
Abstract
Background:
Colorectal cancer (CRC) is a common malignancy of digestive tract. Pinocembrin (PINO) has been discovered to have a proapoptotic effect on CRC. This study aimed to elucidate how other biological behaviors of CRC cells were affected under PINO treatment.
Materials and Methods:
The effect of PINO on HT29 and HCT116 cells were detected through treatment of different concentrations of PINO. The role of LACTB in PINO treatment was investigated by transfection of siRNA-LACTB. Cell counting kit-8 assay, wound healing assay, and Transwell assay were conducted to evaluate the proliferation, migration, and invasiveness of CRC cells, respectively. Western blot or quantitative reverse transcription–polymerase chain reaction was carried out to measure the expressions of LACTB, matrix metalloproteinase (MMP)-2, E-cadherin, and N-cadherin.
Results:
Gradient PINO inhibited the viability, migration, invasiveness, and expressions of MMP-2 and N-cadherin in CRC cells, while promoted E-cadherin and LACTB expressions. Silencing LACTB promoted the viability, migration, invasiveness, and expressions of MMP-2 and N-cadherin in CRC cells and inhibited E-cadherin expression. PINO counteracted the effect of silenced LACTB, and yet silencing LACTB partially abolished the effect of PINO on CRC cells.
Conclusion:
PINO inhibited the proliferation, migration, invasiveness, and epithelial-to-mesenchymal transition of CRC cells by regulating LACTB.
Introduction
Colorectal cancer (CRC) has ranked fourth in cancer-related mortality worldwide, following lung, liver, and stomach cancer. 1,2 An extremely high incidence of CRC has been detected in developed countries. 1 Meanwhile, with the increasing trends of global modernization, people have become more predisposed to adopt Western foods and lifestyle, which contributes to the increment of the incidence of CRC. 1,3 Thus, CRC is regarded as a disease of modernity. 1 It is recorded that around 50% of CRC patients will undergo metastases, with a 5-year survival rate of 50%–60%. 4 The reason for the low survival rate of CRC may be attributable to that the diagnosis of CRC generally occurs at the late stage and the late-stage diagnosis is fundamentally caused by quick emergence of metastasis and high-rate bloodstream transfer. 4 Hence, targeting metastasis in cancer development has a huge significance for achieving an effective treatment for CRC.
Although multifarious properties and mechanisms matter to the emergence of metastasis, the reduction of cell-cell adhesion phenotype in the early formed tumors is the key and primary factor to metastasis. 5 Besides, increased cell mobility and the degradation of the extracellular matrix also stand out subsequently to initiate metastasis. 6 Accumulating researches have reported that epithelial-mesenchymal transition (EMT) plays a pivotal role in cancer infiltration and metastasis. 7 During EMT, cells lose the properties of polarity and adhesion, while acquiring the traits of mobility and mesenchymal phenotype. 8 In many types of carcinomas, the transition to mesenchymal phenotype can accomplish many of the steps required in invasion-metastasis cascade, such as neoplastic cell invasion on the primary tumor site and their intravasation into the circulations and extravasation to the distant area. 9 Previous studies showed that higher degree of transition to mesenchymal phonotype is positively associated to a more unfavorable prognosis of CRC. 10 Therefore, exploration on the mechanism of EMT in metastasis is likely to benefit containment of the development of CRC.
LACTB is a mitochondrial protein that is related evolutionarily to bacterial penicillin-binding/B-lactamase proteins, are ubiquitous in several human tissues, especially in skeletal, heart, muscle, and liver. 11 –13 LACTB has been reported as a tumor suppressor that modulates lipid metabolism and cell state. 14 In addition, studies have found that LACTB is downregulated in many cancers, such as breast, 15 glioma, 16 and liver cancer. 17 Low expression of LACTB promotes tumor progression and predicts poor prognosis in hepatocellular carcinoma 17 ; overexpression of LACTB could suppress the proliferation, invasion, and angiogenesis of glioma cells. 16 In addition, LACTB plays an important regulatory role in the development of CRC. LACTB inhibits CRC progression by attenuating MDM2-mediated p53 ubiquitination and degradation 18 ; and LACTB regulates PIK3R3 to promote autophagy and inhibit EMT and proliferation through the PI3K/AKT/mTOR signaling pathway in CRC. 19
Pinocembrin (PINO), a natural flavonoid, first isolated from honey, with high content in propolis, has a series of incredible pharmacological effects, including antimicrobial, antimutagen, antioxidation, and anti-inflammation. 20 –24 Recent studies revealed that the phytochemicals from PINO and its related derivatives are implicated in cytotoxicity on HCT 116 CRC cells by inducing apoptosis. 25 The inhibitory effect of PINO on the proliferation and migration of ovarian cancer cells was discovered to be exerted by lowering the mRNA levels of N-cadherin and GABAB receptor. 26 These findings suggest an antitumor effect of PINO, while the performance of PINO in the migration, invasion, and EMT in CRC remains unclear. In this study, the authors sought to uncover the underlying molecular mechanism of the effect of PINO on the biological behaviors of CRC cells.
