
Abstract
Background:
APOBEC1 complementation factor (A1CF) is a component of the apolipoprotein-B messenger RNA editing complex that participates in various cellular processes and acts as an oncogene in many cancers. In this study, it was aimed to investigate the roles of A1CF and its potential mechanism in endometrial cancer (EC).
Materials and Methods:
Gene expression prolife was downloaded from The Cancer Genome Atlas database. Then Kaplan–Meier and Cox regression analyses were conducted to assess the prognostic value of A1CF in EC. Cell Counting Kit-8, plate clone formation, and transwell assays were used to estimate the functions of A1CF on the proliferation, invasion, and migration of EC cell. The gene set enrichment analysis was used to analyze the pathway that is enriched by A1CF, whereas quantitative real-time polymerase chain reaction and Western blot analyses were utilized to detect the mRNA and protein expression involved.
Results:
It was detected that the upregulated A1CF was enriched in P53/P21 signaling pathway and tightly associated with patients' age, stage, and death. Besides, high A1CF expression led to a shorter overall survival of patients and predicted a poor prognosis in EC. The overexpression of A1CF promoted the proliferation, invasion, and migration of EC cells, whereas the depletion of A1CF suppressed these processes. Moreover, P21 and P53 were reduced whereas cyclin D1 and proliferating cell nuclear antigen were induced along with the increasing of A1CF. However, the effects of silencing A1CF on these protein expressions were on the contrary.
Conclusion:
A1CF was highly expressed and closely related to the prognosis and progression of EC through the regulation of P53/P21 signaling pathway, providing a possible new therapy target site for EC.
Introduction
Endometrial cancer (EC) is the most frequent cancer in the female reproductive tract with ∼63,230 new cases in 2018. 1,2 Although most patients are diagnosed in the early stages of EC, some show poor prognosis and a high risk of recurrence and metastasis. 3 Surgery is the major therapy method for EC supplemented with other adjuvant therapies, including radiotherapy, chemotherapy, and therapy with hormones or targeted therapy. 4 For the past few years, targeted molecular therapies for controlling endometrial malignancies have aroused wide interest of many scholars, 5 but the studies are still deficient. 6 Hence, it is necessary to identify more molecule markers and clarify their mechanisms that are involved in the development and progression of EC.
APOBEC1 complementation factor (A1CF), also called ACF, is a kind of RNA binding protein, which belongs to the heterogeneous nuclear ribonucleoproteins (hnRNP) family. 7,8 There were three nonidentical RNA recognition motifs in A1CF that was initially described as an essential component for apolipoprotein B (ApoB) mRNA editing. 9 A1CF is once thought to be a docking site to recruit the APOBEC1 deaminase so that it regulates site-specific C to U edition of apoB mRNA. 10 Despite A1CF has been reported to play important roles in the malignant biological behaviors of several cancers such as glioma, 11 renal cell carcinoma, 12 and breast carcinoma, 13 its role in EC has never been revealed.
This study was conducted to investigate the functions of A1CF and its molecular mechanism in EC. The expression levels of A1CF in tissues and cells of EC were analyzed using bioinformatic analysis and quantitative real-time polymerase chain reaction (qRT-PCR) analysis, respectively. Associated with this, the prognostic value of A1CF was estimated, meanwhile the effects of A1CF on the proliferation, invasion, and migration of EC cells were detected after A1CF was overexpressed or silenced. Furthermore, the expression levels of several key proteins in P53/P21 pathways were checked to explore the underlying mechanism of A1CF in the tumor progression of EC.
Materials and Methods
Data collection
Research methods in this study did not include any clinical trials. The Cancer Genome Atlas (TCGA,
Cell culture
Human EC cell lines (RL95-2, HEC-1-A, and KLE) and normal uterine endometrial epithelial cells were provided by
Cell transfection
Si-A1CF#1 (5′-GCAACAATAGAGGATATGCA-3′), si-A1CF#2 (5′-GCTACTGC-TTTCCCAGGATA-3′), and si-con (5′-CGAACUCACUGGUCUGACC-3′), pcDNA3.1-A1CF or vector control pcDNA3.1 was synthesized by Shanghai GenePharma Co., Ltd. Cell transfection was carried out using Lipofectamine 2000 (Invitrogen) according to manufacturer's protocols.
