
Abstract
Background:
Triple-negative breast cancer (TNBC) is the most serious subtype of breast cancer (BC) and has been a great health threat to females. Although chemotherapeutic agent contributes a lot to TNBC treatment, drug resistance has been a great obstacle for chemotherapies. Ursolic acid (UA), a pentacyclic triterpenoid compound, was reported to reverse paclitaxel resistance in BC. However, whether UA could affect the resistance of TNBC cells to other drugs such as doxorubicin (DOX) remains to be discovered.
Materials and Methods:
MTT assay, EdU assay, colony formation assay, and flow cytometry analysis were implemented to detect the viability, proliferation, and apoptosis of DOX-resistant MDA-MB-468 and MDA-MB-436 cells with or without UA treatment. Mechanism assays including RIP, RNA pull-down, and luciferase reporter assays verified the interaction between RNAs.
Results:
UA treatment hindered the growth and mitigated the DOX resistance of DOX-resistant MDA-MB-468 and MDA-MB-436 cells. Mechanistically, multidrug resistance-associated protein 1 (ABCC1) expression was downregulated by UA treatment. MiR-186-5p was verified to target ABCC1. Further, UA-inhibited ZEB1-AS1 (zinc finger E-box binding homeobox 1 antisense RNA 1) was verified as a competitive endogenous RNA (ceRNA) to upregulate ABCC1 through sponging miR-186-5p. Importantly, UA treatment impaired the malignant phenotypes of DOX-resistant MDA-MB-468 and MDA-MB-436 cells through ZEB1-AS1/ABCC1 axis.
Conclusion:
UA promotes TNBC cell sensitivity to DOX through inactivating ZEB1-AS1/miR-186-5p/ABCC1 signaling.
Introduction
As a common female cancer, breast cancer (BC) accounts for ∼6.6% of the global cancer fatality rate. 1 Triple-negative breast cancer (TNBC) is a BC subtype that accounts for ∼20% of all BC cases. 2 Due to higher recurrence and poorer outcomes, TNBC is more aggressive than other subtypes of BC. 3 Doxorubicin (DOX) is currently deemed as one of the most effective drugs for TNBC treatment. 4 Disappointingly, as DOX treatment is prolonged, drug resistance builds up, ultimately leading to the failure of cancer treatment. 5,6 Yet, the mechanism of DOX resistance remains poorly understood. Hence, it is in urgent need to strengthen the research on the underlying mechanism of DOX resistance in TNBC.
Ursolic acid (UA) is a pentacyclic triterpenoid identified in medical herbs such as loquat leaf and rosemary. Previous studies have discovered that UA could inhibit the development of lung cancer 7 and gastric cancer. 8 In addition, a recent report revealed that UA reverses the paclitaxel resistance of BC cells by miR-149-5p/MyD88 axis. 9 This study was designed to explore the role and underlying mechanism of UA in the DOX resistance of TNBC cells.
Recently, molecular research has put the emphasis on long noncoding RNAs (lncRNAs). LncRNAs possess a length of >200 nucleotides and are incapable of encoding proteins. It has been widely reported that lncRNAs play pivotal roles in the building of drug resistance in various cancers. As an example, depletion of lncRNA-HOTAIR alleviates DOX resistance of BC cells through inducing inactivation of PI3K/AKT signaling. 10
It was also reported that lncRNAs regulate drug resistance through competitive endogenous RNA (ceRNA) patterns. In such mechanisms, lncRNAs serve as a ceRNA to endogenously sponge microRNAs (miRNAs), thereby blocking miRNAs-mediated mRNA suppression. For instance, Wang et al. revealed that downregulation of lncRNA GAS5 promotes drug resistance in liver cancer by inhibiting PTEN expression through interacting with miR-21. 11 Upregulated PDIA3P1 expression confers chemoresistance through serving as an endogenous sponge of miR-125a/b/miR-124 to increase TRAF6 expression. 12 LncRNA AFAP1-AS1 facilitates cell proliferation and drug resistance in nonsmall cell lung cancer through miR-139-5p/RRM2 axis. 13 However, the role of lncRNAs in the DOX resistance of TNBC cells remains to be uncovered.