Materials and Methods
Cell culture
CRC cell lines (HT29 and HCT116 cells) were procured from ATCC (HTB-38 and CCL-247, Manassas, VA). HT29 and HCT116 cells were cultured in modified McCoy's 5a Medium (30-2007; ATCC) supplemented with 10% fetal bovine serum (FBS, 16140071; ThermoFisher, Walthma, MA). Both cells were grown at 37°C under 5% CO2.
PINO treatment
PINO (C15H12O4, purity ≥98.0%, PHL80061; Sigma-Aldrich, St. Louis, MO) was dissolved in distilled water supplemented with 20% hydroxypropyl-β-cyclodextrin (H107; Sigma-Aldrich) to prepare stock solution. PINO stock solution was diluted at concentrations of 50, 100, and 200 μM. The diluted PINO solution was added into the culture media of HT29 and HCT116 and incubated at 37°C under 5% CO2 for 48 h. HT29 or HCT116 cells were divided into four groups: control (untreated cells), PINO-50 (cells treated with 50 μM PINO), PINO-100 (cells treated with 100 μM PINO), and PINO-200 (cells treated with 200 μM PINO).
Transfection
Before treatment with PINO for 48 h, HT29 and HCT116 cells underwent transfection of siLACTB (5′-CAUAGUAAGAGAUUAGCAACA-3′, siG15130150426-1-5, RIBOBIO, Guangzhou, China) using Lipofectamine 3000 Transfection Reagent (L3000015; ThermoFisher). Two × 104 of HT29 or HCT116 cells were seeded into each well of 96-well plates (260836; ThermoFisher) to reach a cell confluence of 60%. siLACTB and siNC (5′-GUAACACGAUAAUGUAGAACA-3′; QIAGEN, Hilden, Germany), as well as Lipofectamine 3000 reagent were all diluted using Opti-MEM media (31985088; ThermoFisher). siLACTB solution was (1:1) added with the diluted Lipofectamine 3000 reagent, followed by incubation for 10 min at room temperature. The incubated solution was added into 10 μL lipid complex supplemented with 0.2 μL P3000 reagent and again incubated for 2 d at 37°C.
Cell counting kit-8 assay
Each well of 96-well plates was filled with 100 μL solution of HT29 or HCT116 cells with or without transfection of siLACTB. After PINO treatment at indicated concentrations for 48 h, HT29 or HCT116 cells were added with 10 μL Cell counting kit (CCK)-8 reagent (CA1210; Solarbio, Beijing, China). Following incubation for 4 h at 37°C, the absorbance of HT29 or HCT116 cells was determined at 450 nm on a microplate reader (ELx808; BioTek, Winooski, VT).
Wound healing assay
Five × 105/well of HT29 or HCT116 cells were seeded into the 24-well plate to form an optically confluent monolayer. Linear gaps were made manually with a sterilized steel rule among the monolayer of HT29 or HCT116 cells, followed by a wash with phosphate buffer saline (PBS, 28348; ThermoFisher) to remove unattached cells. HT29 or HCT116 cells were added with serum-free culture media and incubated with PINO at indicated concentrations for 48 h. After incubation for 48 h, the migratory state of HT29 or HCT116 cells was recorded under a microscope (Axioskop 40, Carl Zeiss AG, Dresden, Germany) and analyzed using an image analysis system (Wound Healing ACAS, ibidi, Germany).