Cell treatment
For P53 inhibition, after transfection, RL95-2 cells were treated with 10 μM pifithrin (PFT-α) for 48 h to inhibit p53 expression.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from EC cell lines and normal uterine endometrial epithelial cells using TRIzol reagent (Invitrogen) in accordance with the specification. Then the reverse transcription was performed to synthesize the first strand of cDNA. Next, a qRT-PCR was carried out in a Step One Plus Real-Time PCR System (Applied Biosystems, Life Technologies) to detect the relative mRNA expression of A1CF, which was calculated by the 2–△△Ct method. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was taken as the internal control. Each experiment was repeated three times. The sequences of relevant primers are listed as follows:
A1CF: F-5′-CAGCCACCAAAGGACATCTCAG-3′;
R-5′-CTGTGTATGCCAAATAGCCACGG-3′;
GAPDH: F-5′-GGAGCGAGATCCCTCCAAAAT-3′;
R-5′-GCCATCACGCCACAGTTTC-3′.
Western blot assay
Total protein was isolated using RIPA lysis buffer (with protease inhibitor) and measured by bicinchoninic acid (BCA) protein assay kit (Sigma). After being heated for 5 min, 20 μg proteins was added into each well of vertical electrophoresis tank and separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis. Subsequently, the separated protein was transferred onto the polyvinylidene difluoride membranes that were then blocked by 5% nonfat milk. The membranes were incubated with the primary antibodies: anti-A1CF (ab89050; Abcam), anti-P21 (ab109199; Abcam), anti-P53 (ab26; Abcam), anti-cyclin D1 (ab16663; Abcam), and antiproliferating cell nuclear antigen (PCNA) (Proteintech Group, Inc.) overnight at 4°C. After that, the membranes were washed by tris-buffered saline tween-20 (three times and each time for 5 min) and mingled with the secondary antibody for 1 h at indoor temperature. Finally, the membranes were washed and developed by enhanced chemiluminescence. The gray value was scanned by QUANTITY ONE software and the relative protein expression was calculated. GAPDH was taken as the internal control.
Assessment of cell viability
Cell suspension solution was prepared and 100 μL of it was planted into 96-well plates (1000 cells/well) and cultured in a CO2 incubator. Cell viability was checked every 24 h, before which 10 μL CCK8 regent was put into each well and cultured at 37°C for 1.5 h. Then the OD450 value was tested using a microplate reader.
Transwell assay
The invasive and migratory abilities of cells were analyzed by transwell assays. For invasion assay, 100 μL matrigel was added into the upper chambers of the 24-well plates that stood for 4–6 h at 37°C in a CO2 incubator after being shaken up. Then 500 μL serum-free medium was put into the lower chamber that stood for 0.5 h. Subsequently, 100 μL (1 × 105 cells) cell suspension solution was put into the upper chamber, whereas 500 μL complete medium was added into the lower chamber. The chambers were taken out and the remanent cells in the upper chambers were erased and fixed by 4% paraformaldehyde for 30 min. Next, 0.1% crystal violent was used to stain the cells for 20 min. Finally, the number of cells was counted in five views that were chosen randomly after the cells were washed by phosphate-buffered saline (PBS). For cell migration, the steps were similar to the invasion assay except for adding matrigel.
Plate clone formation assay
Cells in logarithmic phase were digested by trypsase and blown into individual cells, which were made into cell suspension solution and counted. Then the cells were planted into 60 mm petri dishes (400 cells in each dish) containing 5 mL of 37°C prewarmed nutrient solution and cultured in an incubator at 37°C with an atmosphere of 5% CO2 for 1–2 weeks after the cells were homogeneously dispersed until the colony occurred. Next, the cells were rinsed by PBS for two times after the liquid supernatant was discarded and fixed by 5 mL of 4% paraformaldehyde for 30 min, after that 0.1% crystal violent was utilized to dye the cells for 30 min. After the staining solution was rinsed and the cells were dried, the number of colonies was counted.
Statistical analysis
SPSS22.0 software was used to analyze the data involved that are presented as mean ± standard deviation. χ 2 test was utilized to analyze the relationship between A1CF and the clinicopathologic characteristics of patients with EC. The differences between two groups were compared by Students' t test, whereas those among three groups or above were analyzed by one way analysis of variance. The difference was considered to be significant when p < 0.05.
