
Abstract
Background:
Acute myeloid leukemia (AML) is the most common acute leukemia in adults. Circular RNAs (circRNAs) participate in the deterioration of many hominine cancers, including AML. In this study, the authors investigated the role and potential mechanism of circ_0058058 in AML progression.
Methods:
The expression of circ_0058058, microRNA-4319 (miR-4319), and eukaryotic initiation factor 5A2 (EIF5A2) was determined by quantitative real-time polymerase chain reaction. Cell proliferation, apoptosis, migration, and invasion were evaluated by cell counting kit-8 (CCK-8), cell colony formation, flow cytometry, and transwell assay, respectively. Levels of the relative proteins were detected by Western blot. The connection among circ_0058058, miR-4319, and EIF5A2 was verified by dual-luciferase reporter assay.
Results:
Circ_0058058 and EIF5A2 were enhanced, whereas miR-4319 was declined in AML. Circ_0058058 knockdown inhibited cell proliferation, migration, and invasion, and facilitated cell apoptosis by targeting miR-4319 in AML cells. Moreover, as a target of miR-4319, EIF5A2 overexpression overturned the inhibitory effects of miR-4319 upregulation on AML progression. Besides, circ_0058058 sponged miR-4319 to upregulate EIF5A2 expression in AML cells.
Conclusion:
Circ_0058058 knockdown inhibited cell proliferation, migration, and invasion, but accelerated cell apoptosis by reducing EIF5A2 expression by targeting miR-4319, suggesting that circ_0058058 could be a therapeutic target for the treatment of AML.
Introduction
Acute myeloid leukemia (AML) is a disease of the hematological system, which originates from the malignant myeloid hematopoietic stem and progenitor cells. 1,2 AML is s characterized by rapid proliferation and high heterogeneity of immature myeloid cells, resulting in interference of the production of normal blood cells. 3 Despite having achieved prominent improvements in stem cell transplantation and chemotherapy techniques, the survival rate of AML patients remains a concern due to metastasis and recurrence. 4 In 2016, nearly 20,000 new cases were diagnosed with AML in the United States and more than half of these AML patients died. 5 Therefore, it is required to explore the potential molecular mechanisms of AML.
Circular RNAs (circRNAs) are noncoding RNAs with highly stable and conservative covalent closed-loop structures. 6 Previous studies clarified that circRNAs played the pivotal part in a variety of tumor biological behaviors. 7,8 Recently, multiple circRNAs were revealed to modulate the development of AML. For instance, circPAN3 could promote drug resistance by influencing cell autophagy and apoptosis. 9 Ping et al. proved that circ_0009910 knockdown restrained cell growth by sponging miR-20a-5p in AML. 10 In addition, circ_0000370 facilitated cell growth through modulating miR-1299 and S100A7A in AML. 11 Nevertheless, the role and molecular mechanism of circ_0058058 in AML progression remain dim.
The role of microRNAs (miRNAs) in the progression of AML has received extensive attention, such as miR-125a, 12 miR-29b, 13 and miR-193a. 14 MicroRNA-4319 (MiR-4319), as a tumor suppressor, was involved in the development of many cancers. For instance, miR-4319 expression was drastically degraded in prostate cancer, 15 colorectal cancer, 16 and non-small cell lung cancer, 17 and miR-4319 facilitation restrained tumor cell growth, migration, and invasion. However, the investigation of miR-4319 in AML is limited.
Eukaryotic initiation factor 5A2 (EIF5A2) is a member of EIF family. Recently, EIF5A2, located on the chromosome 3q25-q27, was a gene region closely related to tumor. 18 Moreover, EIF5A2 was highly expressed in renal cancer, 19 gallbladder cancer, 20 and prostate cancer. 21 However, the role of EIF5A2 in the progression of AML is unknown.
In this study, the authors analyzed the data of GSE94591 dataset, and circ_0058058 was identified as a differentially expressed circRNAs in AML. Then, the functional role of circ_0058058 in AML progression was investigated, and the interaction among circ_0058058, miR-4319, and EIF5A2 in AML cells was verified.