Transwell assay
The invasiveness of HT29 or HCT116 cells with or without transfection of siLACTB was assessed using 24 mm Transwell chambers with 8.0 μm Pore Polycarbonate Membrane Insert (3428; Corning, Corning, NY). The upper chamber was coated with 100 μL diluted Matrigel (BD Biosciences, Franklin Lakes, NJ), infused with 100 μL serum-free culture media, and incubated for 5 h at 37°C. HT29 or HCT116 cells were digested adjusting to 2 × 105/mL and added into the upper chamber. The bottom chamber was added with 600 μL culture media containing 10% FBS to induce cell invasion. After incubation with PINO for 48 h, the upper chamber was fixed by 4% paraformaldehyde (P6148; Sigma-Aldrich) for 30 min at room temperature and stained with 0.1% crystal violet at room temperature for 30 min. The invading cells were counted under a microscope (Axioskop 40; Carl Zeiss AG, Dresden, Germany).
Western blot
PINO-treated HT29 or HCT116 cells were harvested and lysed using RIPA Buffer (89900; ThermoFisher). Total protein was obtained by centrifugation of the cell lysate at 2000 g for 10 min and quantitated using a BCA kit (A53227; ThermoFisher). Proteins were separated using 12% SDS-PAGE gel (P0053A; Beyotime, Shanghai, China) for 1 h. The separated proteins were then transferred onto polyvinylidene difluoride (PVDF) membranes (P2438; Sigma-Aldrich). After being blocked by 5% skimmed milk in Tris buffered saline with 1% Tween 20 (TBST, TA-125-TT; ThermoFisher) for 1 h at room temperature, the membranes were incubated overnight with the following primary antibodies at 4°C: MMP-2 (#40994, 72 kDa, 1:1000; Cell Signaling, Danvers, MA), E-cadherin (ab1416, 110 kDa, 1:50; Abcam, Cambridge, MA), N-cadherin (ab18203, 100 kDa, 1:1000; Abcam), and GAPDH (ab181602, 36 kDa, 1:10,000; Abcam). The membranes were then washed with TBST and incubated with goat anti-mouse IgG H&L (HRP) (ab205719, 1:2000; Abcam) or goat anti-rabbit IgG H&L (HRP) (ab205718, 1:2000; Abcam) for 2 h at room temperature. The visualization of the proteins was carried out using enhanced chemiluminescence reagent kit (WP20005; ThermoFisher) on an imaging System (iBright CL1500A; ThermoFisher). Protein bands were analyzed by densitometry using ImageJ software (1.52s version; National Institutes of Health, Bethesda, MA).
Quantitative reverse transcription–polymerase chain reaction
Total mRNA was isolated by TRIzol LS Reagent (10296010; ThermoFisher) from HT29 or HCT116 cells. SuperScript IV reverse transcriptase (18090010; ThermoFisher) was used to synthesize the cDNA of the isolated mRNA. cDNA was added with PowerUp SYBR Green Master Mix (A25742; ThermoFisher) and primers listed in Table 1 were to undergo real-time quantitative reverse transcription–polymerase chain reaction (qRT-PCR) on a real-time PCR detection system (CFX Connect; Bio-Rad, Philadelphia, PA). PCR parameters were as follows: 95°C predenaturation for 10 min, 95°C denaturation for 10 s, and 40 circles of 55°C annealing for 30 s and 72°C elongation for 30 s. The relative changes in the LACTB, matrix metalloproteinase (MMP)-2, E-cadherin, and N-cadherin expression levels were calculated by 2−ΔΔCt method. 27
Primers Sequence Used in Quantitative Polymerase Chain Reaction
Statistical analysis
All data were represented as mean ± SD. All experiments were performed in triplicate. Statistical analyses were performed using SPSS software (version 13.0; SPSS, Inc., Chicago, IL). Statistical significance was determined by one-way ANOVA with a successive Dunnett's post hoc test. p < 0.05 was expected to be significant.
Results
PINO inhibited CRC cell viability, migration, and invasion
Previous study demonstrated a marked cytotoxicity induced in CRC cells under exposure of PINO at 50, 100, and 200 μM for 48 h. 28 Gao's in vitro preclinical study showed that a 48-h treatment of PINO at 100 and 200 μM has an inhibitory effect on ovarian cancer cell viability and migration. 29 Thus, based on these pretested PINO concentrations, the authors launched investigations on the mechanism of how PINO affects the biological behaviors of CRC cells. Treatment with increasing concentrations of PINO (50, 100, and 200 μM) on CRC HT29 and HCT116 cells decreased the viabilities of HT29 and HCT116 cells (p < 0.05, Fig. 1A). Accordingly, they further conducted experiments on the effect of PINO on HT29 and HCT116 cell migration and invasion. Likewise, PINO treatment at 50, 100, and 200 μM resulted in increasingly reduced migratory distance and invading range of HT29 and HCT116 cells (p < 0.01, Fig. 1B–E). The above results suggested an antagonizing effect of PINO on CRC cell progression.