Results
A1CF is highly expressed in the tissues and cells and associated with the prognosis of patients with EC
To explore the expression of A1CF in EC, the data from TCGA database were used. The result showed that A1CF expression was significantly higher in the tissues of patients with EC than in normal tissues (Fig. 1A, p < 0.05). qRT-PCR analysis also revealed that A1CF was highly expressed in EC cells compared with normal uterine endometrial epithelial cells (Fig. 1B, p < 0.05). Besides, GSEA suggested that A1CF was enriched in the P53/P21 signaling pathway (Fig. 1C).
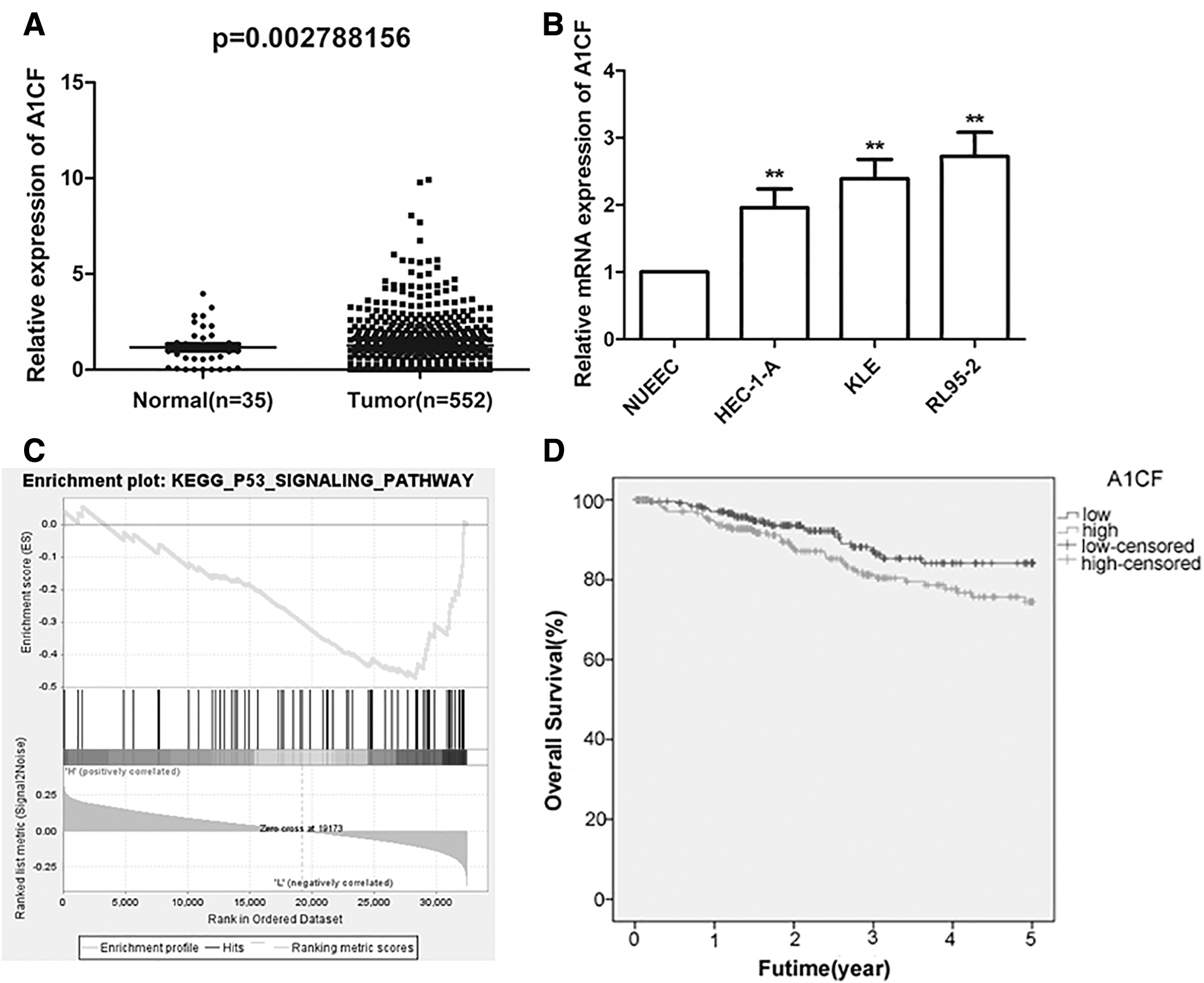
The identification of A1CF in EC.
