
Abstract
Background:
Lung cancer is a social problem of increasing concern, and non-small cell lung cancer (NSCLC) accounts for 80%–85% incidence of lung cancer. Cisplatin (DDP) is reported as a first-line chemotherapy drug for NSCLC, but the resistance has became a main obstacle for NSCLC treatment. The high level of circular RNA circ_0076305 was related to the DDP resistance in NSCLC. However, the mechanism of circ_0076305 remains unclear in DDP resistance of NSCLC.
Materials and Methods:
Exosomes were detected by a transmission electron microscope and nanoparticle tracking analysis. The protein levels of CD63, CD81, P-glycoprotein (P-gp), Lung resistance-related protein, and ATP-binding cassette subfamily C member 1 (ABCC1) were examined by Western blot assay. Circ_0076305, microRNA-186-5p (miR-186-5p), and ABCC1 levels were tested by real-time quantitative polymerase chain reaction. DDP resistance was examined by 3-(4, 5-dimethyl-2-thiazolyl)-2, 5-diphenyl-2-H-tetrazolium bromide assay. The binding relationship between miR-186-5p and circ_0076305 or ABCC1 was predicted by circRNA interactome or starBase, and then verified by dual-luciferase reporter and RNA immunoprecipitation assays. The effect of circ_0076305 on DDP resistance in NSCLC was examined by xenograft tumor model in vivo.
Results:
Circ_0076305 was increased in NSCLC cell-derived exosomes, DDP-resistant NSCLC tissues and cells. Circ_0076305 knockdown elevated DDP sensitivity in vitro. Mechanically, circ_0076305 enhanced ABCC1 expression through sponging miR-186-5p, thus regulating DDP resistance of NSCLC. Furthermore, circ_0076305 silencing improved DDP sensitivity of NSCLC in vivo.
Conclusion:
The results from this study disclosed that circ_0076305 knockdown improved DDP sensitivity by the miR-186-5p/ABCC1 axis in NSCLC, hinting a potential circRNA-targeted therapy for NSCLC.
Introduction
An incurable cancer, lung cancer has gradually become a social problem of increasing concern, with ∼228,150 new cases and 142,670 deaths in the United States in 2019. 1 Currently, as the major subtype of lung cancer, non-small cell lung cancer (NSCLC) accounts for 80%–85% incidence of lung cancer. 2 Due to the inadequate screening program and lack of specific symptoms in the early stage, most NSCLC patients are diagnosed at an advanced stage with a poor prognosis. 3 Notably, initially, the majority of patients with NSCLC received successful chemotherapy and radiotherapy at the advanced stage. 4 –6 Nevertheless, tumors often recur owing to the development of multidrug resistance (MDR). 7,8 In recent years, cisplatin (DDP) is considered a first-line chemotherapy drug for NSCLC, 9 but its resistance has become the main cause of treatment failure. 10 Hence, to identify an effective therapeutic target for DDP sensitive in NSCLC, it is necessary to expound the molecular mechanism of drug resistance.
Over the last decade, circular RNAs (circRNAs), a novel subgroup of the noncoding transcript, have been reported to be widely expressed in the cytoplasm of mammalian cells. 11 Accumulating evidence suggested that circRNAs, as oncogenes or tumor suppressors, participated in the occurrence and development in diverse cancers. 12,13 Furthermore, some studies have confirmed that the dysregulation of circRNAs was closely linked to the chemoresistance of multiple cancers, including NSCLC. 14 The upregulation of circRNAhsa_circ_0004015 was confirmed to improve proliferation, invasion, and TKI drug resistance through regulating the miR-1183/PDPK1 axis in NSCLC. 15 Moreover, Li et al. showed that circRNAhsa_circ_0002483 could repress the progression and boost the taxol sensitivity in NSCLC by sponging mir-182-5p. 16 Importantly, recent literature confirmed that circRNA circ_0076305 was increased in NSCLC and associated with the DDP resistance of NSCLC. 17 Yet, the underlying role of circ_0076305 in the DDP resistance of NSCLC is still unclear.
Currently, microRNAs (miRNAs), a kind of endogenous noncoding RNA at the length of 19–25 nucleotides, could inhibit gene expression by complementary binding with the 3′-untranslated region (3′UTR) of target mRNAs. 18 It is becoming increasingly apparent that abnormal expression of miRNAs is correlated with the drug resistance and pathology in various tumors, including NSCLC. 19 –21 MicroRNA-186-5p (miR-186-5p), a form of mature miR-186, exerts the tumor suppressor in a variety of cancers, such as osteosarcoma, 22 gastric cancer, 23 and NSCLC. 24 Intriguingly, previous studies displayed that miR-186 could repress the drug resistance through interacting with target genes in NSCLC, 25,26 implying the important role of miR-186-5p in drug resistance of NSCLC.
ATP-binding cassette subfamily C member 1 (ABCC1), also known as MRP1, has been reported to have three transmembrane domains and two cytoplasmic nucleotide-binding domains. Widely expressed in human lung, kidney, and spleen, ABCC1 could transport a variety of substrates, inducing anticancer drugs. 27,28 Moreover, some publications suggested that ABCC1 played an important role in the chemoresistance of diverse tumors. 29,30 Interestingly, in DDP-resistant NSCLC cells, ABCC1 was found to be upregulated, and the abnormal expression of ABCC1 promoted the resistance of DDP in NSCLC. 31,32 These results implied the vital function of ABCC1 in DDP-resistant NSCLC.
In this study, their data showed that circ_0076305 was increased in DDP-resistant NSCLC cells, and expedited DDP resistance in NSCLC. Furthermore, bioinformatics analysis first suggested that circ_0076305 has some complementary sites with miR-186-5p, and the underlying binding between miR-186-5p and ABCC1. Hence, the authors aimed to explore whether circ_0076305 could regulate the resistance of DDP in NSCLC through the miR-186-5p/ABCC1 axis.
