
Abstract
Background:
Long noncoding RNA small nucleolar RNA host gene 3 (SNHG3) is related to the proliferation and metastasis of cancer cells. This study aims to reveal the role of SNHG3 in prostate cancer (PCa), which may help prevent PCa metastasis.
Methods:
SNHG3 plasmid, SNHG3 siRNA, miR-487a-3p mimic, miR-487a-3p inhibitor, TRIM25 plasmid, and TRIM25 siRNA were transfected or cotransfected into LNCaP and PC-3 cells. The proliferation, migration, and invasion of PCa cells were measured by Cell Counting Kit-8, wound-healing, and transwell assays, respectively. The expressions of SNHG3, miR-487a-3p, E-cadherin, N-cadherin, Snail, and TRIM25 in PCa tissues and cells were measured by quantitative reverse transcription polymerase chain reaction or Western blot.
Results:
SNHG3 expression level was upregulated in PCa tissues and cells. SNHG3 overexpression and miR-487a-3p inhibitor promoted cell viability, migration, invasion, and N-cadherin and Snail levels, and inhibited E-cadherin level in LNCaP cells, while SNHG3 silencing and miR-487a-3p mimic had the opposite effects on PC-3 cells. The inhibitory effect of miR-487a-3p mimic on the migration, invasion, and epithelial-mesenchymal transition (EMT) of LNCaP cells was inversed by both SNHG3 and TRIM25 plasmids. Similarly, the function of miR-487a-3p inhibitor in PC-3 cells was also inversed by SNHG3 siRNA and TRIM25 siRNA.
Conclusion:
SNHG3 mediates PCa migration, invasion, and EMT by sponging miR-487a-3p to regulate TRIM25. The Clinical Trial Registration number: Y20180831.
Introduction
The incidence of prostate cancer (PCa) in men ranks the second among all cancers in the world, which seriously threatens men's health. 1 Statistics show that as many as 250,000 patients die each year from PCa. 2 Although androgen-blocking therapy has achieved a certain effect in patients with PCa, some patients eventually remain uncured with worsened tumor metastasis. 3 Once distant metastases occur in PCa, such as bone metastases and lung metastases, the 5-year survival rate of patients will decrease significantly to 27.9%. 4 However, the mechanism of distant metastasis of PCa is still not clear, and the current treatment method has multiple limitations. Hence, clinical research on PCa is urgently required for treating PCa.
The occurrence of PCa is usually accompanied with gene imbalance in cells. 5 In recent years, the potential role of long noncoding RNA (lncRNA) in regulating the function of malignant tumor cells has received widespread attention. 6 Increasing evidence indicates that lncRNAs are involved in many biological behaviors, including proliferation, metastasis, differentiation, inflammation, angiogenesis, and metabolism. 7
Small nucleolar RNA host gene (SNHG) is a type of lncRNA that plays a crucial role in the biological regulation of malignant tumors. 8 SNHG3, which is located in 1p31, 9 has been reported to be highly expressed in patients with liver cancer, osteosarcoma, colorectal cancer, and breast cancer, and high expression of SNHG3 is associated with poor prognosis of patients. 10 –12 SNHG3 overexpression can promote human colorectal cancer cell proliferation and mice tumor growth. 11 SNHG3 has also been reported to be implicated in the occurrence and development of many malignancies, 9 but there are few reports on its function and mechanism in PCa. Therefore, this study investigated the effect of SNHG3 on the proliferation and metastasis of PCa cells by overexpressing or silencing the SNHG3 gene in PCa cells.
Recently, lncRNA has been reported to act as a ceRNA via sponging miRNA and releasing the target mRNA from specific miRNA. 13 The target microRNA of SNHG3 as well as the target gene of the microRNA may reveal the mechanism, by which SNHG3 affects PCa cell proliferation and metastasis. In previous study, miR-487a-3p expression levels had been found to be downregulated in PCa tissue and cells, 14 and related in many functions, including cancer cells metastasis, proliferation, and so on, 14,15 however, the mechanism of action of miR-487a-3p in PCa remains to be further explored.
Therefore, in this study, the authors have further explored the effect of miR-487a-3p and SNHG3 on PCa progress. In addition, TRIM25 is also reported having an increased expression in PCa tissues, 16,17 and the mechanism of TRIM25 in PCa development has not been studied in detail, so they further explored the effect of TRIM25 and miR-487a-3p on PCa tissue.
A deeper understanding of the possible molecular mechanisms that regulate the migration, invasion, and epithelial-mesenchymal transition (EMT) of PCa will be conducive to further developing targeted interventions, and thereby providing new ideas and theoretical foundations for clinical prevention of PCa metastasis.
Materials and Methods
Preparation of clinical samples
A total of 50 pairs of PCa tissue and adjacent noncancerous tissue samples were acquired from PCa patients undergoing radical prostatectomy in Shaoxing People's Hospital from 2014 to 2018. Corresponding adjacent tissues were collected from the prostate at least 1 cm away from the cancer site.
Ethical statement
All experiments in the current study were approved by the Ethics Committee of Shaoxing People's Hospital (Y20180831). Besides, written informed consent forms were signed by each subject.
Cell culture
Human normal prostatic epithelial cell line RWPE-1 (CBP60347; Nanjing Cobioer biotechnology Co. Ltd.) and human PCa cell lines PC-3 (CBP60343; Nanjing Cobioer biotechnology Co. Ltd.), 22RV1 (CRL-2505; ATCC), DU145 (CBP60345; Nanjing Cobioer biotechnology Co. Ltd.), and LNCaP (CBP60346; Nanjing Cobioer biotechnology Co. Ltd.) were cultured in DMEM high-glucose medium (12800017; Solarbio) supplemented with 10% fetal bovine serum (FBS, YZ-1270548; Solarbio) and 1% penicillin–streptomycin (P1400; Solarbio), in a humidified atmosphere with 5% CO2 at 37°C. The medium was changed every 3 d.
Transient transfection
SNHG3 plasmid (SNHG3), SNHG3 siRNA (siSNHG3), miR-487a-3p mimic (Mimic, miR10002178-1-5, Guangzhou RiboBio Co., Ltd.), miR-487a-3p inhibitor (Inhibitor, miR20002178-1-5, Guangzhou RiboBio Co., Ltd.,), TRIM25 plasmid (TRIM25), or TRIM25 siRNA (siTRIM25) were transfected or cotransfected into LNCaP and PC-3 cells by using Lipofectamine 2000 transfection reagent (11668019; Thermo Fisher Scientific) according to the manufacturer's instructions.