ATP-binding cassette transporter C (ABCC) family includes 12 members, among which 9 (ABCC1-ABCC6 and ABCC10-ABCC12) are multidrug resistance proteins (MRPs; MRP1-9) that are known as drug transporters mediating drug efflux in cancer cells. 14 As a result, this drug efflux impairs the accumulation of drugs within cancer cells, making them multiresistant to diverse drugs. 15,16 The status of ABCC1 (MRP1) has been previously suggested to predict treatment response and eventual clinical outcomes in TNBC. 17 Further, targeting ABCC1 has been exploited to overcome chemoresistance in TNBC. 18 However, the upstream regulatory mechanisms of ABCC1 in TNBC are largely unexplored.
MicroRNAs (miRNAs) are introduced as the regulators of protein-coding genes through direct silencing messenger RNAs (mRNAs) through RNA-induced silencing complexes (RISCs). 19 Interestingly, lncRNAs have been proposed to destroy the suppression of miRNAs on mRNAs through a competing endogenous RNA network over decades. 20 Nonetheless, the existence of certain ceRNA network influenced by UA in TNBC cells needs to be proved.
In this study, we aimed to determine the impact of UA on the DOX resistance of TNBC cells. Further, we sought to explore the underlying ceRNA mechanism of how UA affected DOX resistance in TNBC, which might enrich the academic knowledge on DOX resistance in TNBC treatment.
Materials and Methods
Cell culture and treatment
The human TNBC cell lines (MDA-MB-468 and MDA-MB-436) were available from Shanghai Institute of Biochemistry and Cell Biology (Shanghai, China). Cells were cultured in RPMI-1640 medium (A1049101; Thermo Fisher Scientific, Inc., Waltham, MA) with 10% FBS and 1% antibiotics as medium supplements. Cell culture was maintained in 5% CO2 at 37°C. UA was purchased from Shanghai Yuanye Biological Technology (77-52-1; Shanghai, China) and dissolved in DMSO (D2650; Sigma-Aldrich, St. Louis, MO). For UA treatment, the cells were subjected to RPMI-1640 medium supplemented with 20, 40, 80, 160, 320, and 640 mg/mL UA. Besides, in the following experiments, cells treated with 40 mg/mL UA were defined as UA group, while those without UA treatment were defined as control group.
Establishment of DOX-resistant TNBC cells
The parental MDA-MB-468 and MDA-MB-436 cells were first cultured in normal culture medium until cells were in the logarithmic growth phase. Then, cells were successively processed with RPMI-1640 medium containing 20, 40, 60, 80, and 100 μg/mL of DOX (D1515; Sigma-Aldrich), with each dose cultured for ∼2 weeks. Culture medium was changed every 3 d. After that, the DOX-resistant cells (termed DOX-MDA-MB-468 and DOX-MDA-MB-436) were thought to be successfully established once they could survive well in the medium with 80 μg/mL of DOX. Before using, the DOX-resistant cells maintained in the medium with 80 μg/mL of DOX were grown in the drug-free RPMI-1640 medium for 3 d.
Total RNA isolation and qRT-PCR
The isolation of total RNA was achieved using TRIzol reagent (15596018; Thermo Fisher Scientific), followed by cDNA synthesis using PrimeScript Reverse Transcriptase Kit (2690A; Takara, Shiga, Japan). Quantitative PCR analyses were conducted with SYBR Green PCR Kit (Takara) through StepOnePlus™ Real-time PCR Systems (Applied Biosystems, Foster City, CA). PCR cycling conditions for all genes were provided as below: 3 min of 95°C for 1 cycle; 30 s of 95°C, 30 s of 60°C, and 1 kb/min of 72°C, for 35 cycles; 10 min of 72°C for 1 cycle. The relative gene expression was processed based on 2−ΔΔCt method. 21 The U6 snRNA or GAPDH mRNA served as the internal control. All primers used in this experiment are provided in Supplementary Table S1.
Cell transfection
The shRNAs designed for ABCC1 or ZEB1-AS1, as well as negative control shRNA (sh-NC), were all acquired from GenePharma (Shanghai, China). The pcDNA3.1 vectors (Invitrogen) were procured to construct pcDNA3.1-ZEB1-AS1 and pcDNA3.1-ABCC1, while the empty vector was the negative control. In addition, miRNA mimics and miR-186-5p inhibitor, as well as NC mimics/inhibitor, were synthesized by Ribobio (Guangzhou, China). Cells were transfected for 48 h using Lipofectamine 2000 (11668019; Invitrogen, Carlsbad, CA).