Materials and Methods
AML samples and cell culture
Blood samples were recruited from 35 AML patients and 35 healthy volunteers at Wuhan NO.1 Hospital. Serums were obtained by centrifugation of the blood samples at 5000 rpm for 5 min. All participants provided the written informed consent forms. The procedure in the authors' research was authorized by the Ethics Committee of Wuhan NO.1 Hospital.
AML cells (KG-1 and HL-60) and human bone marrow stroma cells (HS-5) were bought from BeNa Culture Collection (Beijing, China). Roswell Park Memorial Institute 1640 (RPMI 1640; Hyclone, South Logan, UT) with 10% fetal bovine serum (FBS) was utilized to culture the cells at 37°C with 5% CO2.
Transfection
Small interfering RNA against circ_0058058 (si-circ_0058058#1, si-circ_0058058#2, or si-circ_0058058#3) and the control (si-NC), miR-4319 mimic (miR-4319), miR-4319 inhibitor (anti-miR-4319), and the corresponding controls (miR-NC and anti-miR-NC), and EIF5A2 overexpression plasmid (pcDNA-EIF5A2) and its control (pcDNA-NC) (GenePharma Shanghai, China) were transfected into KG-1 and HL-60 cells by Lipofectamine 3000 (Invitrogen, Carlsbad, CA).
Quantitative real-time polymerase chain reaction
Total RNA was extracted from AML samples and cells by TRIzol (Invitrogen). The PrimeScript™ strand cDNA synthesis kit (Takara, Dalian, China) was employed to reverse transcript complementary DNA (cDNA). Relative expression of gene was estimated by SYBR Premix Ex Taq (Takara). U6 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were used as the endogenous controls for miR-4319 and circ_0058058 or EIF5A2. The relative gene expression was calculated using 2−ΔΔCt method. Primers were listed as follows: circ_0058058, F: 5′-GAGGACTATGTGGTGGTGTCC-3′ and R: 5′-CCTGAGAGCTTTTGCAGTCC-3′; miR-4319, F: 5′-GCACAGCTCCCTGAGCAA-3′ and R: 5′-CAGTGCGTGTCGTGGAGT-3′; EIF5A2, F: 5′-TATGCAGTGCTCGGCCTTG-3′ and R: 5′-TTGGAACATCCATGTTGTGAGTAGA-3′; GAPDH, F: 5′-TGGGCTACACTGAGCACC-3′ and R: 5′-AAGTGGTCGTTGAGGGCAATG-3′; and U6, F: 5′-CAGCACATATACTAAAATTGGAACG-3′ and R: 5′-ACGAATTTGCGTGTCATCC-3′.
Cell proliferation assay
Cell counting kit-8 (CCK-8, Beyotime, Shanghai, China) assay and cell colony formation assay were used to assess cell proliferation. For CCK-8 assay, transfected KG-1 and HL-60 cells were placed into the 96-well plates at 37°C. Zero hour, 24 h, 48 h, or 72 h later, 10 μL CCK-8 solution was added to each well and cultured for another 2 h. The optical density was checked using a microplate reader at 450 nm.
For cell colony formation assay, transfected AML cells were seeded into the six-well plates containing fresh RPMI 1640 at 37°C. After incubation for 2 weeks, 0.1% crystal violet (Solarbio, Beijing, China) was used to stain cells. Finally, the number of colonies was counted using a microscope.
Cell apoptosis analysis
The apoptosis of transfected KG-1 and HL-60 cells was detected using an Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) detection kit (Keygen, Beijing, China) by flow cytometry. After 48 h of transfection, AML cells were collected and incubated with 5 μL Annexin V-FITC and 5 μL PI for 20 min in the dark. Cell apoptosis was evaluated by the flow cytometer (BD Biosciences, Hercules, CA).
Transwell assay
Transwell assay with chambers was applied for cell migration and invasion analysis. For cell invasion, the chambers were precoated with Matrigel (BD Biosciences). AML cells were transfected and resuspended in serum-free medium, and then seeded into the upper chambers, while the lower chambers were filled with complete medium. For cell migration, the chambers were noncoated with Matrigel (BD Biosciences). Finally, the migratory or invasive cells were stained with 0.1% crystal violet (Solarbio) after culture for 24 h. The migrated and invaded cells were counted using a microscope.