In brief, 50 μL of DMEM medium (11668027; Invitrogen) was used to dilute 0.8–1.0 μg of DNA. Another 50 μL of DMEM medium was used to dilute 1–3 μL of Lipofectamine 2000 reagent for 5 min at 25°C. Then the diluted DNA and Lipofectamine 2000 reagent were mixed and placed for 20 min at 25°C, followed by 48 h of incubation at 37°C with 5% CO2. After 24–72 h, gene activity was assessed for further experiment. The sequence of SNHG3 and TRIM25, as well as the negative controls (listed in Table 1), were designed with siDirect version 2.0 and synthesized by GenePharma Technologies.
The siRNA and the Negative Control Designed by siDirect Version 2.0
SNHG3, small nucleolar RNA host gene 3; TRIM25, tripartite motif containing 25.
Quantitative reverse transcription polymerase chain reaction
The levels of lncRNA SNHG3 and microRNA-487a-3p were measured by quantitative reverse transcription polymerase chain reaction (qRT-PCR). The extraction of RNA from tissues or cells was performed with the TRIzol reagent (15596018; Thermo Fisher Scientific) according to the manufacturer's instruction. Then, the purity and concentration of the extracted RNA solution were measured with the NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific), and RNA integrity was assessed using agarose gel electrophoresis (G442001; Thermo Fisher Scientific). Real-time PCR was conducted using SYBR Green Real-Time PCR Master Mix (QPK-201, TOYOBO CO., Ltd.) and the LightCycler 480 qPCR System (Roche Molecular Systems, Inc.). The primers listed in Table 2 were designed with PrimerBank, online tools GETPrime or Primer3 Plus, while the primers of U6 were determined by referring to the previous report. 14 All the above primers were synthesized by Thermo Fisher.
The Specific Primer for the Quantitative Reverse Transcription Polymerase Chain Reaction
SNHG3, small nucleolar RNA host gene 3; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
RNA reverse transcription was conducted with a PrimeScript RT Reagent Kit (RR037B; Takara Bio USA, Inc.) with gDNA Eraser (RR047B; Takara). First, 5 × gDNA Eraser Buffer (2 μL), gDNA Eraser (1 μL), total RNA (1 μg), and RNase Free ddH2O (10 μL) were added in Applied Biosystems Veriti PCR (4484073; Thermo Fisher Scientific) and incubated for 2 min at 42°C. Then, PrimeScript RT Enzyme Mix I (1 μL), RT primer Mix (4 μL), 5 × PrimeScript Buffer 2 (4 μL), and RNase Free ddH2O (1 μL) were added into sample (10 μL) and incubated for 15 min at 37°C, followed by 5 seconds at 85°C. PCR was performed using Thermo Fisher Scientific 7500 Real-Time PCR System (4351105; Thermo Fisher Scientific) under the following conditions: 95°C for 5 s and 60°C for 30 s for a total of 40 cycles. The data were analyzed by the 2−ΔΔ Ct method.
The results were normalized to the expression of GAPDH or U6.
Cell Counting Kit-8 assay
The proliferation of LNCaP and PC-3 cells was assessed with a Cell Counting Kit-8 (CCK-8) (M4839; Abmole Bioscience, Inc.). In brief, 2 × 103 cells were seeded into 96-well plates in triplicate and cultured for 24, 48, or 72 h at 37°C. Then 10 μL of CCK-8 solution was added to each well and then the plates were incubated at 37°C for 1 h. The optical density (450 nm) of each well was measured by using a microplate reader (RNE-90002; Shenzhen rongjin technology Co. Ltd).
Wound healing
The migration rates of LNCaP and PC-3 cells were assessed using wound healing assay. The LNCaP and PC-3 cells were trypsinized and planted in 60 mm Petri dishes. When the cell density reached 90% or higher, a 200 mL sterile pipette tip was used to gently create a scratch in the Petri dish. LNCaP and PC-3 cells were rinsed with PBS and then cultured in serum-free medium. The healing process of the scratch was recorded using an inverted microscope (IXplore Standard; OLYMPUS CORPORATION) at 0 and 24 h, and the migration of LNCaP and PC-3 cells was evaluated by the percentage of wound area.
Transwell assay
The invasion rates of LNCaP and PC-3 cells were assessed using Transwell assay.
Matrigel gel (M8370; Solarbio) diluted at 1: 8 was added to the Transwell chamber (3413; Corning), and incubated for 30 min at 37°C. The BD chamber was placed in a 24-well plate. The LNCaP and PC-3 cells in logarithmic growth phase were selected, digested, and resuspended in serum-free medium at the concentration of 1.5 × 107/mL. Then, 400 μL of the cell suspension was added into the upper layer of the chamber, and 800 μL of normal medium was added into the lower wells. After 48 h of incubation, the uninvaded LNCaP or PC-3 cells in the upper chamber were wiped off and the invaded cells were stained with 0.25% crystal violet (IC0600; Solarbio). The invaded cells were counted and typical images were captured with a microscope (100 × ) (Eclipse Ti2; Nikon).
Western blot assay
Expression of EMT-related proteins (E-cadherin, N-cadherin, and Snail) and TRIM25 were measured by Western blot. Proteins in LNCaP and PC-3 cells were extracted by using Western and IP cell lysates (P0013J; Beyotime) followed by 12,000 rpm centrifugation (10 min, 4°C). Then 40 μg of protein lysates were separated by SDS-PAGE and transferred to a polyvinylidene fluoride (PVDF) membrane (YA1701; Solarbio). After blocking with skimmed milk for 1 h, the PVDF membrane was incubated with the primary antibodies E-Cadherin Mouse mAb (#14472; Cell Signaling Technology), N-Cadherin Mouse mAb (#14215; Cell Signaling Technology), Mouse Anti-TRIM25/EFP antibody (ab88669; Abcam), Goat Anti-SNAIL antibody (ab53519; Abcam), and Mouse Anti-GAPDH antibody (ab8245; Abcam) overnight.
Then the PVDF membrane was incubated with the secondary antibody Goat Anti-Mouse IgG H&L (ab205719; Abcam) or Donkey anti-Goat IgG (A32849; Thermo Fisher) at room temperature for 2 h. Then the immunodetection was performed with enhanced chemiluminescence substrate (PE0020; Solarbio). Protein levels were normalized to GADPH level using ImageJ (Wayne Rasband).