Cell viability assay
The viability of DOX-MDA-MB-468 and DOX-MDA-MB-436 with UA treatment or not was assessed using MTT assay. About 1 × 103 cells were seeded into each well of 96-well plates and cultured until 90% confluence, followed by the incubation of 20 μL of 0.5 mg/mL MTT (M2128; Sigma-Aldrich) for 4 h. Then, 150 μL of DMSO was added to each well for 10 min of oscillation. In the end, the absorbance at 560 nm was detected with microplate reader (51119080; Thermo Fisher Scientific).
EdU assay
EdU assay was conducted in DOX-MDA-MB-468 and DOX-MDA-MB-436 cells with or without UA treatment using BeyoClick™ EdU Cell Proliferation Kit (ST067; Beyotime, Shanghai, China) with Alexa Fluor 594. Cells were stained by EdU medium and DAPI solution, and finally imaged under a fluorescent microscope (DMI8; Leica, Wetzlar, Germany).
Colony formation assay
The DOX-MDA-MB-468 and DOX-MDA-MB-436 cells, with or without UA treatment, were prepared in six-well plates with 800 cells seeded into each well. After 14 d, cells were fixed by 4% paraformaldehyde (PFA; E672002; Sangon Biotech, Shanghai, China), stained by 0.5% crystal violet (V5265; Sigma-Aldrich), and then counted manually.
Flow cytometry analysis
The apoptosis of DOX-MDA-MB-468 and DOX-MDA-MB-436 cells with UA treatment or not was examined using flow cytometry as required by provider. The double Annexin V/PI staining kit from BD Biosciences (559763; San Jose, CA) was used. At length, apoptotic rate of cell lines was assessed by a Cytoflex flow cytometer (Beckman Coulter, Inc.; 250).
Caspase-3/8/9 activity detection
The caspase-3/8/9 activities were separately examined using the Caspase-3/8/9 activity assay kits (C1115/C1152/C1157; Beyotime), per the guidebooks. Cell protein lysates were mixed with the indicated assay buffer and caspase substrate. Finally, to detect the caspase activities, the absorbance at 405 nm was measured by a microplate reader (51119080; Thermo Fisher Scientific).
Luciferase reporter assay
The ABCC1 (RefSeq number: NM_004996) or ZEB1-AS1 (RefSeq number: NR_024284) fragments covering miR-186-5p binding sites (wild type and mutant) were used to construct ABCC1-WT/Mut and ZEB1-AS1-WT/Mut vectors using pmirGLO vectors (Promega, Madison, WI). The acquired vectors were cotransfected with miR-186-5p (RefSeq number: NR_029707) mimics or NC mimics into DOX-MDA-MB-468 and DOX-MDA-MB-436 cells with or without UA treatment. After 48 h of cotransfection, the luciferase intensity was examined using luciferase reporter assay system (E1910; Promega).
RNA pull-down assay
RNA pull-down assay was performed using Pierce Magnetic RNA-Protein Pull-Down Kit (20164; Thermo Fisher Scientific). Cell lysates were mixed with biotinylated miR-186-5p probes (5'-CAAAGAAUUCUCCUUUUGGGCU-3′), while those with the no biotin-labeled probes served as control groups. One night later, the mixtures were added with streptavidin-linked magnetic beads for 4 h of incubation. RNAs in the pulled-down mixtures were assayed finally using qRT-PCR.
Fluorescence in situ hybridization
Fluorescence in situ hybridization (FISH) experiment was carried out according to the recommended protocol of Ribo™ Fluorescent in Situ Hybridization Kit (C10910; Ribobio, Guangzhou, China). Simply put, the DOX-MDA-MB-468 and DOX-MDA-MB-436 cells were digested and air-dried after fixation, followed by hybridization with ZEB1-AS1-FISH probe (Ribobio). Cell nuclei were probed with DAPI solution. All samples were finally analyzed using a fluorescence microscope (DMI8; Leica, Wetzlar, Germany). The sequence of ZEB1-AS1 probe is shown in Supplementary File S1.
Subcellular fractionation
PARIS™ Kit (Ambion, Austin, TX) was acquired, and used for subcellular fractionation of DOX-MDA-MB-468 and DOX-MDA-MB-436 cells. After lysing in cell fractionation buffer, cell cytoplasm and cell nucleus were obtained. Finally, the ZEB1-AS1 content in both cell fractions was quantified by qRT-PCR. GAPDH was applied as the cytoplasmic control with U6 as the nuclear control.