Western blot
RIPA buffer with a protease inhibitor cocktail (Beyotime) was used to collect and lyse AML samples and cells. The extracted proteins were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred to polyvinylidene fluoride (PVDF; Beyotime) membranes followed by blocking with 5% skim milk for 2 h. Then, the primary antibodies against B cell lymphoma-2 (Bcl-2; Abcam, Cambridge, MA), cyclinD1 (1:200; Abcam), cleaved caspase-3 (1:500; Abcam), EIF5A2 (1:1000, Abcam), and GAPDH (1:5000; Abcam) were used to incubate the above membranes at 4°C overnight. The membranes were probed with horseradish peroxidase-conjugated secondary antibody (1:3000; Abcam) for 1 h. Finally, the signals were evaluated using an ECL reagent (Beyotime).
Dual-luciferase reporter assay
The wild-type (WT) fragments of circ_0058058 or EIF5A2 possessing the predicted miR-4319 binding site, and the mutated-type fragments of circ_0058058 or EIF5A2 were cloned into the pmirGLO vector (Promega Corporation, Madison, WI), named as circ_0058058-WT, circ_0058058-MUT, EIF5A2 3′UTR-WT, and EIF5A2 3′UTR-MUT, respectively. These vectors were co-transfected into AML cells with miR-4319 or miR-NC. After 48 h, the luciferase activity was examined by a dual-luciferase assay kit (AmyJet Scientific, Wuhan, China).
Statistical analysis
Data were exhibited as mean ± standard deviation (SD) and analyzed using GraphPad Prism 6.0. The correlation among circ_0058058, miR-4319, and EIF5A2 was analyzed by Pearson correlation analysis. The difference between two or multiple groups was evaluated by χ 2 test or one-way analysis of variance (ANOVA). p-Value <0.05 was considered significant difference.
Results
Circ_0058058 was highly expressed in AML patient samples and cells
