastatic bone disease or hypercalcemia with amino bisphosphonates, the activation of Vγ9Vδ2 T cells can have negative effects and exacerbate the disease condition. 67 In this situation, preventing the activation of Vγ9Vδ2 T cells therapeutically is desirable but no such clinically approved biologics currently exist. A VHH domain against the TCR of Vγ9Vδ2 T cells was also identified, which blocked the phosphoantigen-independent and -dependent activation and controlled the production of proinflammatory cytokines in response to amino bisphosphonates. 67
NK Cells
NK cells are innate lymphoid cells, which have a pivotal role in antiviral and antitumor immune responses. They do not need MHC presentation for activation and utilize the activate and inhibit (kill and don't kill) signal through many receptors such as CD94/NKG2A, KIR, natural cytotoxicity receptor NKp46, NKp30, NKp44, IgG receptor FcγRIII, and many other C-type lectin-like receptors. 68 –71 A large variety of nanobody constructs have been developed to target FcγRIII (CD16) of NK cells. Two camelid nanobodies (C21 and C28) were generated in llama against the extracellular domain of FcγRIII with significant affinity for FcγRIIIb+ poly morphonuclear cells and FcγRIIIa+ NK cells. 72 These nanobodies when biotinylated and incubated with streptavidin induced significant secretion of IL-2 and interferon gamma (IFN-γ) in Jurkat–huFcγRIIIA/γ cells (engineered immortalized T cells) and NK-cells and presented an alternative approach to activate NK cells against tumor cells. Muc-1 is a well-studied tumor antigen associated with breast and ovarian cancer. 73 Muc1-Bi-1 is a bispecific nanobody developed to target Muc-1 and CD16a, and in in vivo studies Muc1-Bi-1 demonstrated good tumor growth suppression, especially in the presence of peripheral blood mononuclear cells. 74 To reduce immunogenicity, the Muc1-Bi-1 nanobody was humanized by CDR grafting, and to reduce quick elimination and improve the half-life, it was conjugated with PEG and similar compounds. 74 Similarly, a bispecific was developed against carcinoembryonic antigen (CEA) and CD16, this nanobody showed good remission in tumor mass with low or zero cytotoxicity in CEA-positive tumor cells, 75 CEA is a glycoprotein antigen highly expressed in many cancers. 76
Nanobodies as Checkpoint Inhibitors
Immune checkpoints are vital mechanisms for regulating the potential damage to normal cells by effector cells. Tumor cells, however, have exploited this mechanism to suppress the immune response and establish the tumor microenvironment. The checkpoint receptors CTLA-4, LAG-3, TIM-3, PD-1, and KIR are mainly found on T-and NK-cells 77,78 and numerous mAbs and small molecules have been developed to modulate the activity of these checkpoints. Despite the benefits, conventional antibody checkpoint inhibitors have drawbacks, such as low binding affinity, penetration, stability, and production. 79 A nanobody designated KN035-Fc raised against the PD-L1 receptor, with fused Fc fragment of human IgG, was shown to have improved affinity (∼1000-fold) for PD-L1 over PD-180 and KN035-Fc upon interacting with PD L-1, reduced its flexibility and blocked the formation of PD-1 and PD-L1 complex. 81 KN035 binds to the PD-L1 receptor through a loop of 21 amino acids by hydrophobic and ionic interactions with Ile54, Tyr56, and Arg113 residues. 80,81 Similarly, a nanobody known as sdAbK2 was developed against PD-L1, which has a dual role in diagnosis (nuclear imaging), as well as a therapeutic agent with a binding dissociation constant of 3.75 nM, equivalent to avelumab, an anti-PD-L1 mAb. 82 This nanobody also showed excellent performance when labeled with technetium-99m for diagnosis and screening of PD-L1-elevated breast cancer, melanoma, and kidney cancer with high signal to noise ratio. 83
A ground breaking approach to synthesize and deliver a checkpoint inhibitor antibody/nanobody using a probiotic bacteria was developed to overcome toxicity issues with conventional checkpoint inhibitors. 84 Advances in synthetic biology facilitated the development of genetically engineered bacteria with genes for desired therapeutic proteins and a lysis circuit operon (Synchronized Lysis Circuit). 85 These bacteria are capable of growing and replicating until they reach a threshold bacterial population density and undergo cell lysis releasing the synthesized therapeutic 86 and can therefore be used as a drug delivery vehicle. Similarly, Gubatri et al. 2020 developed a probiotic E.coli Nissle 1917 system, expressing genes for dynamic lysis and for nanobodies against PDL-1 and CTLA-4. 87 This system, when tested in syngeneic mice, demonstrated a significant systemic immune response with enhanced T cell activation. 87 TIM-3 (T cell immunoglobulin and mucin domain 3) is also an important checkpoint inhibitor targeted during cancer therapy. A novel nanobody raised against this receptor displayed significant antiproliferative activity in an acute myeloid leukemia cell line in vitro. 88 It should be noted, however, that manipulating systemic immune checkpoints can cause significant toxicities and off-target effects, that requires expert management.
