
Abstract
Background:
Recently, several clinical studies have reported that combination treatments of radiation therapy (RT) and immunotherapy in patients with multiple lesions can improve tumor regression at a distance from the irradiated site, known as the abscopal effect. However, when RT and immunotherapy are concurrently applied, it is hard to distinguish the pure systemic effects of RT from those of the immunotherapy drug. In this preclinical study, the authors investigated the systemic antitumor effects of RT alone according to fraction dose size and splitting schedules.
Materials and Methods:
4T1 mouse breast cancer cells were implanted into the right and left sides of mammary gland fat pads of BALB/c mice, followed by irradiation with 6 Gy × 3, 8 Gy × 2, and 13 Gy × 1 fractions when the right-side tumors were palpable.
Results:
The different irradiation schedules produced similar antitumor effects in irradiated right-side tumors and unirradiated left-side tumors. However, 8 Gy × 2 and 13 Gy × 1 fractions exhibited better antimetastatic potential than that from irradiation using 6 Gy × 3 fractions. Furthermore, 8 Gy × 2 and 13 Gy × 1 fractions produced higher expressions of HMGB1 and lower expressions of the proinflammatory cytokines, IFN-γ, TNF-α, IL-6, and IL-1β, from the irradiated tumor tissues.
Conclusions:
These findings suggest that 8 Gy × 2 and 13 Gy × 1 fractions can provide better systemic antitumor effects than 6 Gy × 3 fractions. The authors hope these results provide clues to optimize RT dose regimens to make the abscopal effect clinically more relevant in future combination treatments.
Introduction
Radiation therapy (RT) is considered a local treatment because radiation is directly delivered to a local disease, leading to DNA damage followed by cellular death. The ability of RT to control a local disease has been well established and there are profound radiobiological studies elucidating its underlying mechanisms. 1,2
In addition to the local tumor-controlling ability of RT, systemic effects of RT have been infrequently reported over the past 50 years. 3 –5 Several clinical studies described patients with multiple lesions exhibiting tumor regression at a distance from the irradiated site. Mole first defined this tumor regression effect at an out-field of radiation as an “abscopal effect.” 3 Although the mechanism producing abscopal effects is still ambiguous, it is believed that abscopal effects are mediated by activation of the immune system. Accumulating evidence suggested that radiation induces immunogenic cell death in a tumor and stimulates the release of a number of danger-associated molecular patterns (DAMPs). The release of DAMPs triggers maturation of dendritic cells (DCs), subsequently enhancing tumor antigen presentation and production of several cytokines to promote immune responses and destroy tumors. 6 –9
However, recognizing a true abscopal effect of RT is very difficult, particularly in a clinical setting because its overall occurrence rate is relatively low and RT is generally used as an ancillary treatment to chemotherapy in patients who present multiple lesions or systemic progression. Nevertheless, systemic effects of RT arising through immune responses have drawn scientific interest, especially in conjunction with the development of immunotherapy in cancer treatment. Immunotherapy is a treatment intended to boost a patient's immune system and destroy cancer. Nowadays, it is evolving markedly in the treatment of several types of cancer, and related clinical studies are underway in many countries. 10 In this regard, the combination of immunotherapy and RT has received attention in the field of radiotherapy research.
Traditionally, a low-dose and high-fractionated irradiation schedule (conventional radiation schedule) has been used as a standard therapy aimed at minimizing damage to normal tissues. However, recent developments in radiation delivery techniques, such as intensity-modulated RT, image-guided RT, and proton therapy, allow the use of hypofractionated irradiation schedules in cancer treatment. 11 –13 Clinical applications of hypofractionated irradiation schedules have rapidly increased, but existing radiobiological studies are not sufficient to elucidate the underlying mechanisms. Interestingly, some clinical studies have shown that hypofractionated irradiation schedules may induce greater immune responses than those from a conventional radiation schedule when RT is combined with immunotherapy. 14 –20
A limited number of clinical trials are currently investigating the combined effects of immunotherapy and RT in several types of cancers. However, there is concern that some clinical trials have proceeded in the absence of an established consensus on the optimal RT fraction, dose size, and splitting schedule needed to induce an abscopal response. Thus, before clinical studies, sufficient preclinical studies should be conducted to determine which RT fraction, dose size, and splitting schedule would improve the systemic antitumor effects of RT.
In this study, the authors investigated the RT fraction dose sizes and splitting schedules needed to induce systemic RT effects in a preclinical model. To evaluate the systemic effects of RT, a preclinical model should present multiple tumor lesions. For that purpose, they utilized mice bearing 4T1 mouse breast cancer cells because 4T1 cells grow rapidly to form a palpable tumor and metastasize to the lung within 2 weeks. To evaluate the systemic effects of RT, they irradiated mice bearing 4T1 tumors with different fractionated doses and schedules and analyzed the release of DAMP and tumor regression in multiple lesions.