To screen the differentially expressed circRNAs in AML patients, the authors analyzed the circRNA microassay dataset GSE94591. As shown in Figure 1A, hierarchical clustering manifested 10 upregulated circRNAs in AML patients compared to healthy control groups. Among these circRNAs, circ_0058058 was significantly increased in AML patient samples. They also detected circ_0058058 expression in 35 AML patients and 35 healthy controls and found that circ_0058058 expression was enhanced in AML patients compared with healthy controls (Fig. 1B). Moreover, circ_0058058 was distinctly increased in AML cell lines (KG-1 and HL-60) compared with bone marrow stroma cells (HS-5) (Fig. 1C). To further verify the circular form of circ_0058058, RNase R treatment was performed in KG-1 and HL-60 cells. The results showed that circ_0058058 was resistant to RNase R digestion (Fig. 1D–E), suggesting that circ_0058058 was indeed circRNA. These data manifested that circ_0058058 could serve as a critical regulator in AML progression.
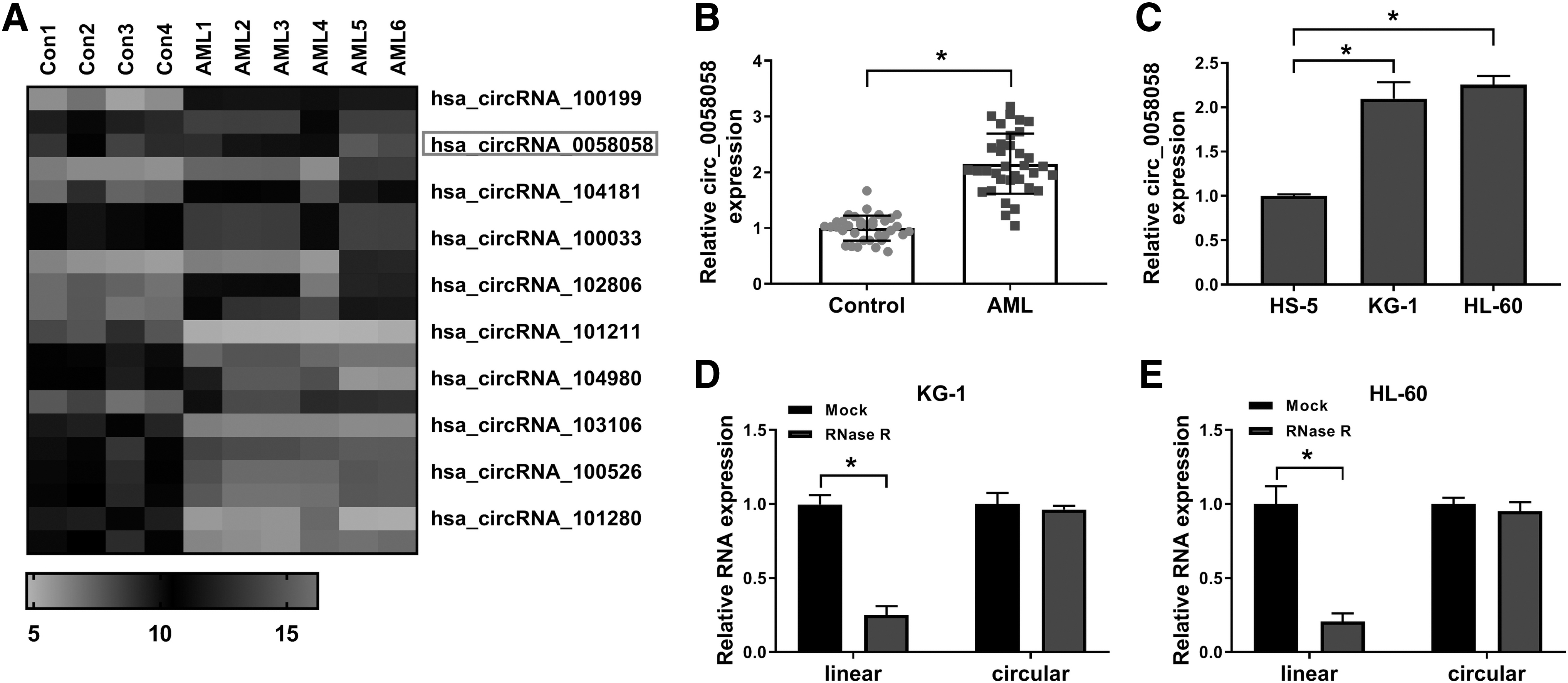
Circ_0058058 was highly expressed in AML patient samples and cells.
Circ_0058058 knockdown inhibited AML cell proliferation, migration, and invasion, and promoted cell apoptosis
To identify the effects of circ_0058058 on AML cells, the authors silenced circ_0058058 expression in KG-1 and HL-60 cells by transfection of siRNAs (si-circ_0058058#1, si-circ_0058058#2, and si-circ_0058058#3) (Fig. 2A, B), and si-circ_0058058#3 was selected for the subsequent experiments. CCK-8 assay showed that circ_0058058 inhibition impaired the proliferation ability of KG-1 and HL-60 cells (Fig. 2C, D). Cell colony formation assay revealed that knockdown of circ_0058058 resulted in an evident reduction of the colonies of KG-1 and HL-60 cells (Fig. 2E). Moreover, downregulation of circ_0058058 accelerated the apoptosis of the two AML cells (Fig. 2F). Transwell assay manifested that circ_0058058 knockdown significantly repressed the ability of migration and invasion in KG-1 and HL-60 cells (Fig. 2G, H). In addition, the protein expression of Bcl-2 and cyclinD1 was decreased in KG-1 and HL-60 cells transfected with si-circ_0058058#3, while cleaved caspase-3 protein expression was increased (Fig. 2I, J). These results indicated that circ_0058058 knockdown could suppress the progression of AML.