Nanobodies and Cytokines
Cytokines have a significant impact on tumor growth and survival and tumor-promoting cytokines coexist along with tumor-suppressing cytokines and also cytokines with dual properties. 89,90 These are key proteins of the immune system and ultimately determine the immune response. The cytokine TGF-β acts as a tumor suppressor by arresting the cell cycle and inducing apoptosis, but gradually in the late stages of tumor growth, it promotes epithelial-mesenchymal transition, tumorigenesis, metastasis, and chemoresistance. 91 TNF-α is a potent proinflammatory cytokine that promotes tumor growth by lymphangiogenesis and lymphatic metastasis in a VEGFR3 independent mechanism. 92 Similarly, many other cytokines influence tumor suppression and progression and the antitumor immune response can be modulated by targeting these proteins.
A novel anti-TNF-α nanobody was developed in Pichia pastoris with a molecular weight of 15 kDa and 2.05 nM affinity, which showed a substantial reduction in proliferation, migration, and invasiveness of cancer cells induced by TNF-α in the breast cancer cell lines: MCF-7 and MDA-MB-231. 93 When this nanobody treatment was complemented with paclitaxel treatment, there was improved efficacy of the chemotherapeutic drug and a decrease in resistance by the cancer cells. 93 CXCR7 is a chemokine receptor for CXC11 and CXC12, which play a significant role in angiogenesis and vascular formation, thereby facilitating metastasis in breast, lung cancer, and other tumors. 94 David et al. (2013) identified elevated levels of CXCR7 in head and neck tumors, and developed CXCR7 antagonist nanobodies, which prevented the recruitment of β-arrestin 2 and showed substantial reduction in head and neck tumors by lowering the secretion of the angiogenic chemokine (CXCL1). 95 Similarly, the cytokine TRAIL (TNF-Related Apoptosis-Inducing Ligand) is a tumor-specific death inducer, which executes its downstream antitumor effects by interacting with death receptors. 96 Antibody constructs have been developed since the mid-1990s to target TRAIL in anticancer therapeutics. 97 TAS266 is a tetravalent death receptor 5 (DR5) agonist nanobody developed by Novartis, which demonstrated significantly higher activation of DR5 and tumor cell death than TRAIL and the conventional DR5 Mab. 98 A side-effect observed, however, was elevated hepatotoxicity due to unexpected immunogenic responses (reversible). 99
Antigen-Presenting Cells
A T cell response is stimulated by APCs such as dendritic cells (DC). A DC-specific anti-CD11b nanobody conjugated with the H-2Db (MHC haplotype)-restricted immunodominant E7 epitope (E749–57) was recently developed to treat human papillomavirus (HPV)-induced cancers, such as cervical cancers. 100 E7 and E6 are viral oncoproteins associated with HPV-induced cervical and oral cancers and inhibit the activity of the tumor suppressor genes p53 and RB1. 101 Administration of the E749–57 epitope conjugated nanobody demonstrated augmented activation of E7-specific49–57 CD4+ and CD8+ T cells by APCs and substantial tumor regression than E749–57 treatment alone, there was also increased antigen cross-presentation and a boosted CD8+ T cell response when the CD11b-specific nanobody was conjugated with H-2Kb(MHC haplotype)-restricted ovalbumin peptide (OVA257–264). 100,102
The mucin MUC1 is a type-I transmembrane glycoprotein widely found on tumor cells, which can hinder chemotherapeutic drug interactions, prevent cell death, and promote tumor invasion. 103 MUC1 is therefore an effective drug target to induce an anticancer immune response. An MHC class II-specific VHH7 nanobody conjugated with the MUC1-Tn antigen showed dynamic antigen presentation of MUC1 by DC and B cells and triggered a robust immune response against αMUC-Tn. 104 Activated DCs and B cells secreted cytokines and immunoglobulins that promote CDC and Th1-lymphocyte-mediated tumor cell death. In addition, immune memory provided by a single boost following nanobody treatment lasted for a period of 12 months. 104
Nanobody-Drug Conjugates