Materials and Methods
Clinical samples and cell culture
All experiments were conducted in compliance with the Ethics Committee of Weifang People's Hospital. The written informed consent was provided by each participant. Samples of serum from NSCLC patients (n = 50) and healthy volunteers (n = 50), and 50 pairs of NSCLC tumor tissues and adjacent normal tissues from NSCLC patients were collected from Weifang People's Hospital. Moreover, these NSCLC patients were classified as DDP-sensitive patients and DDP-resistant patients based on the previous study. 33
Human bronchial epithelial cells (BEAS-2B) and NSCLC cell lines (A549 and H292) were provided by American Type Culture Collection (ATCC, Manassas, VA) and grown in humidified atmosphere containing 5% CO2 at 37°C with Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA). Notably, 10% fetal bovine serum (FBS; Gibco) as the supplement was added to the culture medium. In addition, to obtain cisplatin (DDP)-resistant NSCLC cells (A549/DDP and H292/DDP), A549 and H292 cells were continuously stimulated with DDP (2 μM; Sigma-Aldrich, St. Louis, MO) and established as previously described. 34
Exosome detection
In accordance with the direction of the supplier, the exosomes in serums were isolated and purified using the ultracentrifugation method and the Total Exosome Isolation Kit (Invitrogen). Then, a transmission electron microscopy (Hitachi, Tokyo, Japan) was applied to view the exosomes in fixative. Besides, the size distribution of exosomes was analyzed by the nanoparticle tracking analysis (NTA) (Nanosight NS300; Nanosight, Amesbury, United Kingdom). For inhibition of exosome production, A549 and H292 cells were treated with or untreated the inhibitor of exosome release GW4869 (10 μM; Sigma-Aldrich). After incubation for 48 h, the expression level of circ_0076305 was detected using real-time quantitative polymerase chain reaction (RT-qPCR) assay.
Western blot assay
In this assay, RIPA buffer containing protease and phosphatase inhibitors (Beyotime, Nantong, China) were used to obtain the protein lysates form samples, which then were separated by a sodium dodecyl sulfate/polyacrylamide gel electrophoresis system. Whereafter, the separated proteins were transferred onto nitrocellulose membranes (Millipore, New York, NY), and blocked in 5% skim milk for 2 h. After incubation with primary antibodies against CD81 (1:1000, ab155760; Abcam, Cambridge, MA), CD63 (1:1000, ab216130; Abcam), P-glycoprotein (P-gp; 1:1000, ab170904; Abcam), lung resistance-related protein (LRP, 1:1000, ab97311; Abcam), ABCC1 (1:1000, ab9531; Abcam), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH; 1:1000, ab9485; Abcam) at 4°C overnight, the members were incubated with the corresponding secondary antibodies for 1 h. Finally, an enhanced chemiluminescence reagent (ECL; GE Health Care, Piscataway, NJ) was utilized to analyze the protein bands.
Real-time quantitative polymerase chain reaction
Briefly, TRIzol reagent (Invitrogen) was applied to extract total RNA from the samples, followed by quantification with NanoDrop (NanoDrop Technologies, Wilmington, WI). Then, with the help of PrimeScript™ RT Master Mix (TaKaRa, Dalian, China), the extracted RNAs were reversely transcribed into the first-strand complementary DNA (cDNA). RT-qPCR was conducted on 7500 Real-Time PCR Systems (Applied Biosystems, Darmstadt, Germany) with the SYBR Green PCR Kit (TaKaRa). Finally, the 2–ΔΔCt method was used to calculate relative RNA expression, and GAPDH or U6 small nuclear RNA (snRNA) worked as the internal reference. The primer sequences are as follows: circ_0076305: 5′-CAGTCACTCCCGCTTCAACC-3′ (sense), 5′-CTAGGAGGGTGCAGGGTCAA-3′ (antisense); miR-186-5p: 5′-GCGGATCCGAGCCATGCTTATGCTACTG-3′ (sense), 5′-GCGCGGCCGCCCAGGTATATGGCA-3′ (antisense);
ABCC1: 5′-TGCAAGTGATTAGCCAGGTG-3′ (sense), 5′-CCAGGGGAAAATAACGTGAA-3′ (antisense); U6: 5′-CTCGCTTCGGCAGCACA-3′ (sense), 5′-AACGCTTCACGAATTTGCGT-3′ (antisense); GAPDH: 5′-GTCAACGGATTTGGTCTGTATT-3′ (sense), 5′-AGTCTTCTGGGTGGCAGTGAT-3′ (antisense).
Cell transfection
Circ_0076305 small interfering RNA (si-circ_0076305) and scrambled siRNA control (si-NC), miR-186-5p mimic, miR-186-5p inhibitor, and their negative controls (miRNA NC and inhibitor NC) were obtained from RiboBio (Guangzhou, China). Overexpression of circ_0076305 and ABCC1 was obtained through inserting circ_0076305 and ABCC1 sequences into pcDNA vector (Invitrogen), named as pcDNA-circ_0076305 (circ_0076305) and pcDNA-ABCC1 (pc-ABCC1). Moreover, pcDNA empty vector (Invitrogen) worked as their respective control (termed control and pc-NC). Whereafter, these oligonucleotides and plasmids were transfected with A549/DDP and H292/DDP cells, referring to the operation manual of the Lipofectamine 3000 reagent (Invitrogen).