PINO inhibited CRC cell viability, migration, and invasion.
PINO inhibited EMT and promoted LACTB expression in CRC cells
EMT is widely recognized to closely associate the development of cancer cell migration and invasion. 30 Meanwhile, LACTB is discovered to act as a tumor suppressor by modulating mitochondrial lipid metabolism. 14 In addition, increased expression of MMP-2 is found to facilitate EMT to propel cancer development. 31 Hence, the expressions of E-cadherin, N-cadherin, MMP-2, and LACTB in response to PINO treatment were analyzed. PINO treatment at 50, 100, and 200 μM significantly inhibited the protein and mRNA expressions of MMP-2 and N-cadherin (p < 0.05), while promoting E-cadherin protein and mRNA expressions in HT29 and HCT116 cells (p < 0.01, Fig. 2A–C). LACTB mRNA exhibited a growth in its expression after PINO treatment at 50, 100, and 200 μM in HT29 and HCT116 cells (p < 0.001, Fig. 2D). These findings suggested that the antagonizing effect of PINO was also valid on EMT and might relate to the promotion of LACTB expression.
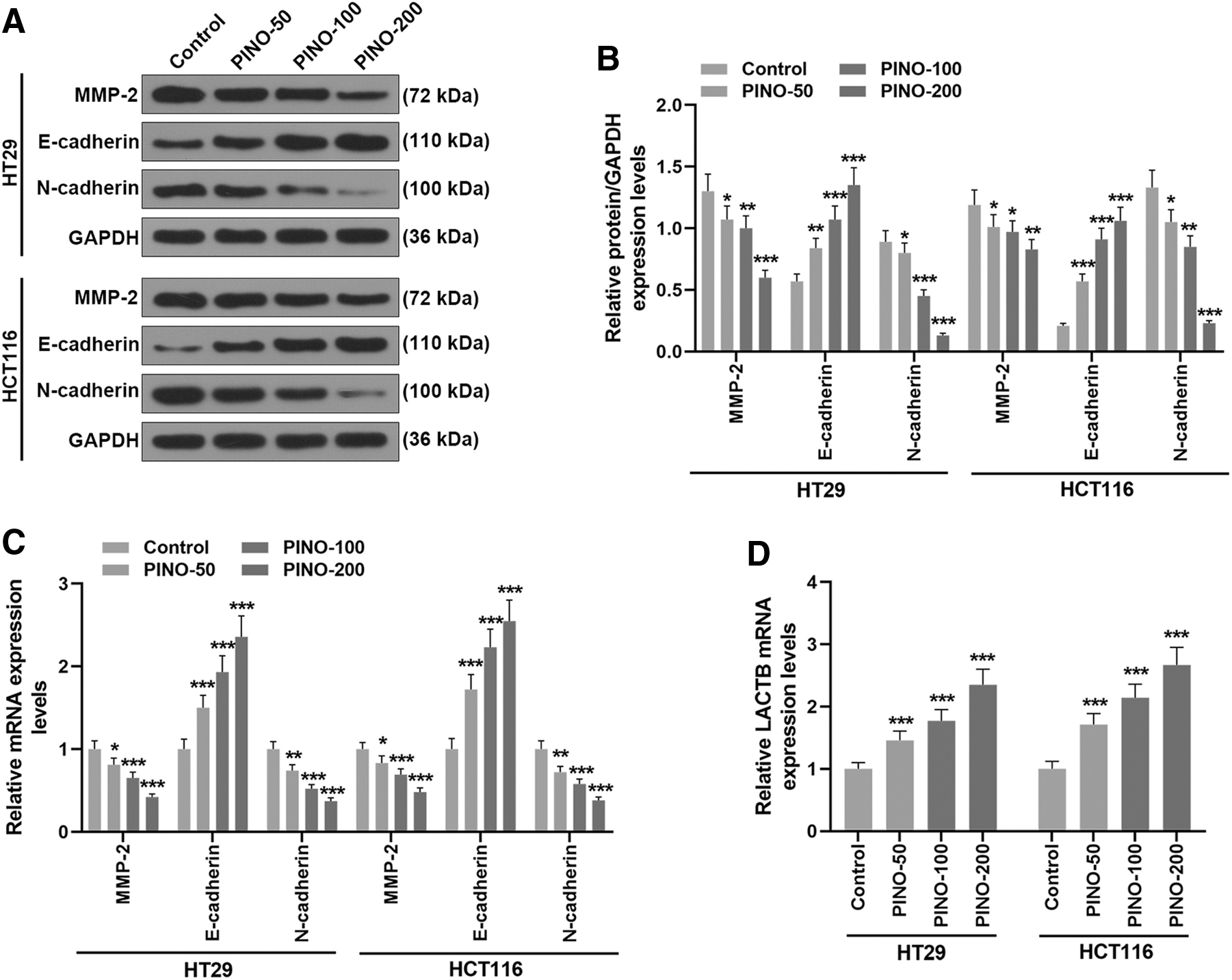
PINO inhibited EMT and promoted LACTB expression in CRC cells.