RNA immunoprecipitation
Magna RIP™ RNA-Binding Protein Immunoprecipitation Kit was purchased from Millipore (17-700; Millipore, Billerica, MA) for RNA immunoprecipitation (RIP) assay. Anti-Ago2 antibody (1:50, #2897; Cell Signaling Technology, Boston, MA) and IgG antibody (control group; 1:50, #3900; Cell Signaling Technology) were used in this assay. The lysates of DOX-MDA-MB-468 and DOX-MDA-MB-436 cells were collected after treatment with or without UA. Then, RIP buffer with antibody-bound magnetic beads was added to the lysates for overnight incubation. After that, the precipitated RNAs were subjected to qRT-PCR analysis.
Statistical analyses
Data were presented as the means + SD of three separately conducted experiments. All statistical analyses in this study were accomplished using Student's t-test or one-way ANOVA, with the application of GraphPad Prism 7 (La Jolla, CA). The differences were significant when p-values were <0.05.
Results
UA decreased the DOX resistance and hindered the growth of DOX-resistant TNBC cells
First, the DOX-resistant TNBC cells (termed DOX-MDA-MB-468 and DOX-MDA-MB-436) were established. The IC50 values of DOX in parent cells and DOX-resistant cells are demonstrated in Figure 1A. Then, we conducted functional assays to explore the effects of UA on the DOX-resistant TNBC cells. It was revealed from the data of MTT assay that UA decreased cell viability in DOX-resistant cells in a concentration-dependent manner (Fig. 1B). Of note, after treatment with UA (40 mg/mL), the IC50 value of DOX in DOX-MDA-MB-468 cells declined to 18 μg/mL and that in DOX-MDA-MB-436 cells declined to 22 μg/mL (Fig. 1C).

UA hampered the growth and reversed the DOX resistance of DOX-resistant TNBC cells.
Then, EdU and colony formation assays were implemented to explore changes in cell proliferation in face of UA treatment. The results showed that UA treatment significantly reduced the proliferation ability of DOX-resistant cells (Fig. 1D, E). Further, we conducted flow cytometry analysis and detected caspase-3/8/9 protein activity. The results revealed that UA treatment efficiently facilitated the apoptosis of DOX-MDA-MB-468 and DOX-MDA-MB-436 cells (Fig. 1F, G). More importantly, we discovered that although DOX treatment had some suppression on the growth of the two resistant TNBC cells, a stronger growth-inhibiting phenomenon was observed after cotreating with UA (Supplementary Fig. S1), which indicated that the two DOX-resistant TNBC cells treated with UA were more sensitive to DOX treatment.
In brief, UA reduced cell proliferation and increased cell apoptosis in DOX-resistant TNBC cells.
UA reversed DOX resistance through downregulation of ABCC1
Since the members of ABCC family are known to mediate drug resistance, we detected the expression of ABCC1, ABCC2, ABCC3, ABCC4, and ABCC5 in DOX-resistant cells with or without UA treatment. It was revealed that only ABCC1 was downregulated by UA treatment (Fig. 2A).

UA restrained the growth of DOX-resistant TNBC cells through downregulation of ABCC1.
Then, we conducted loss-of-function assays in DOX-resistant cells with or without UA treatment. Before that, the depletion efficiency of ABCC1 was verified through qRT-PCR analysis (Supplementary Fig. S2A). It was revealed by EdU and colony formation assays that depletion of ABCC1 restrained the proliferation of DOX-MDA-MB-468 and DOX-MDA-MB-436 cells, and also strengthened the suppression of UA treatment on the proliferation of both cells (Fig. 2B, C). Next, the results of flow cytometry analysis demonstrated that in the two DOX-resistant TNBC cells, silencing ABCC1 enhanced cell apoptosis and meanwhile further reinforced the facilitating effect of UA treatment on cell apoptosis (Fig. 2D). In brief, UA downregulated ABCC1 to hinder the growth of DOX-resistant TNBC cells.
MiR-186-5p targeted ABCC1
Thereafter, the upstream mechanism of ABCC1 was explored. Since the ceRNA pattern was widely involved in the regulation of protein-coding genes, we wondered if ABCC1 was a participant in certain ceRNA network. Hence, miRNAs that might bind to ABCC1 were searched by starBase v2.0 as described. 22
As was screened by three programs (miRmap, microT, and miRanda) included in starBase v2.0, total five miRNAs were identified (Fig. 3A). Afterward, we enhanced the expression of these six miRNAs in DOX-MDA-MB-468 and DOX-MDA-MB-436 cells (Supplementary Fig. S2B), and figured out that only the upregulation of miR-186-5p could reduce the luciferase activity of ABCC1 while that of other 5 miRNAs had no significant impacts on ABCC1 luciferase activity (Fig. 3B). Based on this, here we selected miR-186-5p as the upstream regulator of ABCC1.