Drug resistance assay
The 3-(4, 5-dimethyl-2-thiazolyl)-2, 5-diphenyl-2-H-tetrazolium bromide (MTT; Sigma-Aldrich) was utilized to assess the DDP resistance in DDP-resistant NSCLC cells. In brief, in a 96-well plate, NSCLC cells were exposed to 5 μg/mL DDP for 24 h. After incubation for another 48 h, each well was added 20 μL MTT solution (Sigma-Aldrich), followed by maintenance for 4 h. Then, 150 μL dimethyl sulfoxide (DMSO; Sigma-Aldrich) was added to dissolve the formed formazan crystals. Finally, the absorbance was examined under a microplate reader at 570 nm, and data were displayed as the inhibitory rate. The concentration of DDP triggering 50% inhibition of growth (IC50) was shown with the relative survival curve.
Dual-luciferase reporter assay
The sequences of circ_0076305 and 3′UTR of ABCC1 harboring putative binding sites of wild-type (WT) miR-186-5p or mutated miR-186-5p were cloned and inserted into pGL3-control vectors (Promega, Madison, WI), obtaining WT/mutant type (MUT)-circ_0076305 and WT/MUT-ABCC1 reporter vectors. According to the instruction guidelines of Lipofectamine 3000 reagent (Invitrogen), DDP-resistant NSCLC cells were cotransfected with the reporter vectors with miRNA NC and miR-186-5p mimic, followed by incubation for 48 h. Then, with the help of a dual-luciferase reporter assay system (Promega), the luciferase activities were assessed in NSCLC cell lysates.
RNA immunoprecipitation
In this assay, the biotin-labeled circ_0076305 probe was specifically designed and synthesized by GenePharma (Shanghai, China), and oligo probe worked as a control (scrambled probe), followed by transfection into NSCLC cells. Whereafter, in the formaldehyde, the crosslinking reaction was completed, and then the glycine solution was used to stop the reaction. After washing with phosphate-buffered saline (PBS; Invitrogen), these cells were lysed by using the sonication, followed by collection with the supernatant. Then, M-280 Streptavidin magnetic beads (Invitrogen) were mixed with the supernatant at room temperature. The next day, the lysis buffer with proteinase K was applied to wash and reverse crosslinking the mixture. After total RNA extraction, RT-qPCR was conducted to analyze the enrichment of circ_0076305 and miR-186-5p.
Tumor xenograft assay
Generally, the synthesis of circ_0076305 knockdown stable lentiviral strain (short hairpin RNA expression vector, sh-circRNA), and the lentiviral empty vector as a control (sh-NC) was provided by GeneChem (Shanghai, China). Five-week-old male BALB/C nude mice (n = 4 per group) were obtained from the National Laboratory Animal Center (Beijing, China). This research was approved by the Animal Ethics Committee of the Weifang People's Hospital. Transfected A549/DDP cells (7 × 106) were subcutaneously injected into the left flank of the nude mice. At 1 week after injection, PBS (Invitrogen) or 5 mg/kg DDP were intraperitoneally administrated into mice every 4 d. Finally, after 27 d later, the tumors were excised and weighed, followed by utilization for RT-qPCR assay.
Statistical analysis
GraphPad Prism7 software was applied for data analysis. If the p-value was <0.05, it was considered to be statistically significant. Data of three independent experiments were exhibited as mean ± standard deviation (SD). Student's t-test was used for comparisons between two groups, and one-way analysis of variance (ANOVA) with Tukey's tests were utilized for comparisons among multiple groups.
Results
Circ_0076305 was upregulated in exosomes
First of all, exosomes were isolated from serums of NSCLC patients and normal subjects by using ultracentrifugation. Then, the transmission electron microscope and NTA were applied to examine the morphology of exosomes. Results presented that exosomes were typical rounded particles, and most of them were within a diameter of 70–150 nm (Fig. 1A, B). Moreover, exosomal markers, CD63 and CD81, were observed in exosomes from both groups (Fig. 1C). Interestingly, the authors found that circ_0076305 level was upregulated in serum and exosome derived from serum of NSCLC patients compared with respective control groups (Fig. 1D, E). Therefore, to explore whether exosomal circ_0076305 was delivered into NSCLC cells, exosomes from serums or PBS were coincubated with NSCLC cell lines (A549 and H292 cells). As displayed in Figure 1F, compared with cells treated with PBS, circ_0076305 level was increased in cells treated with exosomes. In addition, they further verified that the expression level of circ_0076305 in A549 and H292 cells treated with GW4869, which inhibited the release of exosomes, was lower than that in cells untreated with GW4869 (Fig. 1G), suggesting that tumor cells could increase the expression of circ_0076305 along with exosome production, and circ_0076305 might affect tumor cells through autocrine/paracrine mechanism.

Circ_0076305 was upregulated in exosomes.
Circ_0076305 was increased in DDP-resistant NSCLC tissues and cells
To explore the correlation of circ_0076305 with DDP resistance in NSCLC, circ_0076305 level was detected by RT-qPCR assay in NSCLC tissues and DDP-resistant NSCLC tissues. As exhibited in Figure 2A, B, circ_0076305 was increased in NSCLC tissues (n = 50) and DDP-resistant NSCLC tissues (n = 25) relative to their respective control groups. Similarly, a dramatic increase in circ_0076305 expression was viewed in DDP-resistant NSCLC cells (A549/DDP and H292/DDP) when compared with human bronchial epithelial cells (BEAS-2B) and NSCLC cell lines (A549 and H292). These results suggested the involvement of circ_0076305 with DDP resistance in NSCLC. Moreover, to investigate the treatment of DDP on circ_0076305 level in NSCLC cells, 5 μg/mL DDP was applied for A549 and H292 cells for 0, 24, 48, and 72 h. The results revealed that DDP treatment increased the intracellular circ_0076305 levels in A549 and H292 cells in a time-dependent manner (Fig. 2D, E), especially in 24 h. Therefore, 24 h was chosen for subsequent experiments.