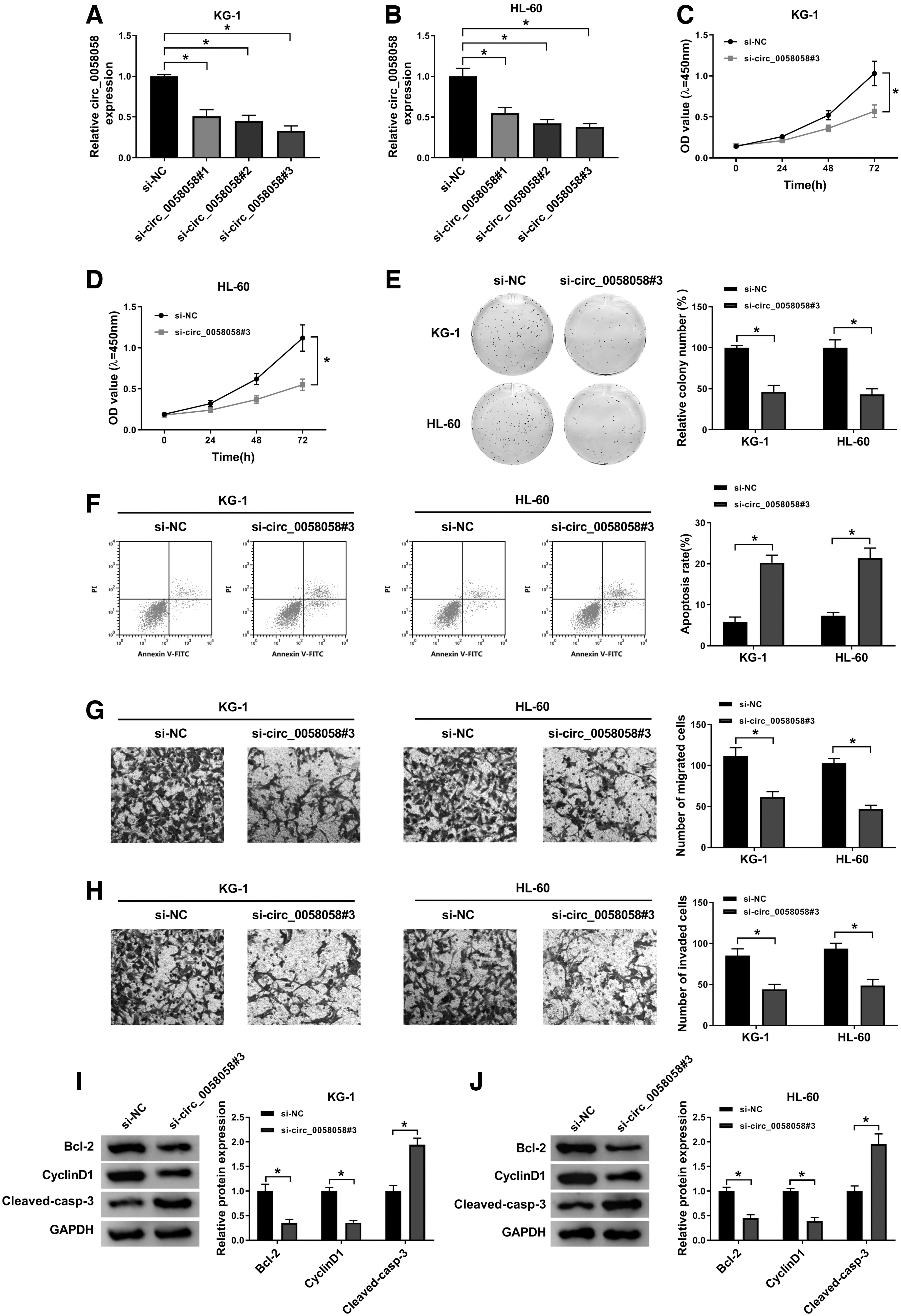
Circ_0058058 knockdown inhibited AML cell proliferation, migration, and invasion, and promoted cell apoptosis.
Circ_0058058 directly interacted with miR-4319 in AML cells
To explore the underlying mechanism of circ_0058058 in AML cells, the downstream target miRNAs of circ_0058058 were searched. The binding sites between circ_0058058 and miR-4319 were predicted by StarBase v2.0 (Fig. 3A). Dual-luciferase reporter assay showed that miR-4319 suppressed the luciferase activity in circ_0058058-WT group, but the luciferase activity of circ_0058058-MUT group had no visible difference in KG-1 and HL-60 cells (Fig. 3B, C). Besides, miR-4319 was inhibited in AML patient samples compared with the normal serum samples (Fig. 3D). The authors also demonstrated that miR-4319 expression was degraded in both KG-1 and HL-60 cells compared with HS-5 cells (Fig. 3E). They then explored the influence of circ_0058058 on the abundance of miR-4319 and observed that knockdown of circ_0058058 upregulated the expression of miR-4319 in KG-1 and HL-60 cells (Fig. 3F). Overall, circ_0058058 targeted miR-4319 and negatively regulated miR-4319 expression in AML cells.