MiR-186-5p targeted ABCC1.
Subsequently, the binding sites for miR-186-5p and ABCC1 were obtained from starBase, and the sequences of ABCC1-WT/Mut with wild-type or mutant miR-186-5p binding sites were also shown (Fig. 3C). Then, luciferase reporter assay results revealed that the luciferase activity of wild-type ABCC1 was significantly decreased by miR-186-5p upregulation, while that of mutant ABCC1 was not impacted, which could be seen in both cells with or without UA treatment (Fig. 3D and Supplementary Fig. S2C). More importantly, it was identified that upregulation of miR-186-5p caused a decrease in ABCC1 expression in two DOX-resistant TNBC cells, regardless of UA treatment (Fig. 3E and Supplementary Fig. S2D).
In the meantime, we also decreased the expression of miR-186-5p by means of miR-186-5p inhibitor (Supplementary Fig. S2E) and figured out that downregulation of miR-186-5p elevated the expression of ABCC1 (Fig. 3F and Supplementary Fig. S2F).
After that, colony formation assay and flow cytometry analysis were carried out in DOX-resistant MDA-MB-468 cells with or without UA treatment. The results manifested that upregulation of miR-186-5p suppressed cell proliferation and promoted cell apoptosis in control groups, while further fortified the proliferation-restraining and apoptosis-accelerating influences on cells under UA treatment (Fig. 3G, H). In conclusion, miR-186-5p targeted ABCC1 to mediate the suppression of UA on DOX-resistant TNBC cell growth.
ZEB1-AS1 served as the ceRNA of ABCC1 through sponging miR-186-5p
Then, we planned to find out the lncRNA that was involved in the ceRNA network consisting of miR-186-5p and ABCC1. Through screening starBase by the condition of low stringency in Degradome data, five lncRNAs possessing miE-186-5p binding sites were uncovered. Among the five candidate lncRNAs, only ZEB1-AS1 was remarkably pulled down by miR-186-5p biotin probe (Fig. 4A). Also, ZEB1-AS1 was downregulated by UA treatment in DOX-resistant cells (Fig. 4B). Based on these data, we further focused on ZEB1-AS1 in the following work.