Bioinformatics analysis
The Cancer Genome Atlas (TCGA) was used to analyze the relationship between SNHG3 expression and PCa. Binding sites between miR-487a-3p and the SNHG3 or TRIM25 gene were analyzed through an algorithm on the website of starBase.
Dual-luciferase reporter assay
Dual-luciferase reporter assay were conducted to verify the interaction miR-487a-3p with SNHG3 or TRIM25. In LNCaP and PC-3 cells, miR-487a-3p binding sites with SNHG3 or TRIM25, including mutant type (MUT) and wild type (WT), were analyzed using dual-luciferase reporter assay based on protocol of Dual-Luciferase Reporter Assay System (11402ES60/80; Yeasen Biotech Co., Ltd.). In brief, the 3′-UTR of SNHG3 contains the highly conserved binding sites of miR-487a-3p, and the sequence containing the binding sites is as follows: 5′-GCCCGGCCACTTTTGTATGATT-3′. The 3′-UTR of TRIM25 contains binding sites of miR-487a-3p, and the sequence containing the binding sites is as follows: 5′-TCTCATTTTGCAGAATATGATG-3′.
Furthermore, the 3′-UTR sequence was inserted into the pmirGLO Vector (Promega) with XhoI and XbaI double digestion to construct the recombinant dual-luciferase reporter vector. Meanwhile, a plasmid containing the mutant SNHG3 3′-UTR, pmirGLO-Mut, was generated by mutating the core sequence of the miR-487a-3p binding sites through DNA synthesis (Sangon Biotech Co. Ltd., Shanghai, China), and the sequence was as follows: 5′-GCCCGGCCACTTTGAGCCTGGT-3′. Meanwhile, the plasmid of TRIM25 3′-UTR Mut was synthesized as same with mutant SNHG3 3′-UTR, and the sequence was as follows: 5′-TCTCATTTTGCAGAAGGCACGG-3′. LNCaP and PC-3 cells were cotransfected with psicheck2 luciferase vectors containing the 3′ untranslated region of target genes with SNHG3 MUT or WT and miR-487a-3p mimic or inhibitor.
The expression of luciferase in the LNCaP or PC-3 cells extract was measured after 48 h according to the instruction of the assay kits. After aspirating medium, LNCaP or PC-3 cells were washed with PBS. Then 1 × Passive lysis buffer (PLB) 150 μL was added to ensure complete and even coverage of the cell monolayer and the plates were rocked for 15 min at 25°C. Luciferase Assay Reagent II 100 μL was dispensed into luminometer tubes and carefully transferred 20 μL PLB. The tube was placed in the luminometer (E5311; Promega Corporation) and initiated reading. After reading, 100 μL Stop & Glo® Reagent (E314B-C; Promega Corporation) was dispensed and then measured again.
Statistical analysis
All experiments were repeated at least three times. All data are presented as mean ± standard error of the mean (SEM). Statistical analyses were conducted with SPSS 18.0 (IBM SPSS) and GraphPad Prism 6.0 (GraphPad Software, Inc.). One-way analysis of variance (ANOVA) was used to analyze the significant differences among all results, and p < 0.05 was considered statistically significant.
Results
SNHG3 expression level was upregulated in tissue and cells of PCa
TCGA data showed that SNHG3 expression was significantly upregulated in the PCa tissue (Fig. 1A). To verify SNHG3 expression in PCa cells, the authors quantitatively analyzed the SNHG3 expression level in PCa tissues, adjacent normal tissue specimens, human PCa cell lines (PC-3, 22RV1, DU145, and LNCaP), and human normal prostatic epithelial cell line RWPE-1 using qRT-PCR assay. The details in Figure 1B demonstrated that the relative SNHG3 expression in PCa tissue was significantly higher than that in adjacent tissue (p < 0.001).

SNHG3 was upregulated in tissue and cells of PCa
The results in Figure 1C also show that the relative SNHG3 level in human PCa cell lines (PC-3, 22RV1, DU145, and LNCaP cells) was significantly higher than that in the human normal prostatic epithelial cell line RWPE-1 (p < 0.01 or p < 0.001). Besides, the PC-3 cells and LNCaP cells had the highest and lowest SNHG3 expression, respectively, among the four human PCa cell lines. These results showed that SNHG3 expression level was upregulated in both PCa tissue and cells.
SNHG3 overexpression promoted the viability and migration rates of LNCaP cells, while SNHG3 silencing had an opposite effect on PC-3 cells
To assess the effect of SNHG3 on PCa cells, the SNHG3 expression levels, the OD values and the migration rates of LNCaP and PC-3 cells were measured by qRT-PCR, CCK-8, and wound healing assay, respectively, after transfection of SNHG3 siRNA, SNHG3 plasmid, or the negative control.
As it can be seen in Figure 2A and B, SNHG3 plasmid markedly upregulated the SNHG3 expression level in LNCaP cells, and SNHG3 siRNA markedly downregulated the SNHG3 expression level in PC-3 cells (p < 0.001, respectively). Figure 2C and D exhibit that the OD value of SNHG3 plasmid-transfected LNCaP cells was significantly greater than that of LNCaP cells transfected with the negative control at 24, 48, and 72 h (p < 0.05, p < 0.01 or p < 0.001), and the OD value of SNHG3 siRNA-transfected PC-3 cells was significantly lower than that of PC-3 cells transfected with the siRNA negative control at 48 and 72 h (p < 0.01 or p < 0.001).