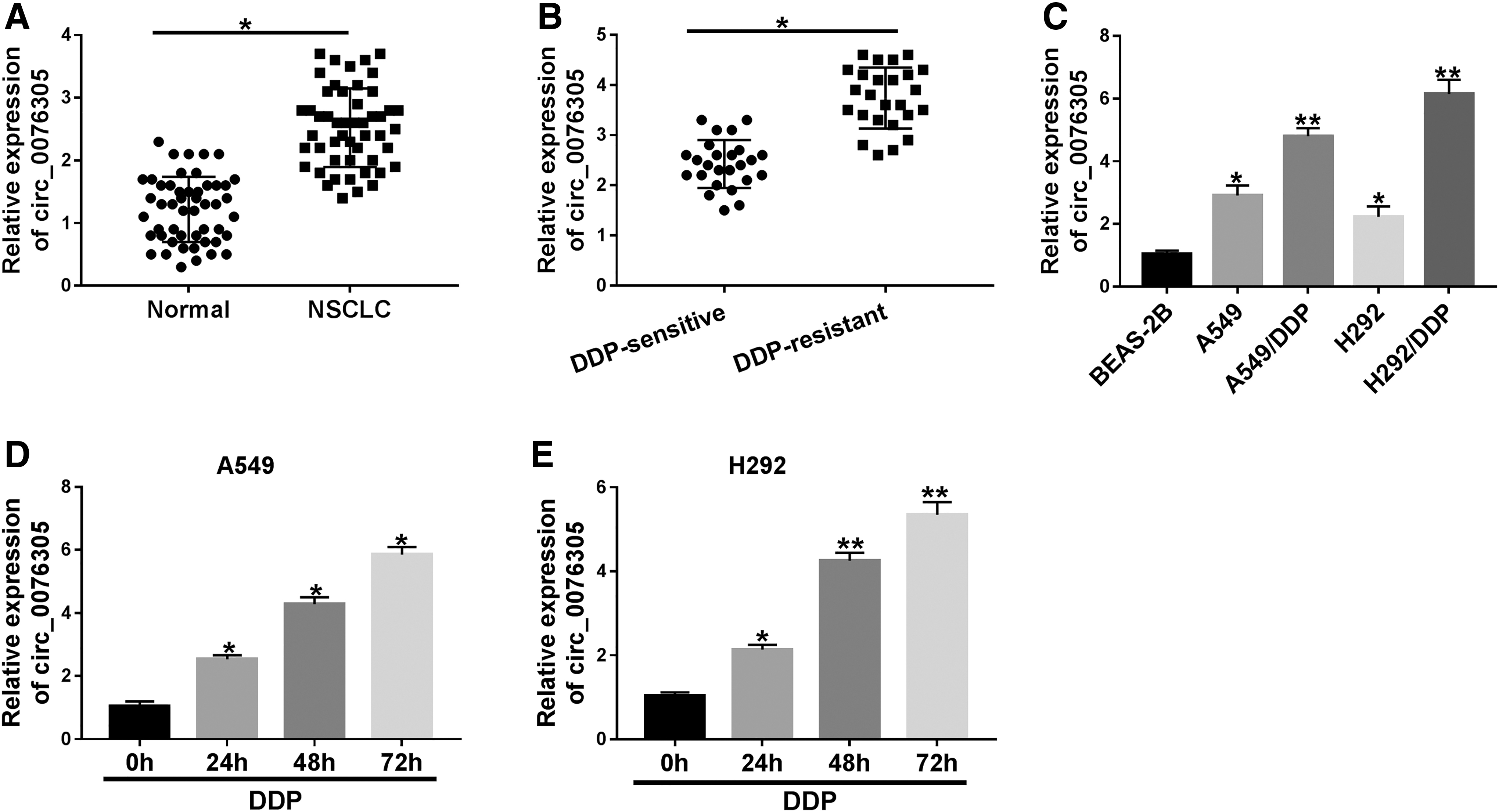
Expression of circ_0076305 was increased in DDP-NSCLC tissues and cells.
Circ_0076305 knockdown improved DDP sensitivity in DDP-resistant NSCLC cells
Considering the high expression of circ_0076305 in DDP-resistant NSCLC cells, they knocked downcirc_0076305 in NSCLC cells. As shown in Figure 3A, circ_0076305 level was markedly downregulated in si-circ_0076305 transfected in A549/DDP and H292/DDP cells versus cells transfected with si-NC. Moreover, the effect of DDP on circ_0076305 level was detected in DDP-resistant NSCLC cells. Data exhibited that the expression level of circ_0076305 was notably increased in A549/DDP and H292/DDP cells treated with 5 μg/mL DDP for 24 h relative to cells treated with 0 μg/mL or 2 μg/mL DDP (Fig. 3B). Thus, 5 μg/mL DDP was selected for the following research. Furthermore, IC50 determination presented that circ_0076305 silencing evidently reduced DDP resistance in A549/DDP (Fig. 3C, D) and H292/DDP cells (Fig. 3E, F), whereas the overexpression of circ_0076305 produced the opposite results. Besides, MDR-related marker proteins, P-gp and LRP, have been confirmed to serve as crucial functions in drug resistance of NSCLC. 35 Results proved that manipulation of circ_0076305 expression changed the protein levels of P-gp and LRP in DDP-resistant NSCLC cells (Supplementary Fig. S1). Collectively, these results suggested that circ_0076305 deficiency sensitized A549/DDP and H292/DDP cells to DDP.