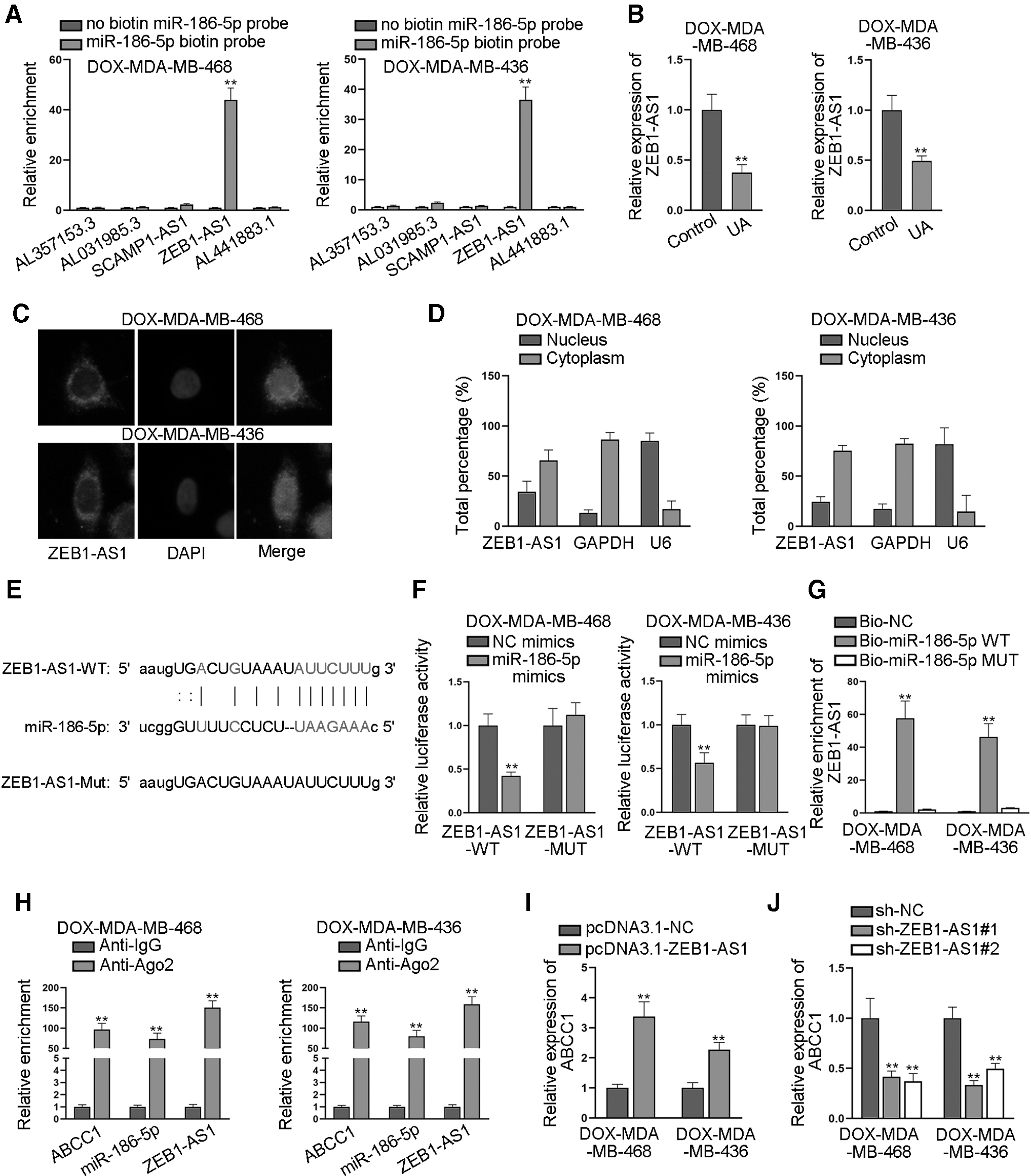
ZEB1-AS1 served as the ceRNA of ABCC1 through sponging miR-186-5p.
FISH and subcellular fraction assays were implemented to determine the subcellular distribution of ZEB1-AS1. The results presented that ZEB1-AS1 mainly resided in the cytoplasm of DOX-MDA-MB-468 and DOX-MDA-MB-436 cells (Fig. 4C, D), which supported that ZEB1-AS1 might serve as a ceRNA in these cells. The putative binding sites between ZEB1-AS1 and miR-186-5p were obtained from starBase (Fig. 4E). We then mutated the binding sites in ZEB1-AS1 to examine whether these sites are responsible for the interaction between ZEB1-AS1 and miR-186-5p. The outcomes of following luciferase reporter assay revealed that upregulation of miR-186-5p significantly reduced the luciferase activity of wild-type ZEB1-AS1 but had no impacts on that of mutant ZEB1-AS1 (Fig. 4F and Supplementary Fig. S3A).
The data of RNA pull-down assay showed that ZEB1-AS1 was significantly pulled down by biotin-labeled wild-type miR-186-5p, while its existence was difficult to be observed in the complex pulled down by biotin-labeled mutant miR-186-5p (Fig. 4G and Supplementary Fig. S3B). Moreover, RIP assay was implemented, and the results depicted that ZEB1-AS1, miR-186-5p, and ABCC1 were all abundantly enriched in the Ago2-precipitated RISCs (Fig. 4H and Supplementary Fig. S3C). Further, we enhanced ZEB1-AS1 expression by transfecting pcDNA3.1-ZEB1-AS1 into DOX-resistant cells (Supplementary Fig. S3D), and found that upregulation of ZEB1-AS1 significantly enhanced the expression of ABCC1 (Fig. 4I and Supplementary Fig. S3E). Similarly, knockdown of ZEB1-AS1 by specific shRNAs significantly reduced the expression of ABCC1 (Fig. 4J and Supplementary Fig. S3F).
Based on these findings, we drew a conclusion that ZEB1-AS1 was the ceRNA of ABCC1 through sponging miR-186-5p.