PINO counteracted the effect of siLACTB on the LACTB expression and viability of CRC cells
To verify the importance of LACTB in the CRC cell progression-antagonizing effect of PINO, siLACTB was used to control LACTB expression in PINO (100 μM)-treated HT29 and HCT116 cells. Transfection of siLACTB, which resulted in an inhibited LACTB mRNA expression (p < 0.001), evidently promoted the viability of HT29 and HCT116 cells (p < 0.001, Fig. 3A, B). More vitally, transfection of siLACTB promoted the viability of PINO (100 μM)-treated CRC cells (p < 0.001, Fig. 3A, B). In turn, PINO treatment promoted LACTB mRNA expression and inhibited the viability of siLACTB-transfected CRC cells (p < 0.001, Fig. 3A, B). Overall, the results showed that PINO treatment counteracted the inhibitory effect of silenced LACTB on LACTB expression and the promotive effect of LACTB on CRC cell viability.
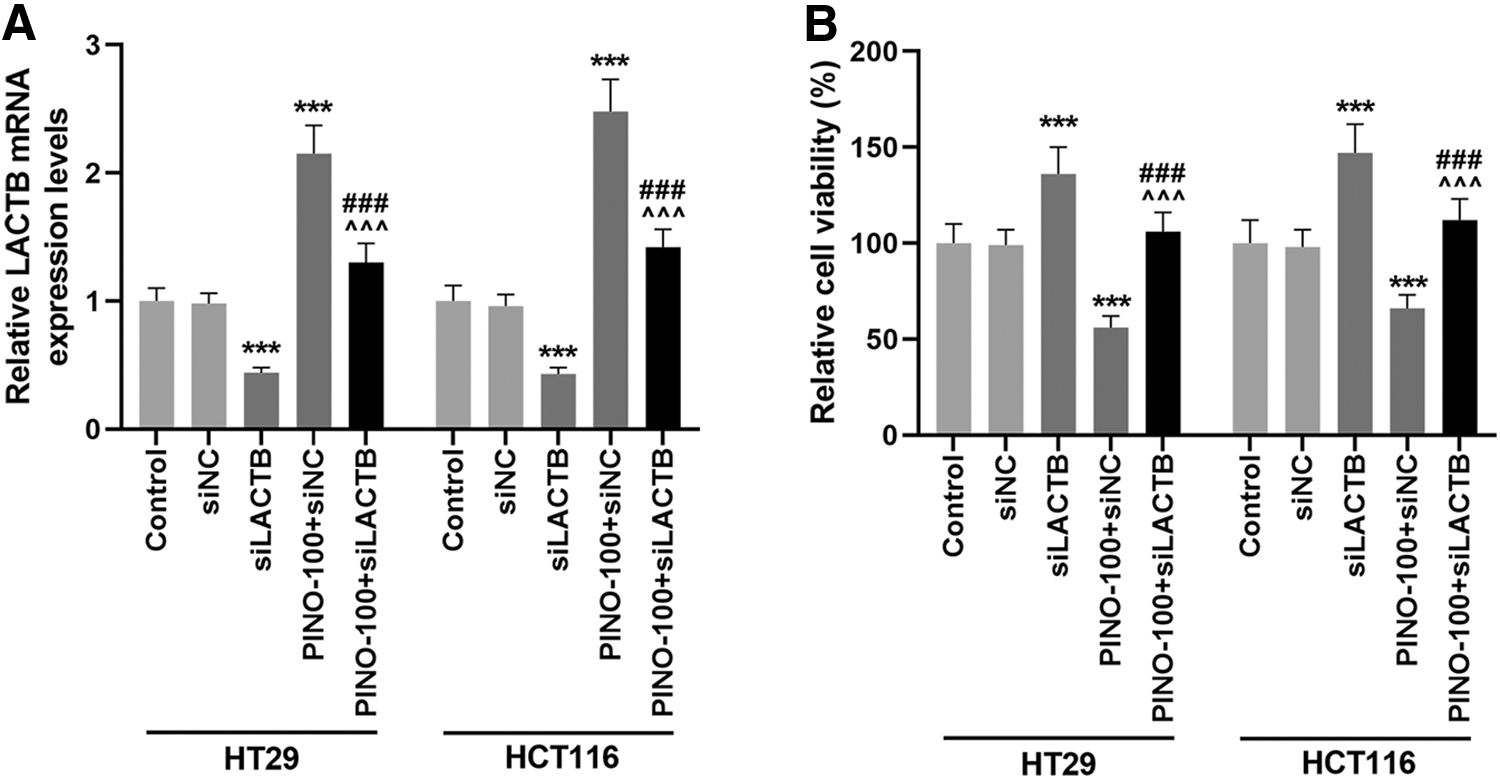
PINO counteracted the effect of siLACTB on the LACTB expression and viability of CRC cells.
PINO counteracted the effect of siLACTB on CRC cell migration and invasion
To further investigate the relationship between LACTB expression and the antagonizing effect of PINO on the migration and invasiveness of CRC cells, the migration and invasiveness of PINO (100 μM)-treated HT29 and HCT116 cells in response to transfection of siLACTB were measured. Transfection of siLACTB displayed longer migratory distance and larger invading range of PINO-treated CRC cells than nontransfected