Materials and Methods
Cell culture
4T1 mouse breast cancer cells were purchased from American Type Culture Collection (Rockville, MD) and subcultured in Dulbecco's modified Eagle's medium (Welgene, Daegu, Korea) supplemented with 5% fetal bovine serum (Invitrogen, Carlsbad, CA) and 1% antibiotic/antimycotic solution (Welgene).
Irradiation
4T1 cells and mice bearing 4T1 breast tumors were irradiated using a 21EX Linac (Varian Medical Systems, Palo Alto, CA). Dose distribution was confirmed using the Eclipse planning system (Varian Medical Systems). To irradiate cells in plates, the plate was placed on a 2 cm solid phantom, and then irradiated by 6 MV X-rays at a gantry angle of 180 degrees. The irradiated field size was 20 × 20 cm, and the surface distance from the source was 98 cm. To irradiate tumors in right-side fat pads of mice, the gantry angle was adjusted according to the location of the tumor, and a 6 MeV electron beam with a 5 mm bolus was used. The source to surface distance was 100 cm and the irradiation area was 1.5 cm in diameter (Fig. 2B).
Colony forming assay
4T1 cells were seeded into a six-well plate at a density of 200–2000 cells/well and irradiated with 6 Gy × 3, 8 Gy × 2, or 13 Gy × 1 fractions. Cells were further incubated for 7 d. The formed colonies were fixed with 10% formalin and stained with 0.01% crystal violet. Colonies of ≥50 cells were counted manually under a microscope (TS 100, Nikon, Japan). Survival fractions of the irradiated cells with the three fractions were normalized by plating efficiencies of unirradiated cells and calculated by comparing the number of colony of unirradiated cells.
Cell viability assay
4T1 cells were seeded in a 96-well plate at a concentration of 700 cells/well and irradiated with 6 Gy × 3, 8 Gy × 2, or 13 Gy × 1 fractions (Fig. 1A). After culturing irradiated cells for 24, 48, and 72 h, MTT reagent was added to each well and the formazan formed by the cells was dissolved by DMSO. The absorbance of the dissolved formazan was measured at 570 nm using a multidetection microplate reader (Molecular Devices, CA).
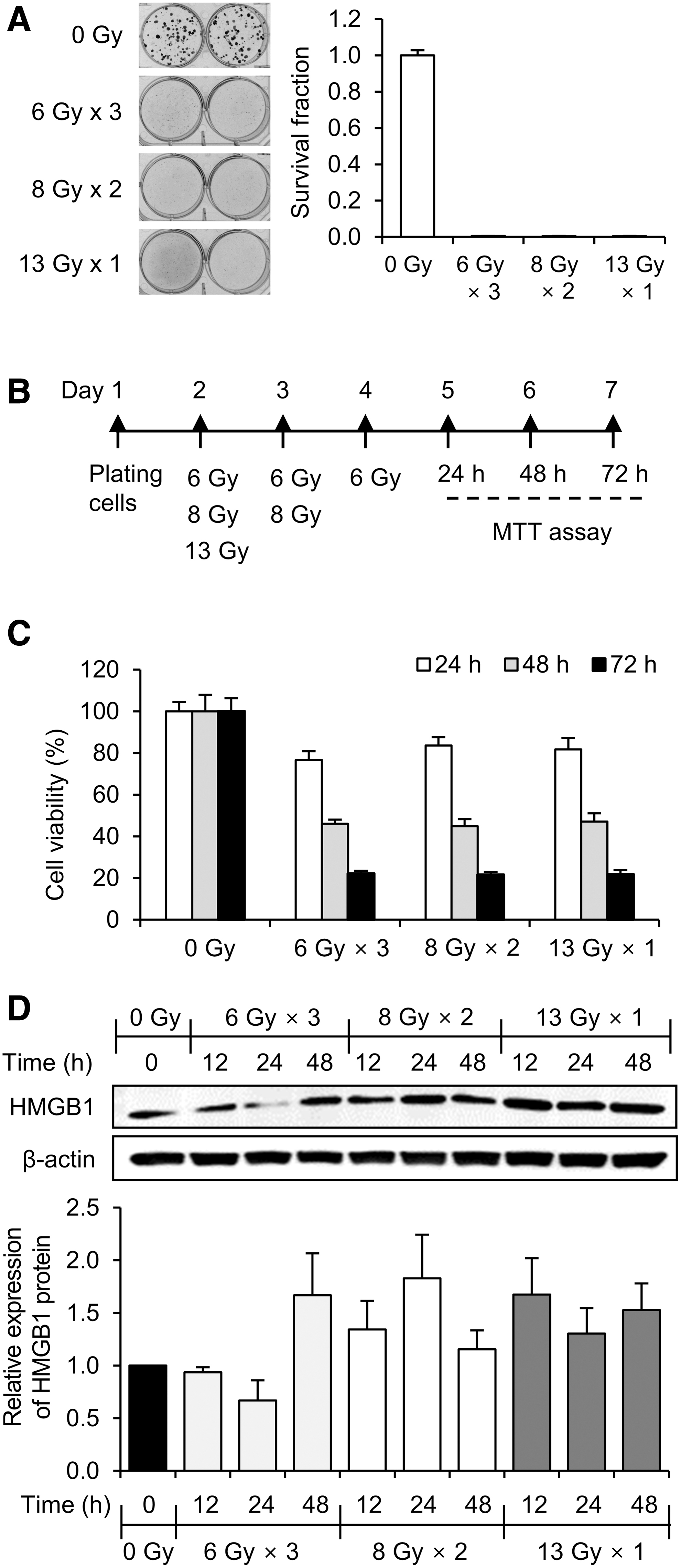
Effect of three different irradiation schedules on in vitro 4T1 mouse breast cancer cells.