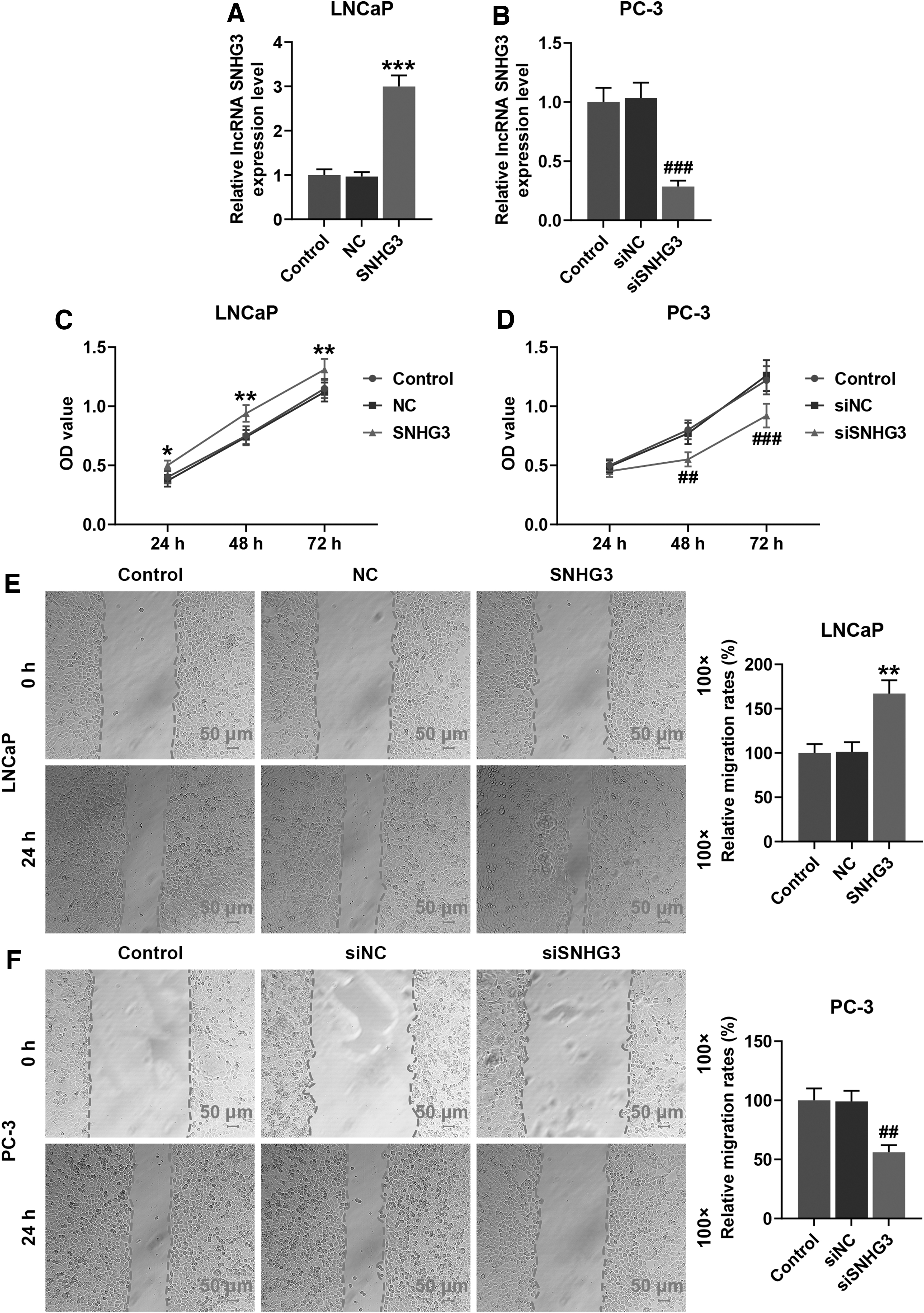
SNHG3 overexpression promoted the cell viability and migration rates in LNCaP cells, and SNHG3 silence had opposite effect in PC-3 cells.
Similarly, the results in Figure 2E and F show that the migration rate of SNHG3 plasmid-transfected LNCaP cells was markedly higher than that of LNCaP cells transfected with the negative control at 24 h (p < 0.01), and the migration rate of SNHG3 siRNA-transfected PC-3 cells was markedly lower than that of PC-3 cells transfected with the SNHG3 siRNA negative control at 24 h (p < 0.01).
These results demonstrated that SNHG3 overexpression promoted the viability and migration rates of LNCaP cells, and SNHG3 silencing inhibited the viability and migration rates of PC-3 cells.
SNHG3 overexpression promoted the invasion and EMT of LNCaP cells, while SNHG3 silencing had an opposite effect on PC-3 cells
To assess the invasion and EMT of LNCaP and PC-3 cells, the invasion rates and the expression of EMT related proteins (E-cadherin, N-cadherin, and Snail) in LNCaP and PC-3 cells were measured by Transwell assay and Western blot, respectively. As illustrated in Figure 3A, SNHG3 plasmid markedly upregulated the invasion rate of LNCaP cells (p < 0.05), and SNHG3 siRNA markedly downregulated the invasion rate of PC-3 cells (p < 0.01). According to Figure 3C, SNHG3 plasmid-transfected LNCaP cells displayed a significantly upregulated E-cadherin level and significantly downregulated levels of N-cadherin and Snail (p < 0.001, respectively). On the contrary, Figure 3D shows that SNHG3 siRNA markedly upregulated the E-cadherin level and downregulated the levels of N-cadherin and Snail in PC-3 cells (p < 0.001, respectively).

SNHG3 overexpression promoted the cells invasion and EMT relative protein level in LNCaP cells, while SNHG3 silence had opposite effect in PC-3 cells.
These results suggest that SNHG3 overexpression promoted the invasion and EMT of LNCaP cells, and SNHG3 silencing inhibited the invasion and EMT of PC-3 cells.
SNHG3 acted as a molecular sponge of miR-487a-3p
Then, the authors performed bioinformatics analysis with starBase to select candidate miRNAs that could bind to SNHG3 for understanding the underlying mechanism of SNHG3 in PCa. In Figure 4A, it was found that miR-487a-3p could bind to SNHG3. The results of dual-luciferase reporter assay implied that cotransfection of SNHG3 WT and miR-487a-3p mimics significantly decreased the luciferase activities in LNCaP or PC-3 cells, compared with cotransfection of SNHG3 MUT and miR-487a-3p mimics (Fig. 4B, C, p < 0.001).