Next, the prognostic value of A1CF was assessed. First, based on the median of A1CF expression, EC patients were equally divided into two groups: the high A1CF expression group (n = 257) and the low A1CF expression group (n = 257). Then the clinicopathologic characteristics of patients with EC were collected and their relationship with A1CF expression was analyzed. The outcomes displayed that the expression of A1CF was obviously associated with the age, stage, and death of the patients (Table 1). Subsequently, Kaplan–Meier analysis was utilized to compare the overall survival of patients with high A1CF expression and low A1CF expression, which indicated that the high expression of A1CF led to a shorter overall survival time compared with low A1CF expression (Fig. 1D, log rank test: p < 0.05). Moreover, univariate Cox regression analysis proved that A1CF expression was closely related to the prognosis of patients, whereas multivariate Cox regression analysis revealed that A1CF could not be an independent prognostic marker in EC (Table 2). These results demonstrated that A1CF was overexpressed and closely related to the age, stage, and death of patients and patients' overall survival, but it could not be an independent prognostic factor in EC.
The Association Between APOBEC1 Complementation Factor Expression and the Clinical Features of Endometrial Cancer Patients from The Cancer Genome Atlas
p < 0.05.
A1CF, APOBEC1 complementation factor.
The Independent Prognostic Factors Were Analyzed by Univariate and Multivariate Cox Regression Analysis
p < 0.05.
Transfection efficiency detection
To investigate the transfection efficiency, the mRNA and protein expression levels of A1CF were checked in RL-95-2 transfected with si-A1CF#1/si-A1CF#2/si-con and HEC-1-A cells transfected with pcDNA3.1-A1CF or vector control pcDNA3.1. qRT-PCR analysis exhibited that the relative mRNA expression of A1CF was markedly downregulated in RL-95-2 cells but upregulated in HEC-1-A cells after the transfection assay, whereas the protein expression of A1CF showed the same trend through Western blot analysis (Fig. 2A–D, p < 0.05).

Expression levels of A1CF in cells transfected with si-A1CF/si-con or pcDNA3.1-A1CF/pcDNA3.1.
A1CF promotes EC cell proliferation
To estimate the influences of A1CF on the proliferative ability of EC cells, the cell viability of RL-95-2 and HEC-1-A was checked. As a consequence, the cell proliferation of RL-95-2 cells was significantly inhibited by the silencing A1CF, whereas overexpression of A1CF obviously accelerated the proliferation of HEC-1-A cells (Fig. 3A, B, p < 0.05). Besides, plate clone formation assay demonstrated that the colonies formed by RL-95-2 cells were notably less in the si-A1CF group than those transfected with si-con (Fig. 3C), meanwhile the colonies were dramatically raised by the overexpression of A1CF in HEC-1-A cells (Fig. 3D). Above all, A1CF was positive to the EC cell proliferation.