Tumor formation and irradiation
Animal experimentation was performed in the Laboratory Animal Research Center after obtaining approval from the Institutional Animal Care and Use Committee at Dongguk University (IACUC-2016-016). Four-week-old female BALB/c mice were purchased from Orient Bio, Inc. (Seongnam, Korea). The mice were maintained under a 12-h light/dark cycle at 25°C ± 2°C and a relative humidity of 50% ± 5% for 2 weeks before 4T1 cell inoculation.
Initially, 4T1 cells (1 × 105) were suspended in 20 μL of serum-free media and directly injected into #4 right-side mammary fat pads (irradiated region) of mice. 21 One week later, 4T1 cells (1 × 105) were prepared by the same method and implanted into left-side mammary fat pads of the same mice (unirradiated region). At day 13 after right-side inoculation, right-side tumors were palpable and the mice were grouped into three treatment groups and one control group (n = 4 per group). Then, the mice were started by irradiating right-side tumors with 6 Gy × 3 fractions, 8 Gy × two fractions, or 13 Gy × one fraction. No more than one fraction was delivered per day. Before irradiation, the mice were anesthetized with sodium pentobarbital (50 mg/kg) by intraperitoneal injection (Fig. 2A). The mice were checked every other day to monitor weight variation and tumor size. The horizontal and vertical lengths of each tumor were measured by using calipers. To reduce measurement error, one researcher repeatedly measured each individual three times and calculated the average value.
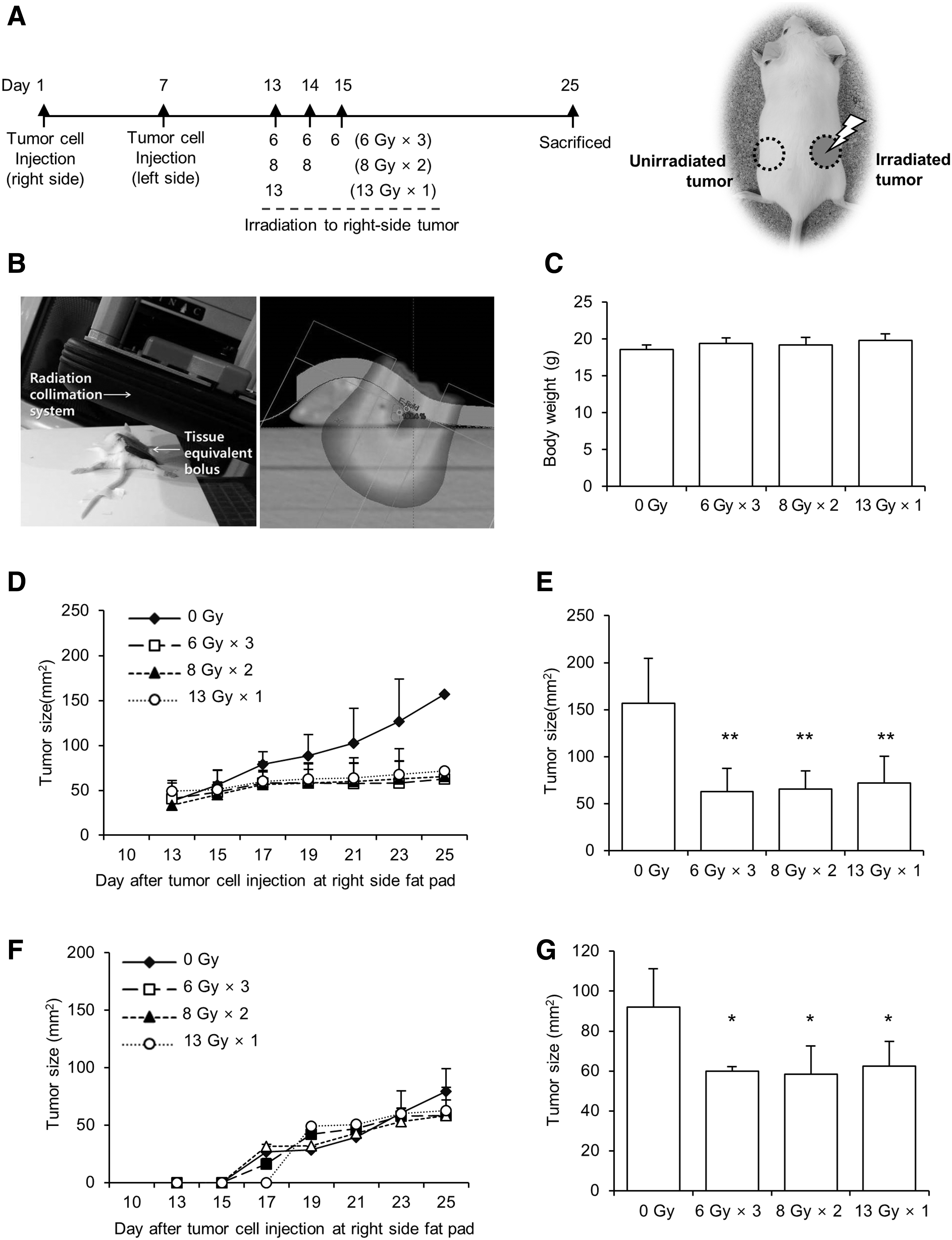
Three different irradiation schedules exhibited similar antitumor effects in irradiated right-side tumors as well as in unirradiated left-side tumors.