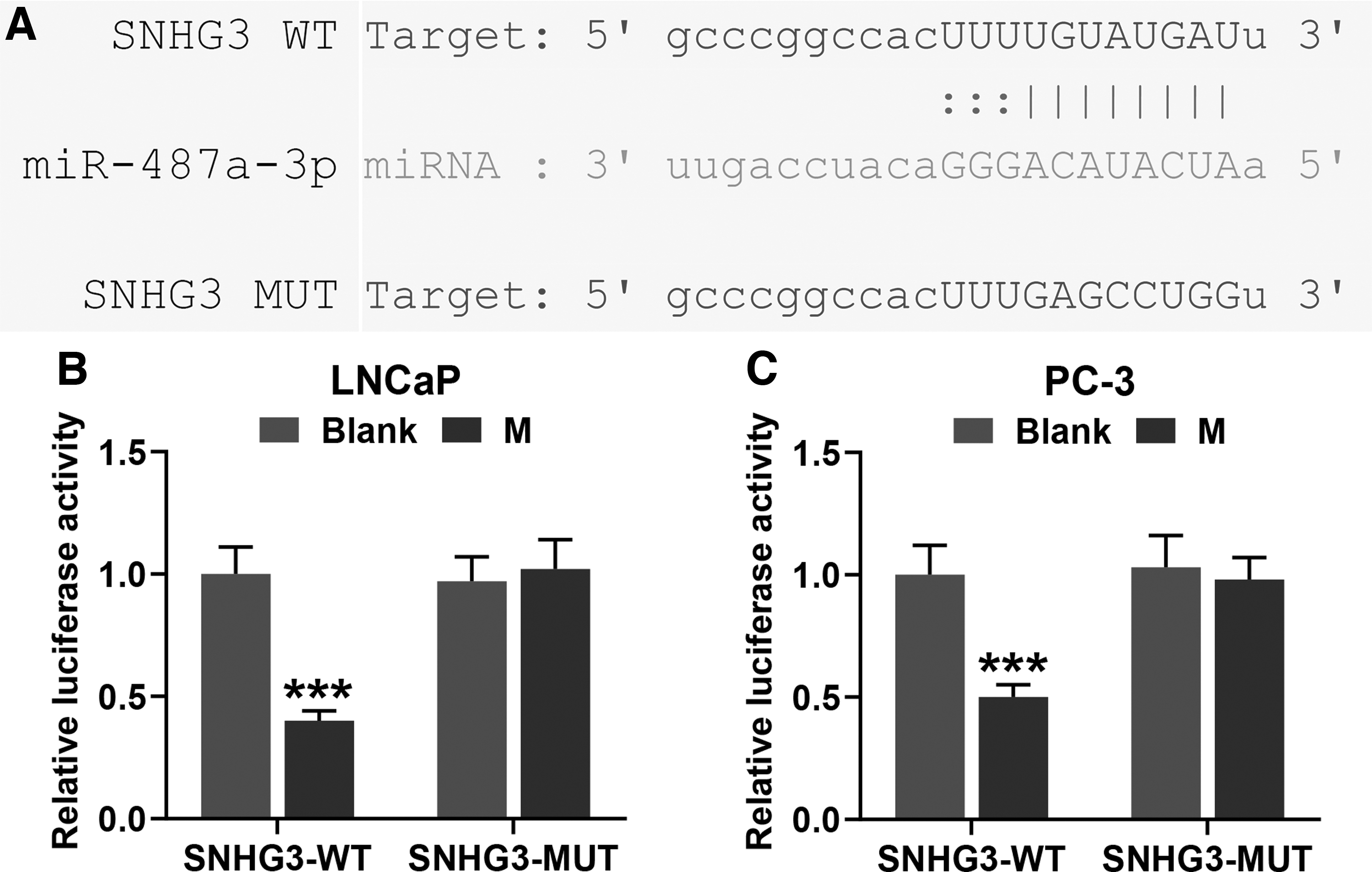
SNHG3 acted as a molecular sponge for miR-487a-3p in LNCaP cells and PC-3 cells.
SNHG3 increased the viability and migration rates of LNCaP and PC-3 cells by regulating miR-487a-3p expression
To assess the effect of SNHG3 on PCa cells, the miR-487a-3p expression levels, the OD values, and the migration rates of LNCaP cells and PC-3 cells were measured by qRT-PCR, CCK-8, and wound healing assay, respectively, after cotransfection of SNHG3 plasmid or SNHG3 siRNA and miR-487a-3p inhibitor or mimic.
According to Figure 5A and B, in LNCaP cells and PC-3 cells, the miR-487a-3p expression level was upregulated by miR-487a-3p mimic and SNHG3 siRNA and downregulated by SNHG3 plasmid and miR-487a-3p inhibitor, and miR-487a-3p mimic and inhibitor inversed the functions of SNHG3 plasmid and SNHG3 siRNA, respectively (p < 0.01, respectively). On the contrary, the details in Figure 5C and D show that the OD values of LNCaP cells and PC-3 cells were markedly decreased by miR-487a-3p mimic and SNHG3 siRNA and markedly increased by SNHG3 plasmid and miR-487a-3p inhibitor, and miR-487a-3p mimic and inhibitor markedly inversed the functions of SNHG3 plasmid and SNHG3 siRNA, respectively (p < 0.05, p < 0.01 or p < 0.001).