Chemotherapy is the use of drugs/chemicals to kill tumor cells, however, the challenge with this approach and to a lesser extent radiotherapy is off-target effect, which leads to acute toxicity in healthy cells. The combination of a targeted therapy such as an antibody/nanobody with a nontargeted therapy such as chemotherapy/radiotherapy was an advance on previous approaches. Fang et al. (2016) used a VHH7 domain (against MHC class-II) as a therapeutic agent, as well as an imaging tool for B cell lymphoma. 105 The VHH7 domain, when conjugated with the thio-containing maytansine derivative mertansine (DM1) and used as a therapeutic, demonstrated excellent specificity, internalization, and cytotoxicity in a murine model of A20 B cell lymphoma, where it resulted in a decrease in tumor growth and metastasis 105 (Fig. 2F). The same VHH7 domain, when conjugated with near infra-red (NIR) fluorophores, exhibited excellent diagnostic properties in visualizing both localized and metastatic tumor cells. 105 Similarly, the combination of targeted radionuclide therapy with nanobodies is gaining huge attention as new targeting constructs. Vaneycken et al. (2011) generated 38 nanobodies against HER2 from an immunized dromedary, and 2Rs15d-nanobody was selected as the lead compound after many in vivo and in vitro studies on tumor uptake and tumor-to-background ratios. 106 2Rs15d-nanobody was further exploited to develop several nanobody conjugated radioactive therapeutics, including an anti-HER2 2Rs15d radiolabeled with 225Ac (225Ac-DOTA-Nb), anti-HER2 2Rs15d labeled with 211At (211At-SAGMB-2Rs15d), and anti-HER2 2Rs15d labeled with 177Lu (177Lu-DTPA-2Rs15d); 107 –109 all, when tested for antitumor activity in mouse models, demonstrated excellent antitumor activity with favorable biodistribution and minimal toxicity (preclinical) 107 –109 ; 131 I labeled 2Rs15d-nanobody ([ 131 I]SGMIB-2Rs15d) successfully completed phase 1 clinical trials (NCT02683083) as a theranostic drug for HER2+ breast cancer. 110 Photothermal therapy is an emerging approach to induce tumor cell death by combining photosensitizers (nanoparticles) and laser light. These nanoparticles generate heat and reactive oxygen species when absorbed and excited by lasers (NIR), and induce cancer/tumor cell death. Branched gold nanoparticles are one such photosensitizer, which showed promising results in killing cancer cells. 111 Van de Broek et al. (2011) developed a nanobody conjugated with branched gold nanoparticles against HER2. These nanobodies showed high binding affinity for elevated HER2 on breast and ovarian cancers and destroyed HER2-positive SKOV3 cells within 5 min of irradiation. 112 These nanobodies may present as an effective targeted photothermal therapy. Photodynamic therapy is a similar approach where tumor cell death is induced by singlet O2 produced upon irradiation. 111 In light of this, a nanobody conjugated with IRDye700DX (photosensitizer) against EGFR was developed and antitumor activity was examined in an orthotopic mouse with 90% tumor cell necrosis 1 h postinjection. 113
Nanobody enzyme-conjugates are another nanobody-based approach, which have shown promising activity in tumor elimination. Cortez-Retamoza et al. (2004) developed a nanobody enzyme-conjugate, cAb-CEA5:βL to target tumor-associated CEA 114 (Fig. 2G). This nanobody was conjugated with the Enterobacter cloacae β-lactamase enzyme, which can site-specifically activate the antitumor prodrug 7-(4-carboxybutanamido cephalosporin mustard into the active drug phenylenediamine mustard. 115 Conjugation was performed using a llama γ2c hinge to provide flexibility and stability and showed rapid clearance from systemic circulation and concentration in tumors, thereby avoiding off-target toxicity. 114
Nanobody-Based CAR T Cell Therapy
CAR T cell therapy is a state-of-the-art technology, where T cells are genetically engineered with an artificial TCR. The engineered TCR comprises an extracellular antigen recognition domain, usually an scFv fragment, a hinge region, a transmembrane domain, and a signal transduction domain with stimulatory (CD3ζ) and costimulatory (CD28, OX40, CD27) molecules. 116,117 This construct facilitates TAA recognition and activation of T cells in an MHC independent manner, where the antigen can be a peptide as well as a nonpeptide. 118,119 Many CAR T cell therapies have been generated against CD19, CD20, and CD30 for treatment of chronic lymphocytic leukemia, B-ALL, Hodgkin's lymphoma (HL), and B cell non-HL. Targeting CD19 with CAR T cells showed the most promising therapeutic results against B-ALL with complete remission rates of 70%–94%. 120,121 To date, Kymriah (anti-CD-19 CAR T cell) and Yescarta (anti-CD19 CAR T cell) are two FDA approved CAR T cell therapies for certain types of non-HL and B-ALL. 122 Advances in CAR T cell therapy are progressing from blood cancers to solid tumors and in addition CAR-NK cell therapy has been developed to overcome drawbacks with T cell therapy. Recently a large number of nanobody-based CAR T cell therapies have been developed and have largely outperformed conventional antibody-based CAR T cell therapies. 123