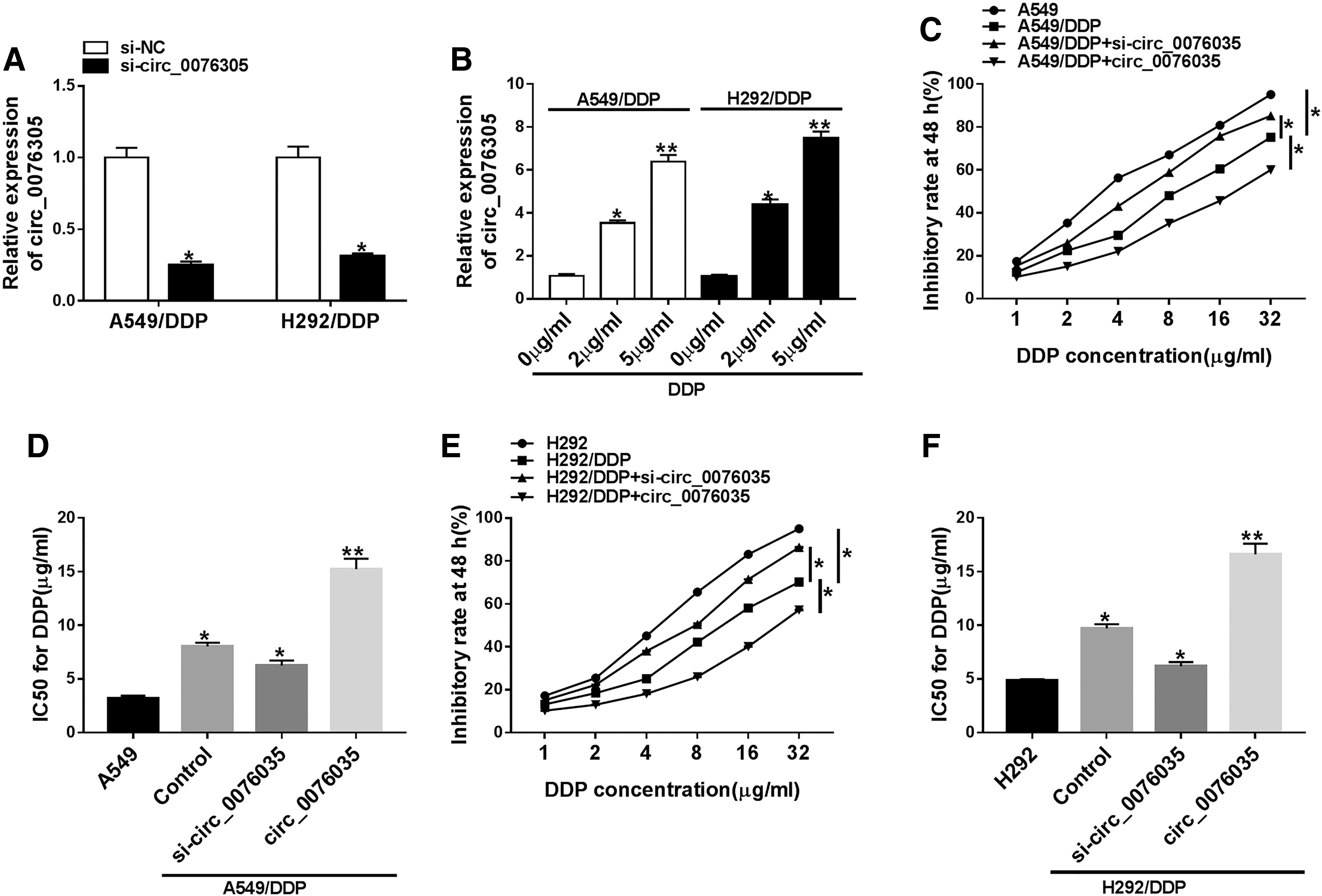
Circ_0076305 knockdown improved DDP sensitivity in DDP-resistant NSCLC cells.
MiR-186-5p was a target of circ_0076305
It is widely accepted that circRNAs could exert the function through interacting with miRNAs. 36 Hence, they searched the underlying target miRNAs of circ_0076305 by circRNA interactome software. As exhibited in Figure 4A, miR-186-5p contained some complementary sites with circ_0076305. Interestingly, they found that miR-186-5p was expressed at the low level in DDP-resistant NSCLC tissues (Fig. 4B) and DDP-resistant NSCLC cells (Fig. 4C) relative to their respective control groups. Importantly, inversely associated with circ_0076305 expression (Fig. 4D). Therefore, they used a dual-luciferase reporter assay to further confirm the correlation between circ_0076305 and miR-186-5p. Data suggested that miR-186-5p overexpression declined the luciferase activity of WT-circ_0076305 reporter vector, whereas had little effect on the luciferase activity of MUT-circ_0076305 reporter vector (Fig. 4E, F). Apart from that, compared with the controls, the expression levels of circ_0076305 and miR-186-5p were enriched in circ_0076305-labeled RNA using the specific circ_0076305 probe (Fig. 4G, H). Besides, they verified that the knockdown of circ_0076305 improved the level of miR-186-5p in A549/DDP and H292/DDP cells (Fig. 4I). Taken together, these results suggested that circ_0076305 could directly bind with miR-186-5p in DDP-resistant NSCLC cells.