Lung metastasis analysis
After harvesting lungs from mice, lungs were fixed with 10% neutral buffered formalin and embedded in paraffin. Lung-embedded paraffin blocks were sectioned at a 4 μm thickness and stained with H&E after deparaffinization. Observation of a definite cluster of more than 15 tumor cells was considered indicating the presence of a metastatic lesion. The total lung area and the tumor metastasis area were determined using an image program (Olympus cellSens™ Microscope Imaging Software). The metastatic tumor proportion was the ratio of the metastatic area to the total lung area and is expressed as a percentage. All procedures were performed by a clinical pathologist.
Western blot analysis
To analyze the expression of HMGB1 in 4T1 cells irradiated with 6 Gy × 3, 8 Gy × 2, or 13 Gy × 1 fractions, 3 × 105 cells/mL were seeded into six-well plates and radiation applied as indicated. Irradiated cells were cultured for a further 24 h and then lysed by RIPA lysis buffer (150 mM NaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS, 50 mM Tris-HCl, pH 7.5, and 2 mM EDTA) containing phosphatase inhibitor and protease inhibitor cocktails (GenDEPOT, Barker, TX). The lysates were centrifuged at 13,000 rpm/min for 10 min to remove cell debris. To prepare lysates from irradiated tumors, tumor masses were homogenized with RIPA lysis buffer containing phosphatase and protease inhibitor cocktails, and the tumor lysates centrifuged at 13,000 rpm/min for 10 min to separate debris.
Protein concentrations were determined using bicinchoninic acid reagent (Sigma) and measured amounts of protein were separated by SDS-PAGE and then transferred to polyvinylidene fluoride membranes. After blocking the membrane with 5% skim milk for 2 h, membrane blots were incubated with HMGB1 and β-actin (Cell Signaling Technology, Beverly, MA) antibodies at 4°C overnight. Blots were incubated with horseradish peroxidase-conjugated secondary anti-rabbit antibody (1:3000; Santa Cruz Biotechnology, Inc., Santa Cruz, CA) at room temperature for 1 h, and developed by using an LAS-4000 (Fujifilm, Tokyo, Japan) luminescent image analyzer.
RNA isolation and real-time PCR
Total RNA was extracted using the easy-BLUE™ Total RNA Extraction kit (iNtRON Biotechnology, Inc., Seongnam, Korea). The cDNA was synthesized using GoScript™ reverse transcriptase (Promega Corporation, Madison, WI) and PCR was performed using Taq DNA polymerase (Thermo Scientific, Rockford, IL). Real-time PCR was performed using the Eco™ Real-Time PCR System (Illumina, CA). The primer sequences used for real-time PCR are presented in Table 1.