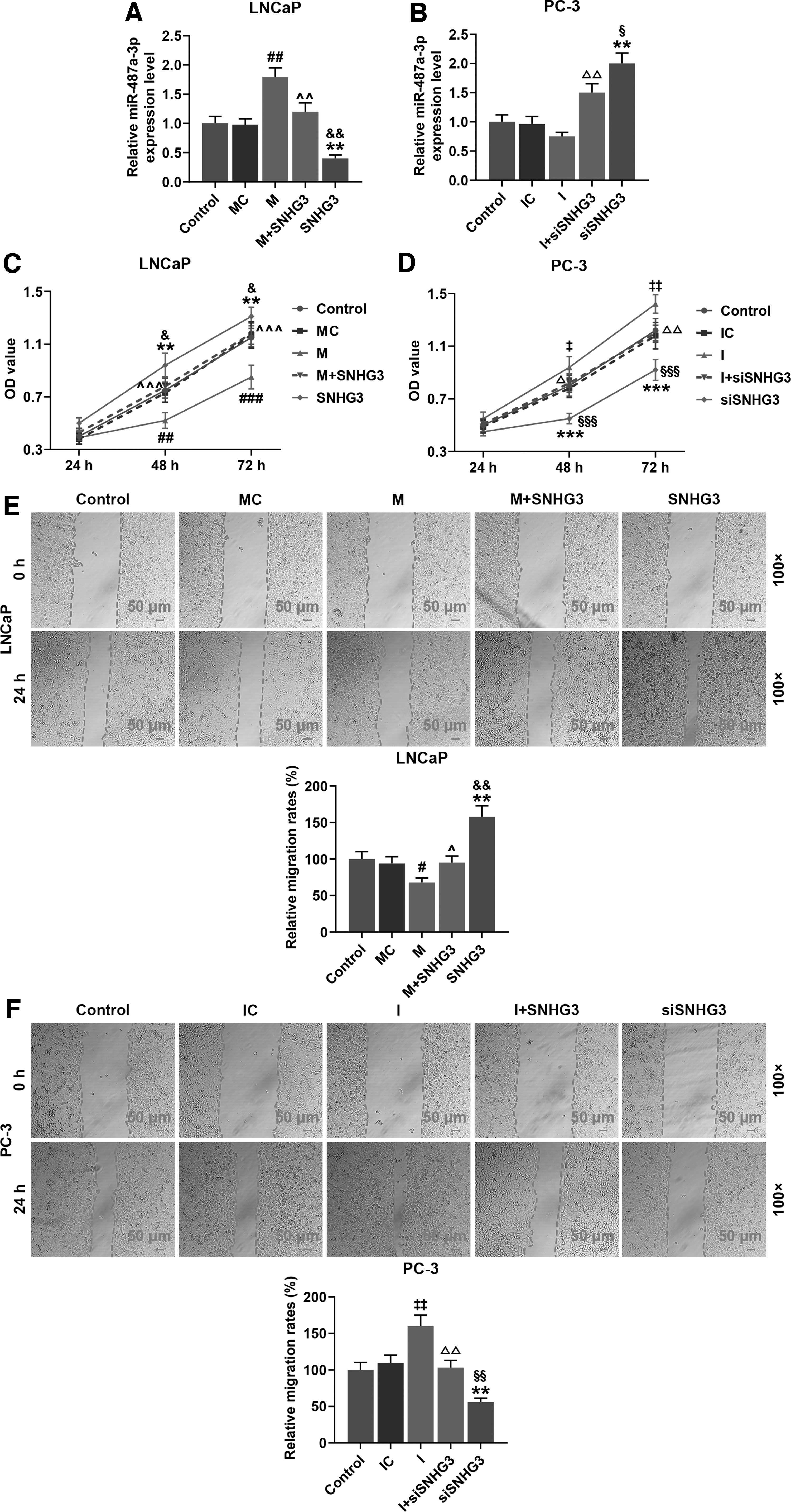
SNHG3 increased the cell viability and migration rates by regulating miR-487a-3p expression in LNCaP and PC-3 cells.
Similarly, Figure 5E and F illustrate that the migration rates of LNCaP cells and PC-3 cells were markedly decreased by miR-487a-3p mimic and SNHG3 siRNA and markedly increased by SNHG3 plasmid and miR-487a-3p inhibitor, and that miR-487a-3p mimic and inhibitor markedly inversed the functions of SNHG3 plasmid and SNHG3 siRNA, respectively (p < 0.05 or p < 0.01). These results indicate that SNHG3 promotes PCa cell proliferation and migration by sponging miR-487a-3p.
SNHG3 mediated the EMT and invasion of LNCaP and PC-3 cells by regulating miR-487a-3p level
To further explore the effects of SNHG3 on PCa metastasis, they measured the invasion rates and the expression of EMT-related proteins (E-cadherin, N-cadherin, and Snail) in LNCaP cells and PC-3 cells using Transwell assay and Western blot, respectively. As can be seen in Figure 6A and B, the invasion rates of LNCaP cells and PC-3 cells were markedly decreased by miR-487a-3p mimic and SNHG3 siRNA and increased by SNHG3 plasmid and miR-487a-3p inhibitor, and miR-487a-3p mimic and inhibitor markedly inversed the functions of SNHG3 plasmid and SNHG3 siRNA, respectively (p < 0.05, p < 0.01 or p < 0.001). As illustrated in Figure 6C, in LNCaP cells, miR-487a-3p mimic significantly upregulated E-cadherin level and downregulated the levels of N-cadherin and Snail; SNHG3 plasmid significantly downregulated E-cadherin level and upregulated the levels of N-cadherin and Snail; and miR-487a-3p mimic markedly inversed the function of SNHG3 plasmid (p < 0.001, respectively). Oppositely, Figure 6D shows that in PC-3 cells, miR-487a-3p inhibitor significantly downregulated E-cadherin level and upregulated the levels of N-cadherin and Snail; SNHG3 siRNA significantly downregulated E-cadherin level and upregulated the levels of N-cadherin and Snail; and miR-487a-3p inhibitor markedly inversed the function of SNHG3 plasmid (p < 0.001, respectively). These results indicate that SNHG3 mediated the EMT and invasion of LNCaP and PC-3 cells by regulating miR-487a-3p level.

SNHG3 mediated EMT and invasion of PCa cells by regulating miR-487a-3p level in LNCaP and PC-3 cells.
TRIM25 acted as the target gene of miR-487a-3p
The authors used the prediction software program starBase to screen the candidate target gene of miR-487a-3p. As shown in Figure 7A, TRIM25 was the target gene of miR-487a-3p. The results of dual-luciferase reporter assay showed that cotransfection of TRIM25 WT and miR-487a-3p mimics significantly downregulated the luciferase activities in LNCaP or PC-3 cells, compared with cotransfection of TRIM25 MUT and miR-487a-3p mimics (Fig. 7B, C, p < 0.001). These results mean that TRIM25 acts as the target gene of miR-487a-3p.

TRIM25 acted as a target gene of miR-487a-3p in LNCaP and PC-3 cells.
SNHG3 increased TRIM25 level by regulating miR-487a-3p expression in LNCaP and PC-3 cells
To further verify the prediction that TRIM25 was the target of miR-487a-3p, TRIM25 levels in LNCaP and PC-3 cells were measured by Western blot and qRT-PCR after cotransfection of miR-487a-3p mimic or inhibitor and TRIM25 plasmid or siRNA. Figure 8A and B show that in LNCaP cells, the TRIM25 expression level was downregulated by miR-487a-3p mimic and upregulated by SNHG3 plasmid, and miR-487a-3p mimic attenuated the function of SNHG3 plasmid (p < 0.05, p < 0.01 or p < 0.001); in PC-3 cells, the TRIM25 expression level was upregulated by miR-487a-3p inhibitor and downregulated by SNHG3 siRNA, and miR-487a-3p inhibitor inversed the function of SNHG3 siRNA (p < 0.05, p < 0.01 or p < 0.001). These results indicate that SNHG3 promotes TRIM25 level by sponging miR-487a-3p in LNCaP and PC-3 cells.