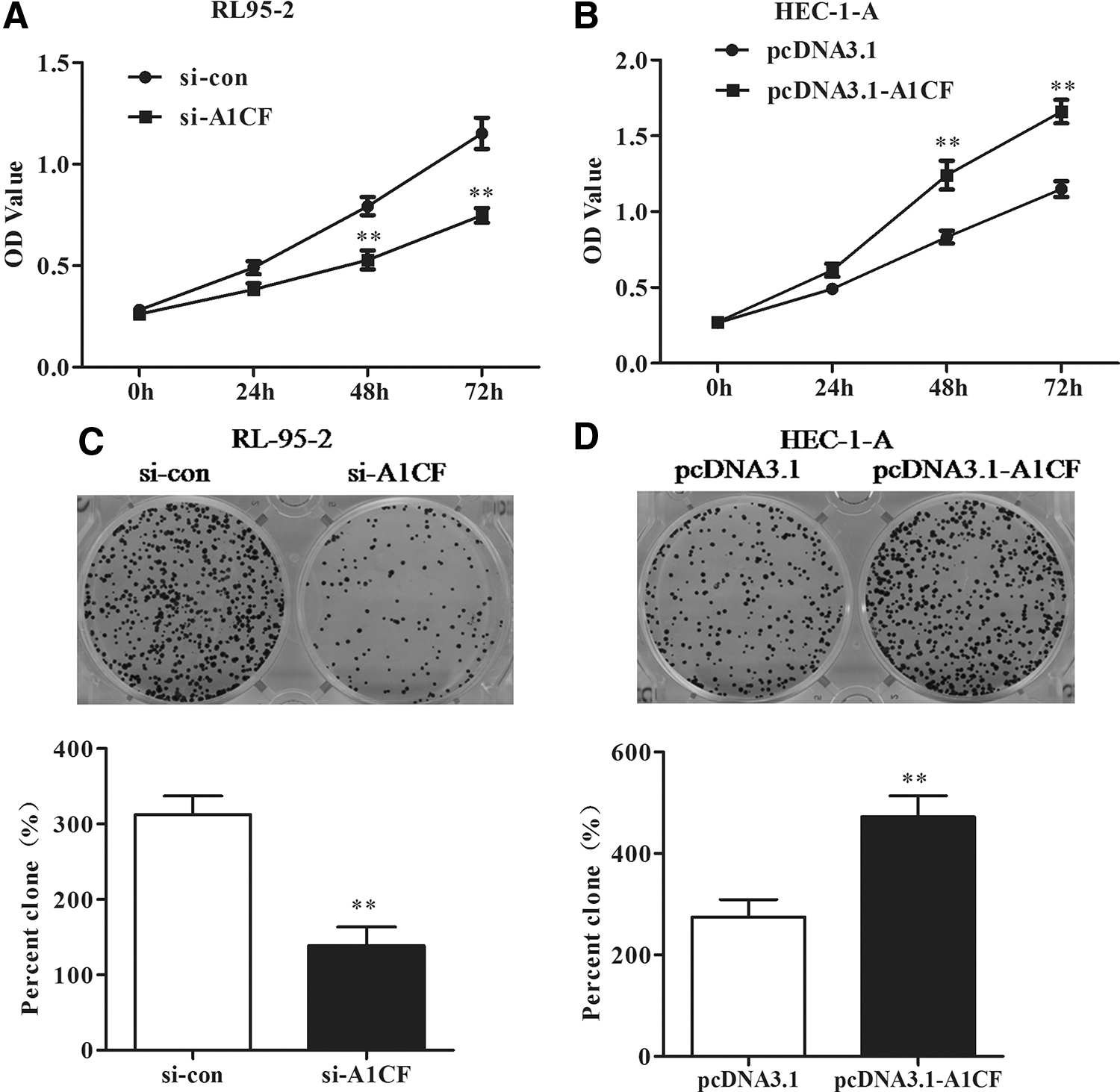
A1CF regulated the cell proliferation and colony formation.
A1CF facilitated the invasion and migration of EC cells
Transwell assay was used to evaluate the invasive and migratory capabilities of EC cells. As shown in Figure 4, the invasion and migration of EC cells were observably restrained by the knockdown of A1CF in RL-95-2 cells (Fig. 4A, p < 0.05), whereas those were expedited by the upregulation of A1CF in HEC-1-A cells (Fig. 4B, p < 0.05). Therefore, A1CF might be a positive factor for the invasion and migration of EC cells.

The influences of A1CF on the invasion and migration of EC cells.
A1CF inactivates the P53/P21 signaling pathway
To further research the underlying mechanism of A1CF in EC, its enriched pathways were screened and P53/P21 signaling pathway was obtained according to GSEA. Then to check on whether A1CF impacted the P53/P21 signaling pathway, the relative expression of relevant proteins was tested using Western blot analysis. The data showed that the expression levels of P21 and P53 were significantly elevated, whereas the expression levels of cyclin D1 and PCNA were observably decreased by silencing A1CF in RL-95-2 cells (Fig. 5A, p < 0.05). On the contrary, P21 and P53 were reduced whereas cyclin D1 and PCNA were induced by the upregulation of A1CF in HEC-1-A cells (Fig. 5B, p < 0.05). Therefore, A1CF possibly exerted its functions in EC through the inactivation of the P53/P21 signaling pathway.