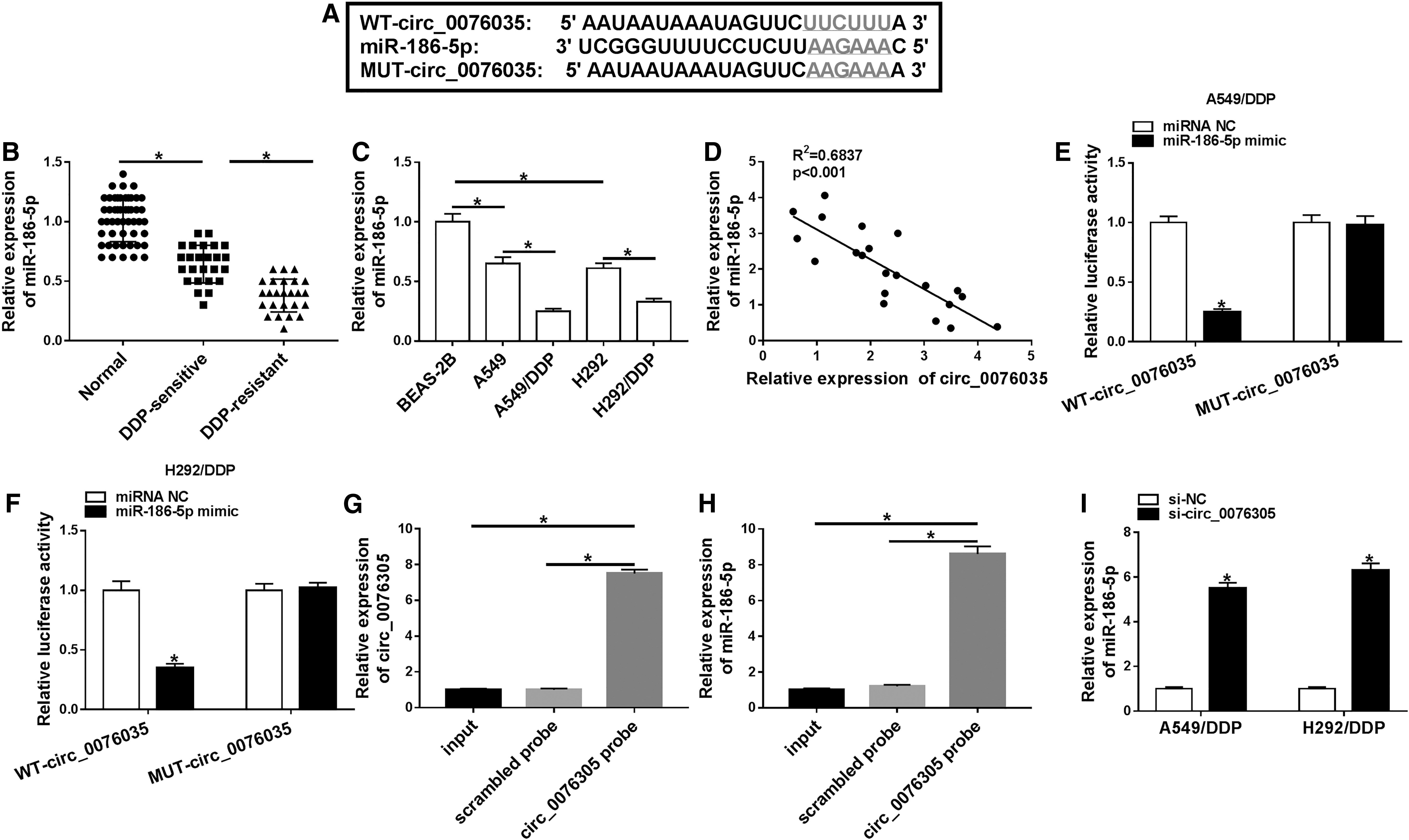
MiR-186-5p acted as a target of circ_0076305.
Circ_0076305 downregulation enhanced DDP sensitivity through negatively modulating miR-186-5p
As mentioned above, circ_0076305 served as a critical role for the DDP resistance in DDP-resistant NSCLC cells, and miR-186-5p was a direct target of circ_0076305 in DDP-resistant NSCLC cells. Therefore, they further explore whether the impact of circ_0076305 on DDP resistance was related to miR-186-5p. As presented in Figure 5A–C, the downregulation of miR-186-5p effectively abrogated si-circ_0076305-induced increase in DDP sensitivity. Furthermore, the protein levels of P-gp and LRP could be regulated by circ_0076305/miR-186-5 in DDP-resistant NSCLC cells (Supplementary Fig. S2). All these data indicated that miR-186-5p silencing could partly reverse circ_0076305-deletion-mediated DDP sensitivity in DDP-resistant NSCLC cells.
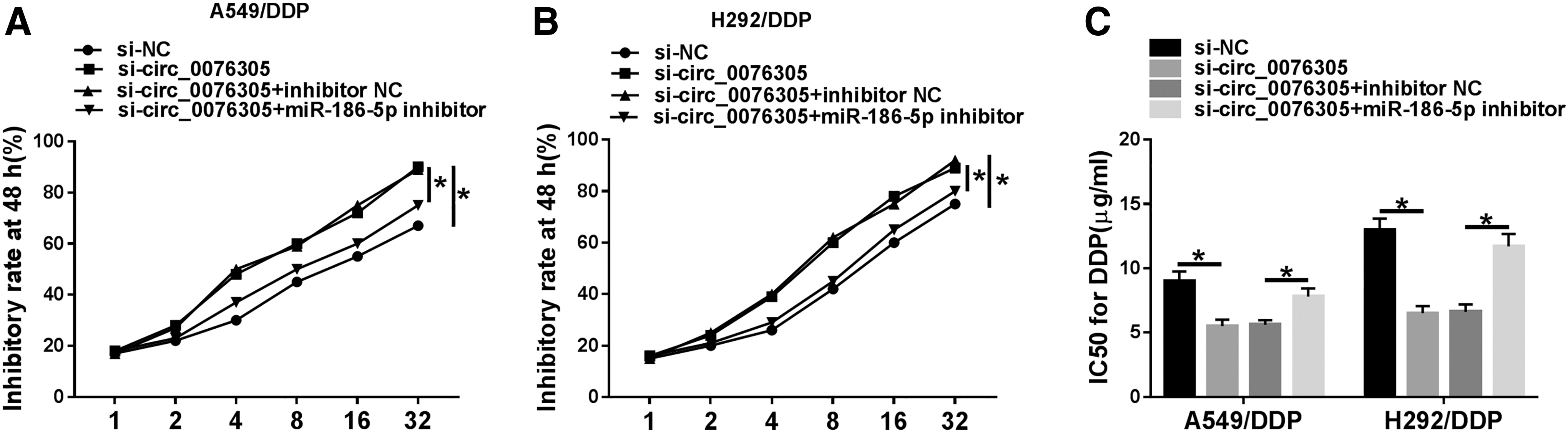
MiR-186-5p deficiency overturned the effects of circ_0076305 downregulation on DDP sensitivity in DDP-resistant NSCLC cells. A549/DDP and H292/DDP cells were transfected with si-NC, si-circ_0076305, si-circ_0076305 + inhibitor NC, and si-circ_0076305 +miR-186-5p inhibitor.