Sequences of Primers and Annealing Temperatures Used for Real-Time Polymerase Chain Reaction
Statistical analysis
Statistical analysis was performed using SPSS 18.0 (SPSS, Inc., Chicago, IL). The experimental results were analyzed by performing one-way ANOVA with LSD applied as the post hoc test. Data are expressed as mean ± standard deviation values. Results were considered to be statistically significant when the p-value was <0.05
Results
Three different isoeffective dose fractionation schemes do not produce similar effects on HMGB1 release
High irradiation dose can cause death in mice due to radiation enteritis. Thus, after considering the results in several published articles, 22 –24 the authors selected 13 Gy as the highest single-fraction dose for mouse irradiation. Next, they applied a biologically effective dose (BED) formula to determine three radiation splitting schedules: 6 Gy × 3, 8 Gy × 2, and 13 Gy × 1 fractions. 25,26 Since BED is based on a linear/quadratic cell survival relationship in radiobiology, the 6 Gy × 3, 8 Gy × 2, and 13 Gy × 1 fractions are theoretically isoeffective doses. However, BED tends to be inaccurate as fraction dose size increases. 25,26 Thus, the authors tested whether the three selected radiation splitting schedules produced similar biological effects under real experimental conditions. First, they performed a clonogenic cell survival assay after irradiating 4T1 mouse breast cancer cells with 6 Gy × 3, 8 Gy × 2, or 13 Gy × 1 fractions, but the survival fractions of all three irradiated cells were too low to show that the 6 Gy × 3, 8 Gy × 2, and 13 Gy × 1 fractions are isoeffective doses (Fig. 1A). It might be due to that the irradiation dose was too high to form colonies. Thus, the authors next measured cell viabilities by using MTT assays after irradiating cells with 6 Gy × 3, 8 Gy × 2, or 13 Gy × 1 fractions (Fig. 1B). As shown in Figure 1C, there was no statistically significant difference among the cells irradiated with the three different fractionations, confirming that the 6 Gy × 3, 8 Gy × 2, and 13 Gy × 1 fractions are isoeffective doses.
Next, they investigated whether different radiation splitting schedules affected the release of DAMPs. Since HMGB1 is one of the major DAMPs, the expression of HMGB1 was analyzed in cells irradiated with 6 Gy × 3, 8 Gy × 2, or 13 Gy × 1 fractions. Interestingly, the more the radiation was fractionated, the less the HMGB1 expression was induced. As shown in Figure 1D, the expression of HMGB1 was relatively low when cells were irradiated with three fractions of 6 Gy. In contrast, HMGB1 expression was highly induced by a single dose of 13 Gy and that increase persisted for up to 48 h (Fig. 1C), implying that the expression of DAMPs may be affected by fraction dose sizes and splitting schemes.
Local RT effects in mice bearing 4T1 cells appear to be similar for three different fractionation schemes
4T1 mouse breast cancer cells mimic closely the behavior of aggressive, metastatic breast cancer. When 4T1 cells are implanted into mouse mammary gland fat pads, cells grow fast enough to form a palpable tumor within 10 d and to metastasize primarily into lungs. Thus, the authors chose 4T1 cells to investigate whether different fraction dose sizes and schemes can affect local and systemic antitumor effects of RT.
4T1 cells were implanted into the right- and left-side mammary gland fat pads of BALB/c female mice at days 1 and 7, respectively (Fig. 2A). When tumors on the right side were palpable at day 13, they started irradiating the right-side tumors with 6 Gy for 3 d, 8 Gy for 2 d, or 13 Gy for 1 d (Fig. 2A). The dose distribution was confirmed by the Eclipse planning system results (Fig. 2B). Mouse body weights were unaffected by these irradiation schedules, suggesting that radiation did not produce harmful effects in normal tissues (Fig. 2C).
The local tumor-controlling abilities of radiation were evaluated by measuring the tumor mass in the right-side fat pad (irradiated region), whereas the systemic antitumor effects of radiation were evaluated by measuring the tumor mass in the left-side fat pad (unirradiated region) and the amount of lung metastasis.
The results showed that right-side tumor growth was efficiently suppressed by all three radiation splitting schedules. As shown in Figure 2D and E, tumor size was significantly decreased in all irradiated groups compared with the control group. However, there was no difference among right-side tumor sizes in mice irradiated with 6 Gy × 3, 8 Gy × 2, and 13 Gy × 1 fractions, suggesting that the local effect of RT was unaffected by the different fractionation schemes.
Tumor growth in a radiation out-field area was delayed by three different fractionation schemes
The 4T1 cells implanted in the left-side fat pads of mice represented tumor cells in an out-field area of radiation. Since left-side fat pads were injected with 4T1 cells at day 7 (Fig. 2A), most tumors in the left-side fat pads began to be palpable on the 5th day after irradiation. The results showed that left-side tumor growth, such as that of right-side growth, was suppressed by all three radiation splitting schedules. As shown in Figure 2F and G, the tumor size was smaller in all three irradiated groups than in the control group; moreover, there was no difference in tumor sizes among the groups irradiated with 6 Gy × 3, 8 Gy × 2, and 13 Gy × 1 fractions. These data suggest that RT exhibits a mild systemic effect, regardless of fraction dose size and splitting schemes, in mice bearing 4T1 cells.
Lung metastasis is more efficiently inhibited by 13 Gy x1 or 8 Gy × 2 fractions than by 6 Gy × 3 fractions
To evaluate the effects of RT against metastasis, lung tissues were stained with H&E after sacrificing mice on day 25 after right-side 4T1 cell inoculation (Fig. 2A). When more than 15 tumor cells formed a definite cluster within lung tissue, the authors considered it indicative of a metastatic lesion (Fig. 3A). Metastatic percentage was calculated based on the proportion of metastatic area in the total lung area. Interestingly, radiation tended to decrease the proportion of lung metastasis (Fig. 3B). Particularly, mice treated with 13 Gy × 1 or 8 Gy × 2 fractions exhibited fewer lung metastases than mice treated with 6 Gy × 3 fractions (Fig. 3B).