PINO-treated CRC cells (p < 0.01, Fig. 4A–D). Similar to its effect on the viability of PINO-treated CRC cells, transfection of siLACTB extended the migratory distance and invading range of PINO-treated CRC cells (p < 0.001, Fig. 4A–D). As PINO treatment could reverse the effect of transfection of siLACTB on CRC cell viability, it shrank the migratory distance and invading range of siLACTB-transfected CRC cells (p < 0.001, Fig. 4A–D). These findings showed that PINO treatment counteracted the promotive effect of LACTB on CRC cell migration and invasion.
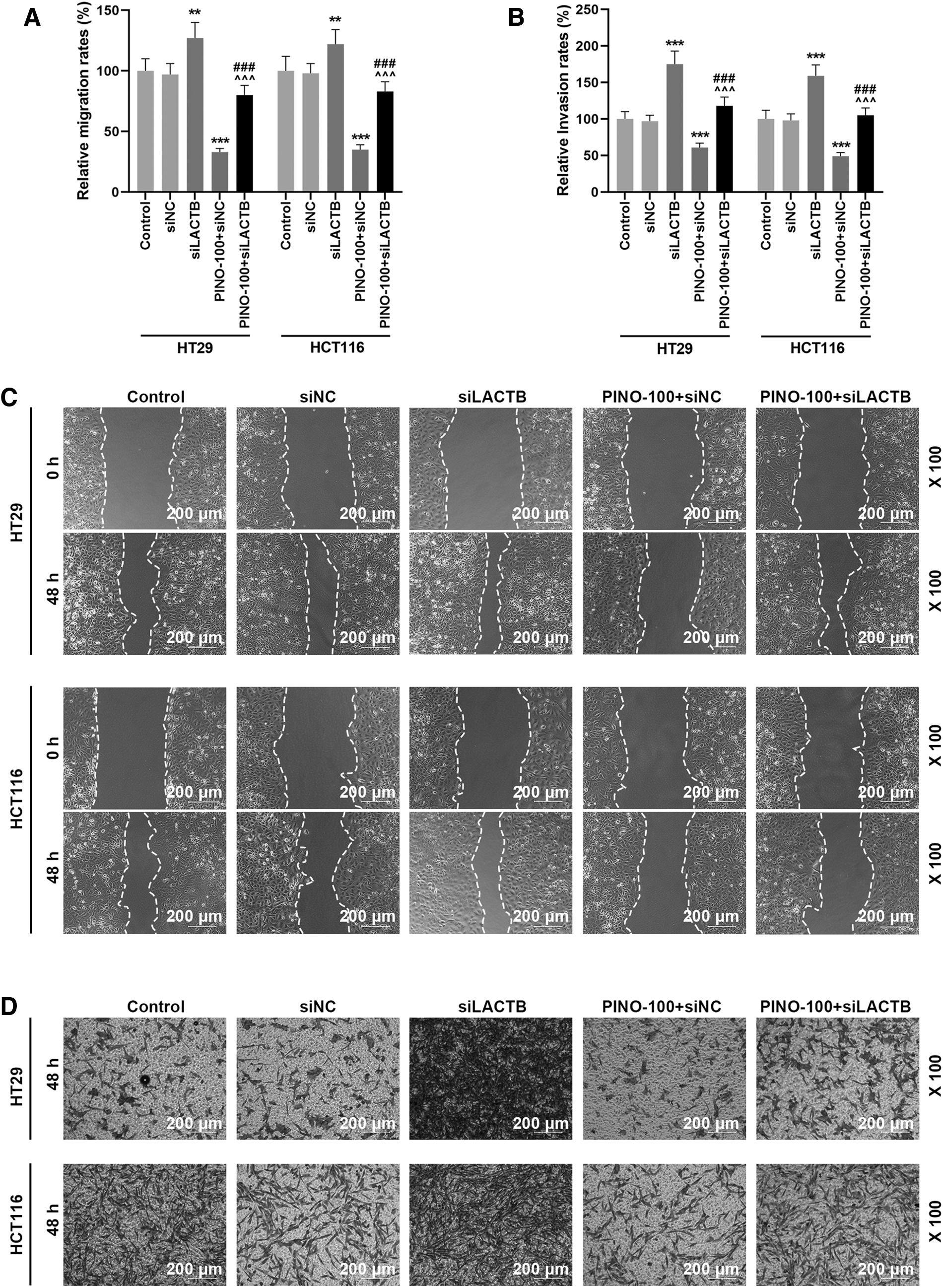
PINO counteracted the effect of siLACTB on CRC cell migration and invasion.
PINO counteracted the effect of siLACTB on EMT
Since silencing LACTB could abolish the inhibitory effect of PINO on the migration and invasiveness of CRC cells and EMT could endow epithelial cells with the ability to migrate and invade, 30 the authors inferred that silencing LACTB exerted the same effect on the PINO-inhibited EMT of HT29 and HCT116 cells and thereby analyzed the expressions of EMT-related factors. Transfection of siLACTB demonstrated pronouncedly promoted the protein and mRNA expressions of MMP-2 and N-cadherin (p < 0.01), but markedly inhibited the protein and mRNA expressions of E-cadherin (p < 0.001, Fig. 5A–D). As was expected, transfection of siLACTB promoted the protein and mRNA expressions of MMP-2 and N-cadherin (p < 0.001), while inhibiting the protein and mRNA expressions of E-cadherin in PINO-treated CRC