ABCC1 was the target of miR-186-5p
As widely believed, miRNAs could exert the function by interacting with mRNAs. Through the bioinformatics software StarBase analysis, the potential targets of miR-186-5p were predicted. Data showed that there existed some binding sequences between miR-186-5p and ABCC1 (Fig. 6A). Moreover, they found that ABCC1 was upregulated in DDP-resistant NSCLC tissues in comparison with DDP-sensitive NSCLC tissues and Normal group (Fig. 6B, C). Similarly, an obvious increase of ABCC1 level was observed in DDP-resistant NSCLC cells, when compared with their respective control groups (Fig. 6D, E). Furthermore, in accordance with the bioinformatics analysis, a dual-luciferase reporter assay displayed that the overexpression of miR-186-5p reduced the luciferase activity of ABCC1 WT reporter vector in 293T, but not that of ABCC1 MUT reporter vector (Fig. 6F). Meanwhile, miR-186-5p level was negatively correlated with the expression level of ABCC1 in DDP-resistant NSCLC tissues (Fig. 6G). Besides, both mRNA level and protein level of ABCC1 were repressed due to the overexpression of miR-186-5p in DDP-resistant NSCLC cells (Fig. 6H, I), whereas the reintroduction of pc-ABCC1 remarkably overturned the inhibitory effect of miR-186-5p mimic on ABCC1 level (Fig. 6J, K). These results suggested that ABCC1 was a direct target of miR-186-5p in DDP-resistant NSCLC cells.

ABCC1 acted as the target of miR-186-5p.
ABCC1 overexpression partially reversed the inductive effect of miR-186-5p on DDP sensitivity in DDP-resistant NSCLC cells
Then, they further explored whether miR-186-5p could regulate DDP resistance by targeting ABCC1 in DDP-resistant NSCLC cells. Drug resistance assay showed that the overexpression of miR-186-5p improved DDP sensitivity in A549/DDP and H292/DDP cells, which was drastically abrogated through the upregulation of ABCC1 (Fig. 7A–C). Synchronously, they detected the effect of miR-186-5p and ABCC1 on the protein levels of P-gp and LRP in A549/DDP and H292/DDP cells. Additionally, the levels of P-gp and LRP could be mediated by the miR-186-5p/ABCC1 in DDP-resistant NSCLC cells (Supplementary Fig. S3). In a word, miR-186-5p could enhance DDP sensitivity by regulating ABCC1 in DDP-resistant NSCLC cells.

MiR-186-5p-mediated DDP sensitivity was reversed through the upregulation of ABCC1 in DDP-resistant NSCLC cells. A549/DDP and H292/DDP cells were transfected with miRNA NC, miR-186-5p mimic, miR-186-5p mimic + pc-NC, and miR-186-5p mimic + pc-ABCC1.
Circ_0076305 regulated ABCC1 expression through targeting miR-186-5p in DDP-resistant NSCLC cells
As mentioned above, they speculated that circ_0076305 could exert the function by modulating the miR-186-5p/ABCC1 axis in DDP-resistant NSCLC cells. Simultaneously, circRNAs have been confirmed as competing endogenous RNAs (ceRNAs) that share miRNA recognition sites with mRNAs or compete for miRNA binding, thus resulting in changes of target mRNAs expression. To verify the hypothesis, the effect of circ_0076305 on ABCC1 expression was further detected. As shown in Figure 8A and B, circ_0076305 knockdown hindered the level of ABCC1 in DDP-resistant NSCLC cells, and the reintroduction of pc-ABCC1 abated the effect. Furthermore, rescue assays confirmed that miR-186-5p inhibitor could partly overturn the negative influence of circ_0076305 deficiency on ABCC1 level in A549/DDP and H292/DDP cells (Fig. 8C, D). All of these data suggested that circ_0076305 could affect ABCC1 expression through acting as a molecular sponge of miR-186-5p in DDP-resistant NSCLC cells.

Circ_0076305 affected ABCC1 expression by sponging miR-186-5p in DDP-resistant NSCLC cells.
Circ_0076305 deficiency suppressed tumor growth and improved DDP sensitivity in NSCLC in vivo
Additionally, they established a xenograft tumor mice model to further prove the effectiveness of circ_0076305 on DDP resistance in vivo. As illustrated in Figure 9A, B, DDP injection or circ_0076305 deletion reduced the tumor volume and weight, implying that DDP treatment or circ_0076305 downregulation could repress NSCLC tumor growth in vivo. Notably, combined circ_0076305 silencing and DDP resulted in a more overt suppression on tumor growth. Furthermore, circ_0076305 level was apparently decreased in tumor tissues from the sh-circ_0076305 group relative to the sh-NC group (Fig. 9C). These data proved that circ_0076305 deficiency elevated DDP sensitivity of NSCLC in vivo.