13 Gy × 1 and 8 Gy × 2 fractions produced a better antimetastatic effect than 6 Gy × 3 fractions.
In vitro and in vivo HMGB1 expression patterns were similar
Since the authors observed increased inhibitory effects against lung metastasis from the 8 Gy × 2 and 13 Gy × 1 fractions, they investigated whether the release of DAMPs is related to this antimetastatic potential of RT. Since HMGB1 is one of the major DAMPs, the authors analyzed the expression of HMGB1 from the irradiated right-side tumor tissues. Interestingly, HMGB1 was expressed at a higher level in the 8 Gy × 2 and 13 Gy × 1 groups than in the 6 Gy × 3 group (Fig. 4A), and this in vivo HMGB1 expression is comparable with the HMGB1 expression levels observed in the in vitro experiments (Fig. 1C). These findings suggest that the radiation-induced antimetastatic effects are related to the increase of HMGB1 expression.
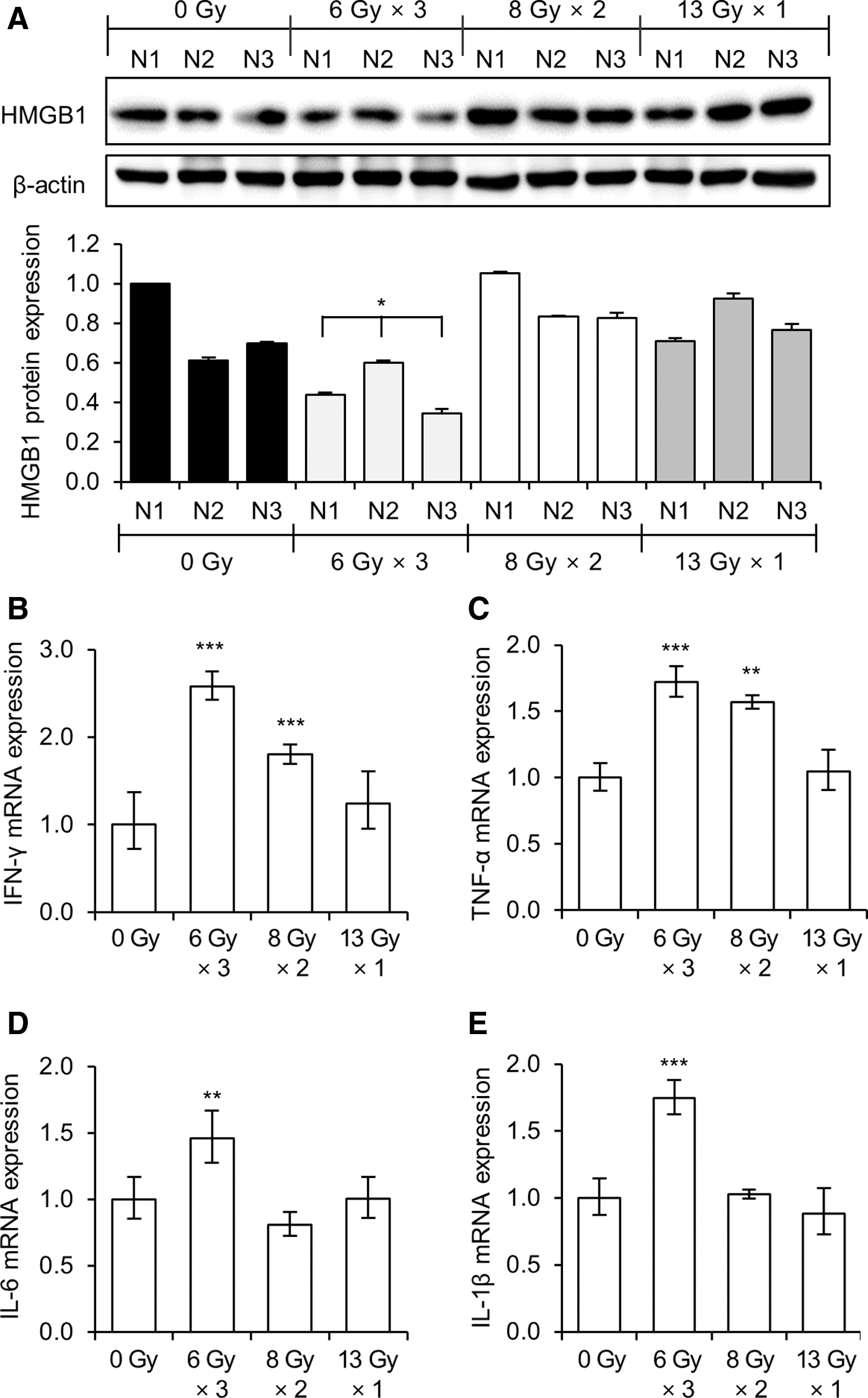
Expressions of HMGB1 and proinflammatory cytokines from the irradiated tumor tissues.