The expression of P21, P53, cyclin D1, and PCNA in EC cells.
A1CF promoted the biological behaviors through suppressing the P53/P21 signaling pathway
To confirm the mentioned notion, the expression levels of P53 in the si-NC and si-A1CF transfected RL95-2 cells were inhibited with 10 μM pifithrin-α (PFTα) and subsequently the biological behaviors were determined by CCK-8 and transwell assays. As shown in Figure 6A, after transfection for 48 h, the OD values were significantly higher upon PFTα treatment compared with the si-A1CF group (p < 0.05). Likewise, PFTα treatment gave rise to the capabilities of invasion and migration in RL95-2 cells transfected with A1CF siRNA (Fig. 6B, C, p < 0.05). Overall, inhibition of P53 attenuated the impairments of A1CF knockdown to biological behaviors of EC cells, suggesting that it was through suppressing the P53/P21 signaling pathway that A1CF promoted EC cell biological behaviors.
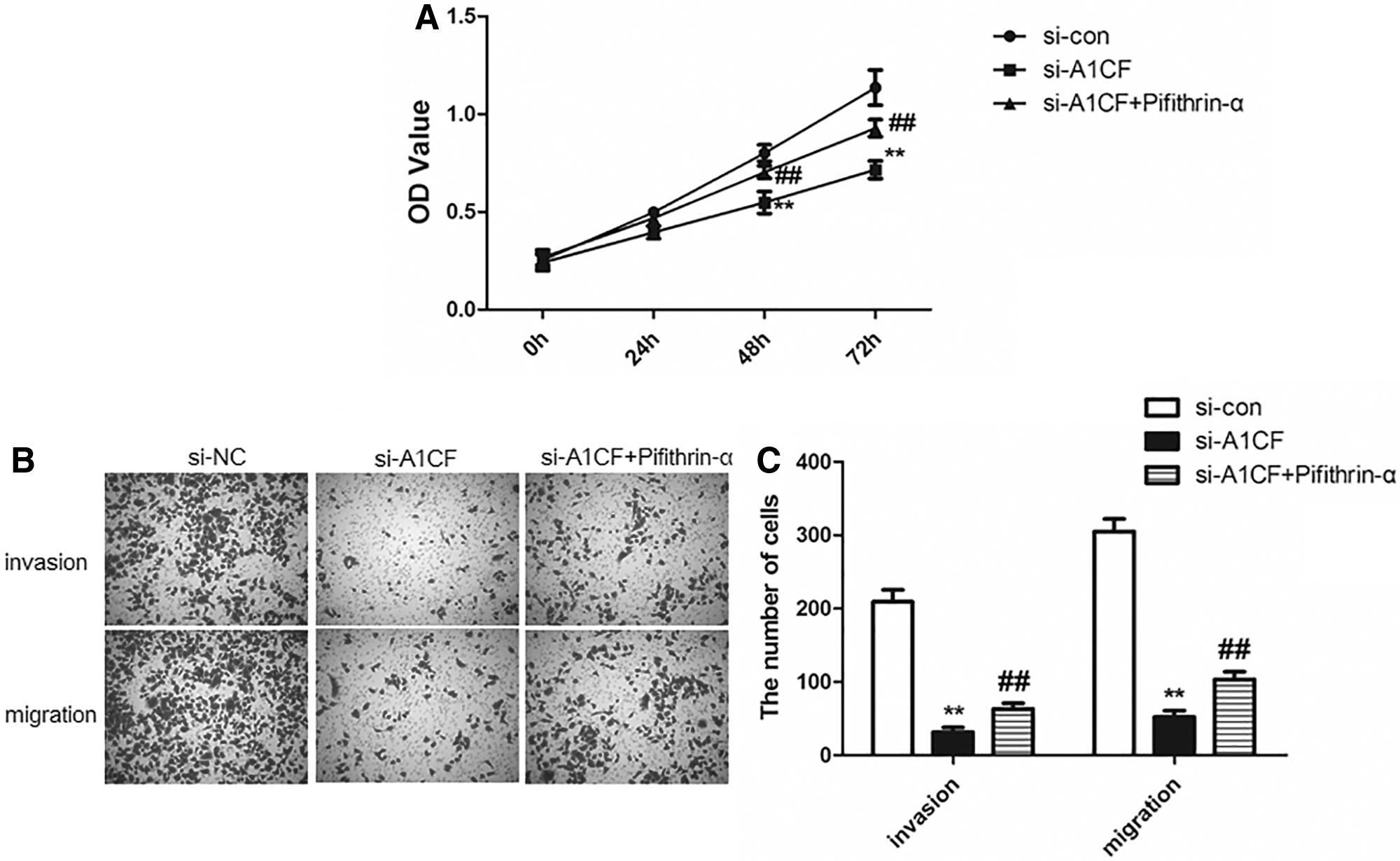
The influence of P53 inhibitor pifithrin-α on the proliferation, invasion and migration of EC cells transfected with A1CF siRNA.