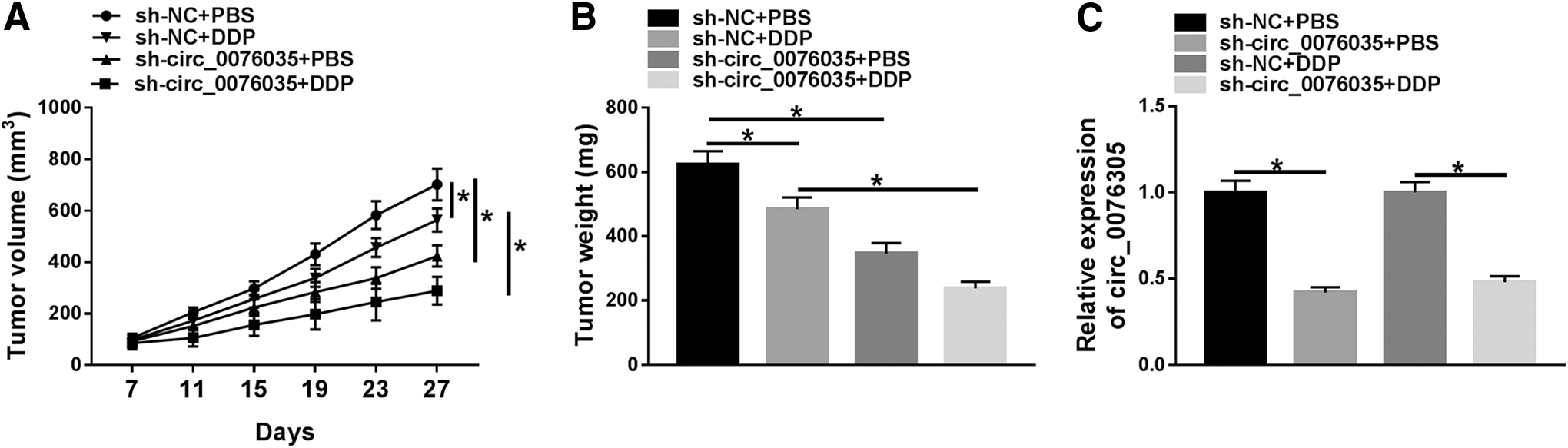
Circ_0076305 deficiency enhanced DDP sensitivity of NSCLC in vivo.
Discussion
Currently, DDP is an effective chemotherapy drug used to treat human malignancies, but the efficacy is frequently limited due to the development of chemotherapeutic resistance. 37 A great amount of evidence has manifested that circRNAs exert the essential function in the drug resistance of various tumors. For example, circRNA circPAN3, a valuable indicator, was reported to be connected with doxorubicin resistance by interacting with miR-153-5p/miR-183-5p in acute myeloid leukemia. 38 Similarly, circRNA hsa_circ_0035483 could contribute to the resistance of gemcitabine through sponging miR-335 in renal cancer. 39 Moreover, it was previously presented that circ_0076305 was aberrantly increased in DDP-resistant NSCLC. 17 However, the underlying mechanisms of circ_0076305 on the chemoresistance of DDP in NSCLC still needed further research.
In this article, circ_0076305 was identified to be highly expressed in DDP-resistant NSCLC tissues and cells, in accordance with former work. 17 Moreover, P-gp and LRP (a main component of the vault) were reported to play important roles in drugs resistance. 40,41 In this study, circ_0076305 deletion could decrease P-gp and LRP expression, implying that circ_0076305 silencing might reduce drug resistance in NSCLC cells. Apart from that, in vivo, the chemoresistance of circ_0076305 was also verified in NSCLC xenografts in nude mice.
Previous research suggested that circRNAs could exert their effects through the interaction with miRNAs. 42 In this manuscript, through circRNA interactome analysis, they viewed some candidate miRNAs that might be related to circ_0076305, containing miR-186-5p. Moreover, some literature has suggested that miR-186-5p was frequently decreased in a variety of tumors. 22,23 Therefore, miR-186-5p was chosen to further export the mechanism of circ_0076305. Their data verified that miR-186-5p was lowly expressed, and there was a binding relationship between circ_0076305 and miR-186-5p in DDP-resistant NSCLC cells. Functionally, the downregulation of miR-186-5p partly abrogated the circ_0076305 deficiency-induced enhancement in DDP sensitivity. In agreement with earlier studies, the promotion action of miR-186-5p on drug sensitivity was proved in NSCLC. 25
In recent decades, miRNAs could exert the role by regulating target mRNAs. 43 In the article, a target of miR-186-5p, ABCC1, was first identified in NSCLC cells. Moreover, ABCC1 has been pointed out to be related to drug resistance in NSCLC. 32 Furthermore, their data proved that overexpression of ABCC1 overturned the miR-186-5p upregulation-mediated improvement in DDP sensitivity, suggesting that miR-186-5p could regulate the chemoresistance of DDP in NSCLC through targeting ABCC1. In addition, rescue assay demonstrated that miR-186-5p inhibitor could partially reverse the suppression action of circ_0076305 knockdown on the level of ABCC1 in DDP-resistant NSCLC cells, supporting that circ_0076305 could impact ABCC1 expression through sponging miR-186-5p in vitro.
Additionally, some recent studies have presented that circRNAs can be enriched and stabilized in exosomes, which are involved in the progression of various diseases. 44 –46 In this study, their data verified that the circ_0076305 level was increased in exosome from the NSCLC serum, and the exosomal circ_0076305 could enter NSCLC cells. In addition, exosome-based diagnosis allows for timely monitoring of changes in molecular markers during cancer progression, and such blood sample-based tests are easier to monitor and collect than repeated tissue biopsies. Therefore, the authors will focus on whether the regulatory function of exosomal circ_0076305 on DDP resistance was mediated through the miR-186-5p/ABCC1 axis in NSCLC in the subsequent study.
Conclusion
The findings of this study first exhibited that circ_0076305 functioned as ceRNA of miR-186-5p to positively regulate ABCC1 expression, thereby boosting DDP resistance in NSCLC. This study implied an underlying circRNA-targeted therapy for NSCLC patients.
Footnotes
Disclosure Statement
The authors declare that they have no financial conflicts of interest.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.