It is known that localized RT can stimulate the release of various cytokines and chemokines from tumor cells, providing an inflammatory tumor microenvironment. Thus, the mRNA expressions of several cytokines (IFN-γ, TNF-α, IL-6, and IL-1β) in the irradiated right-side tumor tissues were analyzed. In contrast to the expression of HMGB1, the expressions of IFN-γ, TNF-α, IL-6, and IL-1β were highly induced by the 6 Gy × 3 fraction radiation compared with the expression levels following the 13 Gy × 1 or 8 Gy × 2 radiation treatments (Fig. 4B–E).
Discussion
In this preclinical study, the authors investigated which RT fraction dose size and splitting schedule would provide improved systemic antitumor effects. When they designed this experimental study, the selection of irradiation splitting schedules was the biggest concern due to the possibility of mouse death caused by radiation-induced enteritis. Since the small bowel of the mouse is located closer to the target mass, more attention was required.
According to the American Association of Physics in Medicine (AAPM) Task Group 101 report, it is recommended that the maximal dose of small bowel at single-fraction stereotactic body RT should be less than 15.4 Gy, and the dose of 11.9 Gy should not be exceeded in the volume of 5 cc. For the three-fraction schedule, it is recommended that the maximal dose of small bowel should be less than 25.2 Gy, and the dose of 17.7 Gy should not be exceeded in the volume of 5 cc. 27 Furthermore, Song et al. reported that a 20 Gy single-fraction irradiation reduced mouse blood vessel size, 24 and Gong et al. reported that mice undergoing 14 Gy and 16 Gy irradiation on the abdomen died after 15 and 9 d, respectively. 23
Considering the AAPM Task Group 101 report and several published articles, the authors selected 13 Gy as the highest single-fraction dose and then applied the BED formula to determine three radiation splitting schedules: 6 Gy × 3, 8 Gy × 2, and 13 Gy × 1 fractions. Since the selected radiation splitting schedules were supposed to be isoeffective according to the BED formula; biological effect (E) = n(αd + βd 2 ), E/α = (nd) × (1 + d/(α/β)). This formula is based on the assumption that the cell response by irradiation follows a linear/quadratic model. The α and β are constants for linear and quadratic responses, respectively. Also, the n and d are total fraction number and one-fraction dose, respectively. E/α is BED, and 10 Gy was applied as α/β value, which is generally considered in cancer cells. They undertook cell viability experiments to confirm that the selected radiation splitting schedules were isoeffective; the results indicated that the 6 Gy × 3, 8 Gy × 2, and 13 Gy × 1 radiations were isoeffective.
When the right-side tumors of mice were irradiated with 6 Gy × 3, 8 Gy × 2, and 13 Gy × 1, the tumor sizes were significantly smaller than that of the unirradiated control group; moreover, there were no significant differences in tumor size among the three irradiated groups. These results confirm that the local tumor-controlling abilities of RT appear to be similar, regardless of fraction dose and splitting scheme.
In this study, the tumors in the left-side fat pad (unirradiated region) and the lung metastasis were considered representative of the presence of multiple lesions in a cancer patient. Thus, the authors measured tumor mass in unirradiated left-side fat pads and lung metastases to evaluate the systemic effects of RT. Similar to the local controlling effects of RT, the left-side tumor growth was also suppressed by all three radiation splitting schedules, regardless of fraction dose and splitting scheme. However, lung metastasis results indicated a greater level of inhibition in mice irradiated with the high-fraction doses (single 13 Gy dose or two fractions of 8 Gy) than that from RT with three fractions of 6 Gy. These results suggest that RT exhibits systemic effects as well as local controlling effects, and a high-fraction radiation dose may be more efficient for the control of distant metastases.
Interestingly, the RT effects on the expression of HMGB1 paralleled with its antimetastatic effects, displaying higher HMGB1 expression in tumors irradiated with a single 13 Gy dose or with two fractions of 8 Gy than in tumors irradiated with three fractions of 6 Gy. It is known that HMGB1 improves tumor antigen presentation by binding to the Toll-like receptor 4 (TLR4) on DCs, subsequently enhancing the function of T cells to destroy tumors. 6 –9 Thus, the higher expression of HMGB1 in tumors irradiated with 8 Gy × 2 and 13 Gy × 1 fractions may be correlated with the superior antimetastatic effects of those RT schemes.