The first nanobody-based CAR T cell therapy was introduced in 2018 with the VHH domain against MUC-1 and CD28 and OX40 as the costimulatory molecule. 124 The killing of MUC-1+ tumor cells by this CAR T cell was observed in tumor xenografts in combination with increased production of the proinflammatory cytokines IL-17 and IFN-γ. 125 This NanoCAR system was further enhanced with a suicidal gene system to avoid unwanted damage to healthy cells and control unwanted replication. In this system a homodimerization-regulated caspase 8 was used, which was activated by the synthetic ligand AP20187 to induce apoptosis. 126 A significant challenge with treating solid tumors, in addition to the tumor microenvironment is tumor-specific target identification. To address this, researchers at MIT and Harvard developed nanobody-based CAR T cells that target the general tumor microenvironment, particularly PD-L1 and EIIIB, which are present on multiple tumor types. 127 These nanoCAR T cell therapies demonstrated excellent identification and binding activity in many tumors and when activated also secreted IL-2 and IL-12, which increased inflammation in tumors. 128
Bispecific nanobody-based Car T cell therapies have also been developed, as relying on a single antigen for tumor identification is generally insufficient and leads to treatment failure with antigen-negative tumors. In this therapeutic approach De Munter et al. (2018) developed two NanoCARs against CD20 and HER2; the antigen-binding domain composed of two VHH tandem domains targeting two different antigens on tumor cells. 129 This NanoCar approach showed promising potential in tumor elimination by overcoming the problem of antigen loss on tumors. In addition, a nanobody-based UniCAR against EGFR+ tumors was developed. 130 Unlike the conventional CAR, the activity of this UniCAR can be modulated and targeted to tumor-specific antigen by recombinant targeting module epitopes, thereby eliminating unnecessary risks such as cytokine syndrome and over expansion of CAR T cells. 131 This nanobody-based UniCAR showed controlled and substantial tumor cell lysis and good biodistribution with prolonged half-life. 130 A number of nanobodies in preclinical development are listed in Table 3.
Therapeutic Nanobodies in Preclinical Development
CEA, carcinoembryonic antigen; GFAP, glial fibrillary acidic protein; HGF, hepatocyte growth factor; MHC, major histocompatibility complex.
Nanobodies in Cancer Imaging
In addition to their use as therapeutic agents, nanobodies also make highly effective and noninvasive molecular imaging reagents due to their high penetration, quick elimination, low immunogenicity, and ease of production and selection. 132,133 Some examples of the applications of nanobodies in oncology/imaging include locating tumors and cancer cell metastasis and visualizing and monitoring lymphocytes and APC infiltration into solid tumors. 134,135 The nanobody 8B6 is a 99mTc radiolabeled nanobody against the EGFR, which is used in the radioimmunodetection of EGFR overexpressing tumors by single-photon emission computed tomography (SPECT). 136 This nanobody is very effective in the detection of human epidermoid carcinoma and human prostate carcinoma. Hepatocyte growth factor (HGF) is a prominent growth factor in promoting invasiveness and metastasis in several types of cancers. 137 1E2 and 6E10 are two nanobodies constructed against HGF, coupled with albumin (to improve half-life), and are radiolabeled with 89 Zr. 138 These are very effective in visualizing HGF+ tumors using PET when administered to nude mice (bearing U87 MG glioblastoma xenograft), with good tumor biodistribution and half-life. These nanobodies also showed tumor suppressive activity by significantly delaying tumor growth. 138 Similarly, 2Rs15d, 1E2, and CEA1 nanobodies have been labeled with 99mTc and 89 Zr to detect ovarian carcinoma, breast cancer, human glioblastoma, colon adenocarcinoma. 106,138,139 Lecocq et al. (2019) developed 99mTechnetium-labeled nanobody specific for LAG-3 (a immune checkpoint receptor), for noninvasive tumor imaging. 140
Despite having good homology with human antibodies, a few camelid nanobodies show immunogenic responses when administered into patients; Ilse Vaneycken et al. (2010), to overcome undesirable immunogenic responses, constructed a humanized nanobody for in vivo molecular imaging of cancer. They drafted the antigen-binding loops of a camelid nanobody developed against CEA (NbCEA5), onto the framework region of a humanized nanobody scaffold. 141 The NbCEA5 and humanized nanobody scaffolds used in this study were previously prepared in the same laboratory and tested for their efficacy by the same group. 114,142 The human grafted nanobody (humanized CEA5 graft) demonstrated excellent specificity for both soluble and membrane bound CEA; it also showed good and safe biodistribution with efficient tumor targeting and quick elimination through kidney, when administered to LS174T tumor bearing mice. 141 This work revealed the prospects of developing humanized nanobodies with no loss in specificity and binding affinity.