UA elevated the growth of DOX-resistant TNBC cells through ZEB1-AS1/miR-186-5p/ABCC1 axis
Finally, rescue assays were implemented to validate whether ABCC1 mediated the effects of ZEB1-AS1 in DOX-resistant TNBC cells with or without UA treatment. At first, the overexpression efficiency of ABCC1 was validated by qRT-PCR (Supplementary Fig. S3G). It was revealed that upregulation of ABCC1 completely rescued the effects of silenced ZEB1-AS1 on cell proliferation in DOX-resistant cells with or without UA treatment (Fig. 5A, B). Moreover, flow cytometry analysis discovered that the promoting effects of silenced ZEB1-AS1 on cell apoptosis were completely restored by upregulation of ABCC1 in DOX-resistant cells with UA treatment or not (Fig. 5C). Taken together, UA hindered the growth of DOX-resistant TNBC cells through mediation on ZEB1-AS1/miR-186-5p/ABCC1 pathway.
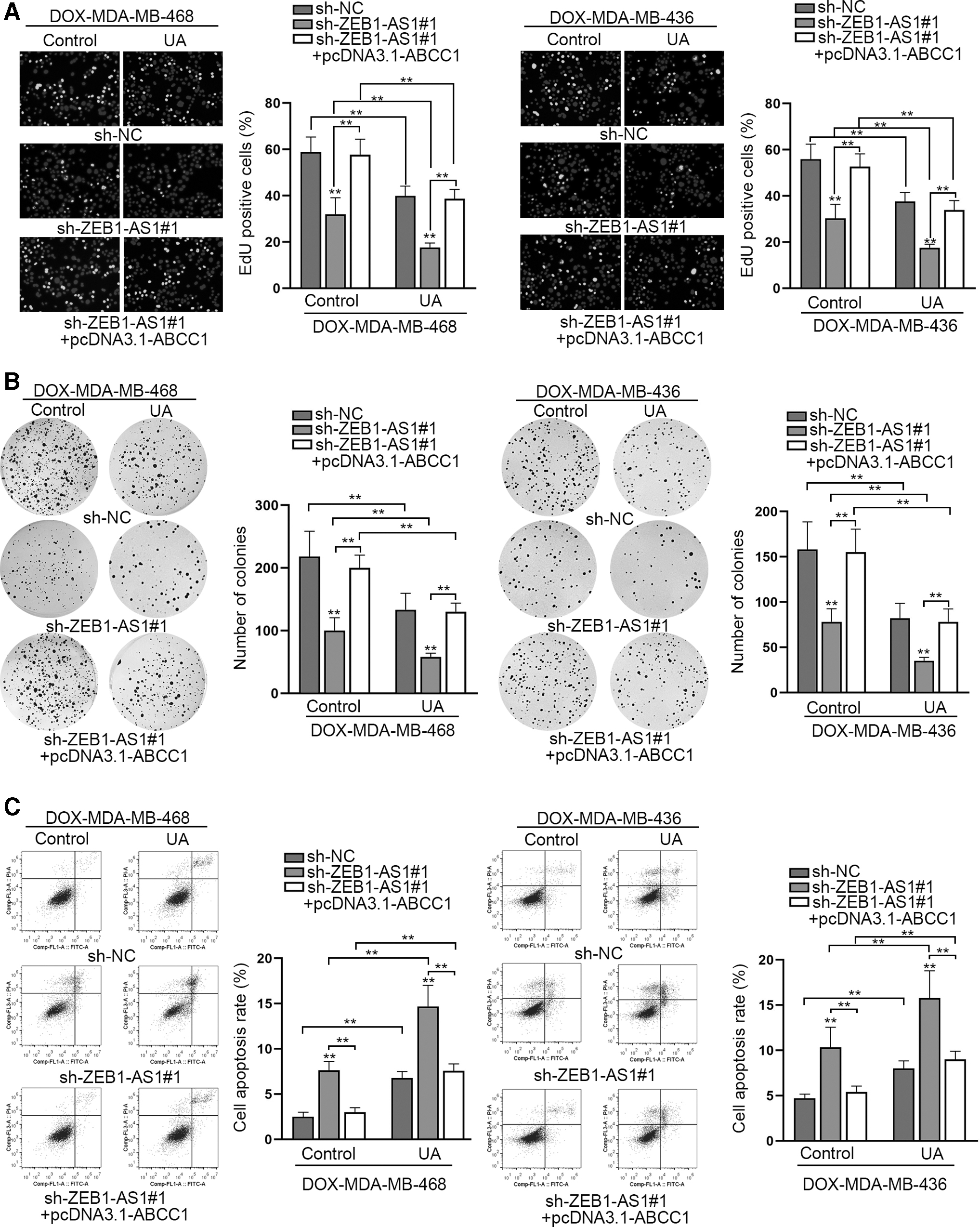
UA suppressed the growth of DOX-resistant TNBC cells through ZEB1-AS1/miR-186-5p/ABCC1 axis.