Discussion
In this study, it was revealed that A1CF was upregulated in EC tissues and cancer cell lines, which was associated with a poor prognosis. In addition, loss of function and gain of function of A1CF implicated that A1CF had a positive effect on the biological behaviors of EC cells. In line with these results, A1CF was previously reported to be highly expressed in several cancers, including glioma, 11 Wilms tumor, renal cell carcinoma, 12 and breast cancer. 14,15 In addition, the upregulation of A1CF was associated with a worse survival outcome of EC. Consistently, high A1CF expression level indicated a poor prognosis of glioma and breast cancer. 11,15 Furthermore, numerous lines of evidence implicated that the upregulation of A1CF supports the malignant progression of some cancers. For instance, overexpression of A1CF modulated the cell apoptosis and migration of Wilms tumor-derived cells through the Wnt/β-catenin pathway. 16 The downregulation of A1CF restrained the proliferation and colony formation of renal cell carcinoma cells depending on DKK1-MEK/ERK signal axis. 17 In this study, the proliferation, migration, and invasion were inhibited after the depletion of A1CF, whereas overexpression of A1CF promoted these cellular processes. These results suggested that A1CF had an oncogenic effect on EC cancer cells.
Moreover, the mechanism of how A1CF regulated cellular processes in EC was further investigated. This study exhibited that A1CF promoted the biological behaviors partly through inactivating P53/P21 signaling pathway. The P53/P21 pathway was involved in the tumor progression of EC according to previous studies. 18,19 P53, an important tumor suppressor, is dysregulated in numerous cancers. 20 –22 P21 is a vital member of the Cip family and the cyclin-dependent kinase inhibitor family, 23,24 and is regulated by P53 in response to DNA damage. 25,26 Cyclin D1 is crucial to cell cycle regulation and can exert oncogenic effects in numerous human cancers, including EC. 27 –29 PCNA, a nuclear protein, acts as a marker of cell proliferation. 30 Moreover, PCNA was upregulated in EC. 31 Besides, the expression of P53 and P21 was considered to mediate the cell cycle by regulating the expression of cyclin D1 and PCNA. 32 Consistently, loss-of-function of A1CF induced the downregulation of cyclin D1 and PCNA and upregulation of P21 and P53, suggesting that P53/P21 signaling pathway was activated after A1CF knockdown. The findings provided evidence that P53/P21 signaling pathway engaged in the A1CF regulation of malignant development of EC. However, the impairments to biological behaviors of EC cells after depletion of A1CF were partly mitigated by the inhibition of P53, suggesting that P53/P21 signaling pathway was not the only target of A1CF. There are many reports that A1CF modulated the malignancy of cancers through several pathways or proteins such as Wnt/β-catenin pathway, 16 long noncoding RNA, 11 and Dickkopf1. 14 Therefore, the targets of A1CF in EC await to be further identified.
Taken together, A1CF, an overexpressed gene, predicted a worse survival of patients and participated in the tumor progression of EC by mediating P53/P21 signaling pathway. This is the first study to research the role of A1CF in EC, which may provide a new therapy target site for EC. Besides, in vivo experiments for validating the function of A1CF await to be conducted and whether A1CF has other functions and mechanisms are also needed to be explored in the future.
Availability of Data and Material
The data and material in this study are available from the corresponding author on reasonable request.
Authors' Contributions
Q.L. and G.-L.C. designed this study. Q.L., C.-Y.C., and G.-L.C. carried out the study and analyzed the data. Q.L. wrote the article and G.-L.C. revised it. All the authors approved the final version of the article.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.