Cytokines can exhibit dual functions in tumor progression, producing either anti- or protumorigenic effects. In modulating the abscopal effects of RT, it has been proposed that induction of proinflammatory cytokines in response to RT may summon immunogenic responses, resulting in tumor regression at out-field areas of RT. 28,29 In contrast to the antitumorigenic effects of cytokines via enhancement of immunogenic responses, the proinflammatory cytokines, such as IFN-γ, TNF-α, IL-6, and IL-1β, are reported to be important mediators of tumor growth and metastasis. 30,31 It is well known that upregulation of IFN-γ, TNF-α, IL-6, and IL-1β is associated with more aggressive phenotypes and stimulation of tumor growth in several types of cancers. 30 Thus, counterbalancing of the dual effects of cytokines as pro- or antitumorigenic may be correlated with the rarity of abscopal effects of RT. In fact, preclinical studies have often failed to show abscopal effects from RT alone, but RT in combination with immunotherapy has been shown to produce synergistic systemic antitumor effects by enhancing cytotoxic immunity and increasing the circulating levels of cytokines from activated immune cells. 4,19,29,32
In this study, the authors were unable to determine whether circulating cytokines are affected by the different fractionation schemes due to technical limitations in isolating immune cells or splenocytes. However, when they analyzed the expressions of proinflammatory cytokines from irradiated tumor tissues, IFN-γ, TNF-α, IL-6, and IL-1β were more highly induced by three fractions of 6 Gy rather than by a single dose of 13 Gy or two fractions of 8 Gy. Considering the better antimetastatic effects of RT in tumors irradiated with 8 Gy × 2 and 13 Gy × 1 fractions compared with those from the 6 Gy × 3 fractions, cytokines released within tumor tissues in response to RT may function as a protumorigenic and promote tumor growth and metastasis. In contrast to the role of cytokines within tumor tissues, there are possibilities that circulating cytokines may have antitumorigenic effects by boosting immunogenic responses. Thus, the role of circulating cytokines in the modulation of abscopal effects after RT needs to be investigated in future studies.
In recent years, immunotherapy has provided remarkable clinical responses in cancer patients and opened up new opportunities to synergize immunotherapeutic agents with RT and elicit durable abscopal effects. 10,29,32,33 However, the optimal dose and fractionation schemes of RT to induce abscopal effects have not been established, yet. Abuodeh et al. reviewed the literature published between 1969 and 2014 and reported that abscopal effects were observed in a total of 46 cases, where the radiation doses were variable from 0.45 Gy to 74.8 Gy. 34 Preclinical studies suggested that hypofractionation is required to elicit effective abscopal effects, but demonstrated mixed results regarding optimal fractionation.
Dewan et al. showed that 8 Gy × 2 and 13 Gy × 1 fractions were more effective for inducing abscopal effects compared with the single ablative dose of 20 Gy in combination with cytotoxic T lymphocyte-associated antigen 4 (CTLA-4). 15 Contrastingly, Lee et al. reported that a single ablative dose of radiation (20 Gy delivered in single fraction) generated strong CD8+ T cell-dependent immunity to lead to tumor reduction and eradication of metastasis, while 5 Gy × 4 fractions resulted in tumor recurrence because fractionated low-dose RT may continuously kill infiltrating effector T cells overtime. 35 Regarding a radiation dose, Schaue et al. demonstrated that the medium-size radiation doses of 7.5 Gy/fraction gave the best tumor control and tumor immunity. 17 Similarly, superiority in 8 Gy × 3 fractions in inducing the abscopal effect in the presence of anti-CTLA4 had been demonstrated. 15
In this study, the authors observed that 13 Gy × 1 or 8 Gy × 2 fractions are more effective than 6 Gy × 3 fractions in reducing distant metastases, which may result from induction of HMGB1 expression and suppression of proinflammatory cytokines from the irradiated tumor tissues. Since these RT schemes did not align with prior studies, there is limitation to generalize whether the single ablative dose of RT was more effective in inducing the abscopal response than a fractionated regimen. However, regarding a radiation dose, these data provide another evidence that the medium-size radiation doses, such as 8 Gy/fraction, would be effective for the abscopal effect induction.
Conclusions
To date, the optimal hypofractionated irradiation schedule to induce abscopal effects is still undefined. Since more evidence is required to establish consensus on the optimal dose and fractionation schemes of RT to maximize the abscopal responses, the authors hope that the results presented herein provide clues for optimizing RT dose regimens to make the abscopal effect clinically more relevant in future combination treatments.
Footnotes
Authors' Contributions
Y.S.K. and M.G.L. contributed equally to the conception, design, and acquisition of data, and wrote the article. J.Y.B performed experiments to acquire data and interpret data. K.S.N. contributed to the conception and design of data. J.I.L. performed experiments to acquire data and analyzed the data. S.Y.K. contributed to the conception and design of data and has been involved in drafting and writing the article. H.S.J. drafted and wrote the article and gave the final approval of the version to be published. All authors have reviewed and approved the published version of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the National Research Foundation of Korea (NRF) grants, funded by the Korea government (MISIT) (Grant No. 2016R1C1B1011783 and Grant No. NRF-2018R1D1A1B07049009).