SNHG3 increased the TRIM25 level by regulating miR-487a-3p expression in LNCaP and PC-3 cells.
TRIM25 overexpression inversed the promoting effect of miR-487a-3p mimic on PCa cell viability and migration, while TRIM25 silencing inversed the inhibitory effect of miR-487a-3p inhibitor on PCa cell viability and migration
To further explore the function of TRIM25 in PCa cell migration, the miR-487a-3p expression level, the OD values and the migration rates of LNCaP and PC-3 cells were measured by qRT-PCR, CCK-8, and wound healing assay, respectively.
According to Figure 9A and B, in LNCaP cells and PC-3 cells, the miR-487a-3p expression level was upregulated by miR-487a-3p mimic and SNHG3 siRNA and downregulated by SNHG3 plasmid and miR-487a-3p inhibitor, and miR-487a-3p mimic and inhibitor inversed the functions of SNHG3 plasmid and SNHG3 siRNA, respectively (p < 0.05 or p < 0.01). On the contrary, the details in Figure 9C and D show that the OD values of LNCaP cells and PC-3 cells were markedly decreased by miR-487a-3p mimic and SNHG3 siRNA and markedly increased by SNHG3 plasmid and miR-487a-3p inhibitor, and miR-487a-3p mimic and inhibitor markedly inversed the functions of SNHG3 plasmid and SNHG3 siRNA, respectively (p < 0.05, p < 0.01 or p < 0.001).

TRIM25 overexpression inversed the miR-487a-3p mimic promoting PCa cells viability and migration, while TRIM25 silence inversed the inhibit effect of miR-487a-3p inhibitor.
Similarly, as shown in Figure 9E and F, the migration rates of LNCaP cells and PC-3 cells were markedly decreased by miR-487a-3p mimic and SNHG3 siRNA and markedly increased by SNHG3 plasmid and miR-487a-3p inhibitor, and miR-487a-3p mimic and inhibitor markedly inversed the functions of SNHG3 plasmid and SNHG3 siRNA, respectively (p < 0.05, p < 0.01 or p < 0.001). These results mean that TRIM25 overexpression inhibited the viability and migration of LNCaP cells, while silencing of TRIM25 promoted the viability and migration of PC-3 cells.
TRIM25 overexpression inversed the promoting effect of miR-487a-3p mimic on PCa cell invasion and EMT, while TRIM25 silencing had the opposite effect of miR-487a-3p inhibitor
To test the effect of TRIM25 on the invasion ability of LNCaP and PC-3 cells, the invasion rates and the expressions of EMT-related proteins (E-cadherin, N-cadherin, and Snail) were measured by Transwell assay and Western blot, respectively. As illustrated in Figure 10A and B, SNHG3 plasmid markedly upregulated the invasion rate of LNCaP cells (p < 0.01, respectively) and SNHG3 siRNA markedly downregulated the invasion rate of PC-3 cells (p < 0.01, respectively). According to Figure 10C, SNHG3 plasmid-transfected LNCaP cells exhibited a significantly upregulated E-cadherin level and significantly downregulated levels of N-cadherin and Snail (p < 0.001, respectively). On the contrary, the details in Figure 10D show that SNHG3 siRNA markedly upregulated the E-cadherin level and downregulated the levels of N-cadherin and Snail in PC-3 cells (p < 0.001, respectively). To sum up, SNHG3 overexpression promoted the invasion ability of LNCaP cells, and SNHG3 silencing inhibited the invasion ability of PC-3 cells.
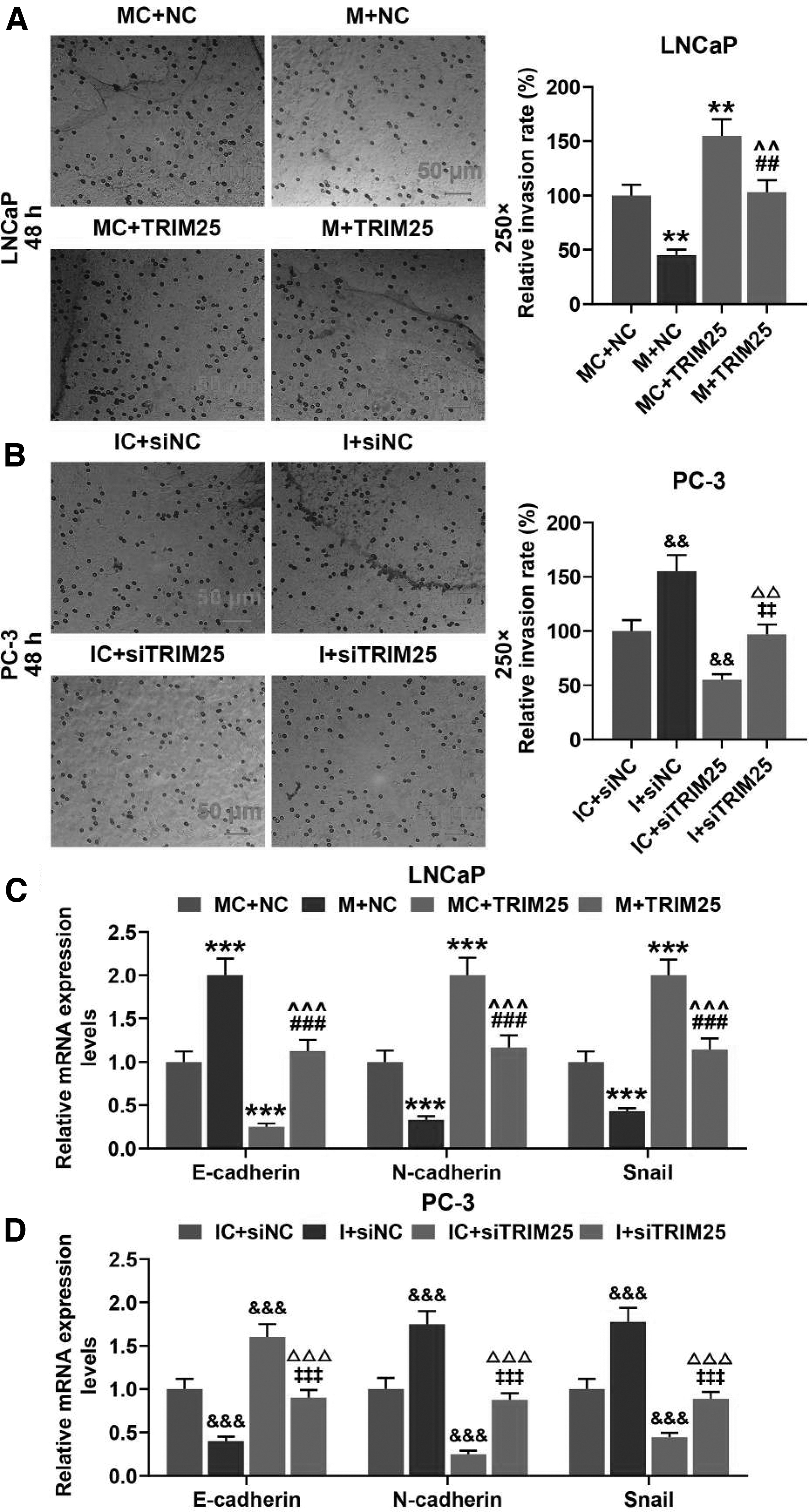
TRIM25 overexpression inversed invasion and EMT promoting effect of miR-487a-3p mimic and TRIM25 silence had opposite effect for miR-487a-3p inhibitor.