Discussion
DOX is a preferred chemotherapeutic option for TNBC treatment. However, after long-term exposure, the effect of DOX is usually limited because of inevitable drug resistance. UA, as well as other small molecules, has been reported to reverse drug resistance in BC cells. 23 This study started from UA to explore its function in DOX-resistant TNBC cells. It was first revealed that UA treatment significantly reduced cell proliferation and promoted cell apoptosis in DOX-resistant TNBC cells. Besides, cotreatment of UA strengthened the impairment of DOX on the growth of DOX-resistant TNBC cells. In other words, UA treatment strengthened chemosensitization of DOX-resistant TNBC cells. Then, we sought to explore the underlying mechanism of UA in the growth and DOX resistance of TNBC cells.
ABCC1 is a member of ABC drug transporters and is closely associated with multidrug resistance in various cancers. For instance, MDIG promotes resistance to cisplatin in lung adenocarcinoma cells by regulating ABCB1, ABCC1, and ABCG2 expression through triggering WNT/β-catenin signaling. 24 LncRNA GHET1 facilitates drug resistance to gemcitabine in bladder cancer cells through ABCC1 upregulation. 25 Oncogenic PITX2 facilitates drug resistance by regulating hOCT3/SLC22A3 and ABC drug transporters, including ABCB1, ABCC1, ABCG2 in colon and kidney cancers. 26 Noticeably, a former research unveiled that ABCC1 is highly expressed in aggressive BC subtypes like TNBC. 27
This study figured out that ABCC1 was downregulated in DOX-resistant TNBC cells under UA treatment. Also, ABCC1 inhibition caused a decrease in cell proliferation but an increase in cell apoptosis in DOX-resistant TNBC cells. Importantly, silencing ABCC1 reinforced the suppression of UA treatment on the growth of DOX-resistant TNBC cells.
Since miRNAs are widely reported to target mRNAs, the putative miRNAs binding to ABCC1 were identified by bioinformatics and mechanistic assays. It was discovered that miR-186-5p targeted ABCC1 and inhibited the growth of DOX-resistant TNBC cells. MiR-186-5p was previously reported to repress cisplatin resistance in nonsmall-cell lung cancer cells through SIX1 inhibition. 28 Besides, it plays a role as a tumor suppressor in colorectal cancer, 29 gastric cancer, 30 and osteosarcoma. 31
Recently, the ceRNA pattern has been proposed as a typical mechanism upstream miRNAs/mRNAs. In addition, lncRNAs have been uncovered to play vital roles in cancer development and drug resistance. Here, we went on to explore the upstream lncRNA of miR-186-5p/ABCC1 axis. The mechanistic investigations revealed that ZEB1-AS1 served as an endogenous sponge of miR-186-5p to activate ABCC1 through the ceRNA pattern. ZEB1-AS1 was downregulated in DOX-resistant TNBC cells after UA treatment. As is discovered by Chai et al., ZEB1-AS1 is significantly upregulated in tissues from patients with gastric cancer receiving chemotherapy. 32 Also, ZEB1-AS1 itself is verified as the tumor inducer in multiple cancers. For instance, depletion of ZEB1-AS1 inhibits cell proliferation and invasion in esophageal squamous cell carcinoma. 33
Moreover, rescue assays performed in this study unveiled that the contribution of ZEB1-AS1 deficiency to UA treatment induced growth-inhibiting influence on DOX-resistant TNBC cells was offset by ABCC1 overexpression. Considering the findings of recent research, which suggested that ZEB1-AS1 facilitates TNBC progression 34 and also induces chemoresistance in BC cells, 35 we deduced that UA relied on ZEB1-AS1-mediated miR-186-5p/ABCC1 signaling to hamper the growth and DOX resistance of TNBC cells.
Conclusion
This study uncovered that UA hindered the growth and reversed the DOX resistance of DOX-resistant TNBC cells through inactivating ZEB1-AS1/miR-186-5p/ABCC1 signaling, which might shed insight into overcoming DOX resistance in TNBC.
Footnotes
Acknowledgment
We sincerely appreciate all laboratory members.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary File S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.