cells (p < 0.001, Fig. 5A–D). PINO treatment inhibited the protein and mRNA expressions of MMP-2 and N-cadherin (p < 0.01), while prompting the protein and mRNA expressions of E-cadherin in siLACTB-transfected CRC cells (p < 0.01, Fig. 5A–D). The results indicated that PINO treatment counteracted the promotive effect of LACTB on CRC cell EMT.

PINO counteracted the effect of siLACTB on EMT.
Discussion
Tumor recurrence and metastasis are responsible for most of the CRC-related deaths. 32 During metastasis, multiple steps were accomplished, including extracellular matrix remodeling, cytoskeleton reorganization, cell-cell adhesion declining, and cell motility increasing, most of which were favored by EMT. 33 EMT is a pivotal regulatory program that is reported to propel invasion and metastasis of epithelium-derived carcinomas by disrupting cell-cell adherence, reducing cell polarity, and reorganizing cytoskeleton into mesenchymal phenotype. 32 E-cadherin, profusely existing in epithelial cells, and N-cadherin, abundantly expressed in cardiomyocytes and neurons, which function depending on calcium ions, are two cell adhesion molecules and typical markers of EMT. 32 Accumulating evidences have verified that low-expressed E-cadherin is indicative of emergence of lymph node metastasis and a worse prognosis in CRC patients. 33,34 Therefore, focusing on the exploration of an antimetastasis and EMT agent has great potential to provide effective treatments for interfering in the development of CRC.