Discussion
At present, distant metastasis of PCa is the main cause of death in male patients with PCa. Researchers believe that EMT and mesenchymal-epithelial transformation is the key step in the long-range metastasis of tumor cells. 18 PCa cells have a strong ability to migrate and invade through EMT, which enable them to enter the human circulatory system through the cell basement membrane and vascular endometrium and then pass through the vascular endometrium and cell basement membrane in a certain part of the body and form a new metastatic lesion. 19 Therefore, exploring genes or biomolecules related to PCa cell EMT can provide new markers for the prognosis of PCa, as well as a new target for PCa treatment.
There is increasing evidence that lncRNA plays important roles in the progression of cancer diseases. 20,21 LncRNA SNHG3 has been reported to be highly expressed in patients with gastric cancer. 9 Similarly, it was found that SNHG3 was overexpressed in 50 pairs of PCa specimens and human PCa cell lines (PC-3, 22RV1, DU145, and LNCaP) compared with the adjacent tissue samples and the human normal prostatic epithelial cell lines RWPE-1, respectively.
In the present study, LNCaP cells and PC-3 cells were chosen to be transfected with SNHG3 plasmid or SNHG3 siRNA given that SNHG3 had the highest and lowest expression levels in the two cells, respectively. After transfection, it was found that overexpression of SNHG3 promoted the viability, migration rate, and invasion rate of LNCaP cells, while silencing of SNHG3 had an opposite effect of SNHG3 overexpression on PC-3 cells, which is consistent with the findings in Xuan's study on gastric cancer 9 and Huang's study on colorectal cancer. 11 E-cadherin, N-cadherin, and Snail are the indexes of EMT, 10,22 and thus e authors measured the levels of the three proteins to assess the EMT of PCa cells. It was found that the E-cadherin level was downregulated by SNHG3 overexpression and upregulated by SNHG3 silencing, and the levels of N-cadherin and Snail were upregulated by SNHG3 overexpression and downregulated by SNHG3 silencing.
The above results indicate that SNHG3 can promote PCa migration, invasion, and EMT.
To reveal the mechanism of SNHG3 in PCa progression, bioinformatics analysis was conducted using starBase, and it was predicted that miR-487a-3p was a competitive binding microRNA. In previous study, miR-487a-3p expression levels had been found to be downregulated in PCa tissue and cells, 14 and related to many biological functions, including cancer cell metastasis and proliferation. 14,15 Through luciferase assay, it was proven that miR-487a-3p could bind to SNHG3 in both LNCaP cells and PC-3 cells. LncRNA exerts its function partly by binding with endogenous competitive RNA. 23 –27
To testify this assumption, they studied the EMT and invasion ability of PCa cells by cotransfection of SNHG3 and miR-487a-3p mimic or miR-487a-3p inhibitor. The authors found that miR-487a-3p mimic inversed the promoting effect of SNHG3 on LNCaP cells and miR-487a-3p inhibitor inversed the inhibitory effect of SNHG3 on PC-3 cells. Similarly, Wang et al. also found that miR-487a-3p can suppress PCa. 14 Taken together with previous studies, these results demonstrated that lncRNA SNHG3 may perform its function through miR-487a-3p.
TRIM25 was also previously reported to have increased expression in PCa tissues 16,17 or other cancer tissue. 28 –30 The data in the research showed that TRIM25 may be the target gene of miR-487a-3p. It was further found that TRIM25 plasmid inversed the inhibitory effect of miR-487a-3p mimic on TRIM25 level in LNCaP cells and TRIM25 siRNA inversed the promoting effect of miR-487a-3p inhibitor on TRIM25 level in PC-3 cells. All these results imply that miR-487a-3p promotes the level of TRIM25 in PCa cells.
Furthermore, they tested the EMT and invasion ability of PCa cells by cotransfection of TRIM25 plasmid or TRIM25 siRNA and miR-487a-3p mimic or miR-487-3p inhibitor. The results showed that TRIM25 plasmid inversed the inhibitory effect of miR-487a-3p mimic on the EMT and invasion ability of LNCaP cells and TRIM25 siRNA inversed the promoting effect of miR-487a-3p inhibitor on the EMT and invasion ability of PC-3 cells. These results imply that miR-487a-3p inhibits PCa cell metastasis through TRIM25. Similar to the discoveries in Wang's study on PCa 16 and Walsh's study on breast cancer, 31 the authors' results also implied that TRIM25 could bind to miR-487a-3p to mediate the function of SNHG3 in promoting the proliferation and metastasis of PCa.
Conclusions
In summary, this research proved that SNHG3 level was upregulated in PCa cells and tissues. Besides, a high SNHG3 level was related with cell proliferation and metastasis, and SNHG3 promotes PCa cell proliferation and metastasis by sponging miR-487a-3p to regulate TRIM25. The findings suggest that SNHG3 may be a new potential diagnostic and therapeutic target for PCa.
Footnotes
Authorship Confirmation Statement
L.Y. designed the research study. L.Y. and Y.R. performed the research and analyzed the data. L.Y. wrote the article. All authors contributed to editorial changes in the article. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Project of Medical and Health Technology Platform of Zhejiang Province [Grant Number 2017ZD028].