Having knowledge of a patient's tumor microenvironment is highly favorable when planning a treatment strategy. An important example of this is knowing the ratio of anti- and proinflammatory macrophages. To identify tumor-associated macrophages (TAM) in the tumor microenvironment, nanobodies specific to macrophage mannose receptor (MMR/CD206) were labeled with 99mTc. 143 The background disturbance by the accumulation of 99mTc labeled nanobodies at extratumoral locations was lowered by injecting unlabeled, bivalent anti-MMR nanobodies. This approach allowed the detection of tumors through hypoxia-associated TAMs. 143 Furthermore, this was improved by replacing 99mTc with 18 F, which reduced nanobody accumulation in the liver and kidney. 144 Similarly, nanobodies for monitoring tumor-infiltrating lymphocytes have been developed. An anti-CD8 nanobody conjugated with 89 Zr enabled excellent surveillance of CD8+ T cells and was also PEGylated to reduce accumulation in the kidneys with improved imaging. 145 Monitoring of tumor burden is essential to understand the impact of a treatment; Gainkam LO et al. (2011) developed a strategy to monitor the treatment response of erlotinib (tyrosine kinase receptor inhibitor), by developing a correlation between tumor burden and the uptake of anti-EGFR nanobody (99mTc-7C12) by tumor. 146 The tumor burden and uptake of 99mTc-7C12 by tumor were monitored in vivo by bioluminescence imaging and pinhole single-photon emission computed tomography, respectively, in A431-fluc mice xenograft model when treated with erlotinib; it was observed that the tumor uptake of 99mTc-7C12 decreased with decrease in tumor burden by the antitumor activity of erlotinib. This demonstrated a strong correlation between tumor burden and uptake of 99mTc-7C12 and formed a basis for monitoring the response of a treatment by measuring the uptake of tumor-specific nanobodies by the tumor. 146
Conclusions
Nanobodies with favorable and dynamic properties, compact size, thermal stability, penetrating capability, even across the BBB, high solubility, and low immunogenicity make them very promising therapeutic molecules. Ease of manipulation and low cost of production further adds to their promise as therapeutics. They are being used to treat a wide variety of diseases, particularly cancer, by activating effector cells and inhibiting immune suppression in several dynamic strategies. The bispecific nanobodies lacking the VL-VH domain showed impressive stability, with minimal aggregation. Many naked antagonistic nanobody constructs showed successful inactivation of tumorigenic pathways by blocking ion channels, growth factor receptors. Their diminutive size displayed remarkable tissue penetration and biodistribution of nanobodies and nanobody-drug conjugates; this opened doors to combat solid tumors with enhanced efficacy. Nanobodies induced good antitumor response when exploited in photothermal and photodynamic therapy. Advances in synthetic biology lead to the development of probiotic bacteria, which are capable of synthesizing therapeutic antitumor nanobodies in the tumor microenvironment, to avoid toxicity challenges with conventional checkpoint inhibitors. CAR T and CAR NK-cells with nanobodies as variable domains enabled the development of bispecific CAR T and CAR NK-cells with excellent antitumor activity and tumor specificity. Their potential has attracted the interest of many pharmaceutical companies, with nanobodies now as the primary products in many drug pipelines. The very recent approval of a CABLIVI nanobody by the FDA for treating thrombotic thrombocytopenic purpura further highlights their emergence in the market and they undoubtedly continue to revolutionize treatment options.
Footnotes
Authors' Contributions
All coauthors have reviewed and approved of the article before submission.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this work.