PINO is derived from and has a prominently high concentration in Elytranthe parasitica (L.) Danser. Elytranthe parasitica (L.) Danser, a parasitic flowering shrub, belongs to Loranthaceae family. 25 Medicinal plants from Loranthaceae family are recorded for use in anticancer therapy. 35,36 Hence, this study surmised that PINO treatment can exert an antagonizing effect in CRC cell proliferation, migration, invasion, and EMT. Aiello's study discovered that the extracts of the byproducts whose main constituent is PINO, from Glycyrrhiza glabra cultivation and harvesting, remarkably lowered the viability of MCF-7, MDA-MB-231, HeLa, PC3, and Caco-2 human cancer cells in a concentration-dependent manner and the efficacy of PINO on some cancer cells was comparable to doxorubicin (a commercial antitumor compound). 29 Kumar's studies displayed dose-dependent antiproliferative, apoptosis-inducing, and cell cycle arrest-inducing properties of PINO in HCT 116 CRC cells. 25 PINO triggers Bax-dependent mitochondrial apoptosis in colon cancer cells. 28 In line with their findings, this study demonstrated PINO at 10, 20, and 50 μM exerted an inhibitory effect, which was gradually strengthening on the viability of CRC cells.
Since metastasis accounts for the worse prognosis of CRC patients and EMT as an indispensable program in metastasis triggers migration and invasion, 37 evaluating the effect of PINO on CRC cell migration, invasion, and EMT can further reflect its anticancer capability. Gao's study revealed the antimigratory effect of PINO on ovarian cell. 26 In Chen's study, the invasion, migration, and EMT of retinoblastoma cells were inhibited by PINO, which was concomitant with N-cadherin expression decreasing. 38 Palaghia's study found a conspicuous membranous expression of E-cadherin in both CRC at primary stage and at lymph node metastatic stage. 39 In addition, the degradation of extracellular matrix caused by cancer cell involves a complex system of proteases such as MMP. 40 A remarkable transcriptional activity of MMP-2 is detected in the front of invasive site built by CRC tumor. 4 Consistent with these findings, PINO in this study could also concentration dependently inhibit invasion and migration, as well as EMT, which was evidenced by upregulated E-cadherin expression and downregulated expressions of N-cadherin and MMP-2 in CRC cells.
LACTB, found in mitochondria, was previously discovered as a breast cancer suppressor, which works through the mechanism of reprogramming mitochondrial lipid metabolism to modulate cancer cell differentiation. 14 LACTB expression induction in breast cancer cells enables the formation of small epithelial islands, accompanied by upregulated epithelial markers and downregulated mesenchymal markers, presenting inhibited tumorigenesis. 14 Recent study has reported a tumor-suppressive effect of LACTB on CRC, and its findings regarding mechanism showed that LACTB could prevent degradation of p53 by binding to the C terminus of p53, thereby suppressing the proliferation, migration, invasion, and EMT of CRC cells. 18 Accordingly, the authors made a hypothesis that the inhibitory effect of PINO on CRC cell progression is ascribed to the expression of LACTB. Indeed, silencing LACTB in PINO-treated CRC cells promoted CRC cell progression. Meanwhile PINO treatment weakened the promotive effect of silenced LACTB on CRC cell progression.
In this study, preliminary results in in vitro experiments for PINO that inhibited the proliferation, migration, invasion, and EMT of HT29 and HCT116 cell were obtained; however, the effect of PINO needs to be further verified using in vivo experiments. In addition, the molecular mechanisms of PION's regulation of CRC, such as signaling pathways, need further study. In follow-up studies, it is necessary to increase the PINO-treated and untreated CRC transcriptome sequencing technology to further explore the potential molecular mechanism of PINO-mediated tumor suppression.
Conclusion
In summary, this study confirmed the antitumor effect of PINO valid in CRC cells, as demonstrated by the fact that PINO inhibited the proliferation, migration, invasion, and EMT of HT29 and HCT116 cells. More importantly, this antitumor effect is positively concentration dependent and may be partially ascribed to the restoration of LACTB expression in CRC cells.
Footnotes
Authors' Contributions
Substantial contributions to conception and design: J.L.; data acquisition, analysis, and interpretation: Y.B., F.Y., Z.F., and L.Y.; drafting the article or critically revising it for important intellectual content: J.L.; final approval of the version to be published: all authors; and agreement to be accountable for all aspects of work in ensuring that questions related to the accuracy or integrity of the work are appropriately investigated and resolved: all authors
Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
No funding was received for this article.