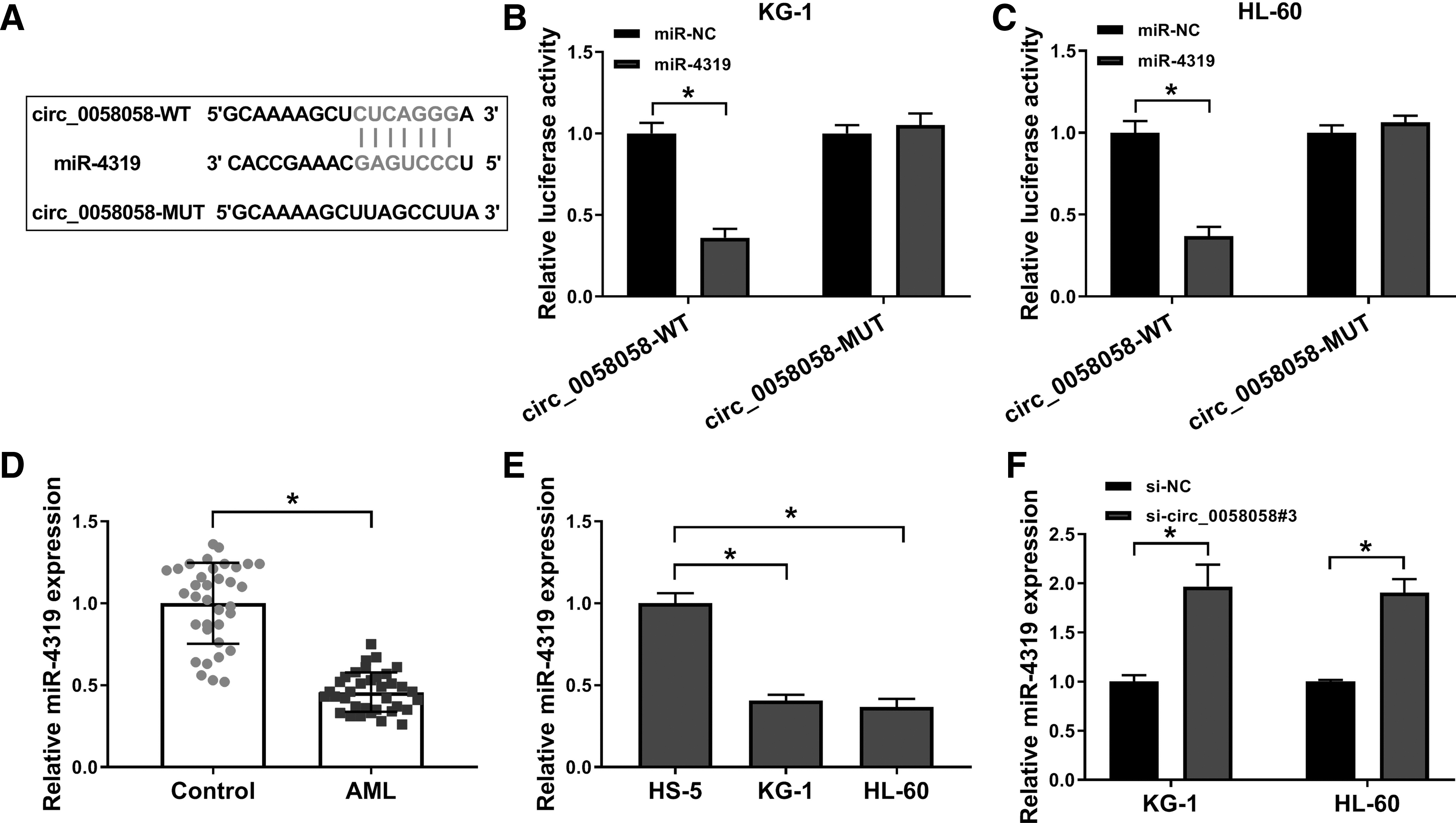
Circ_0058058 directly interacted with miR-4319 in AML cells.
Inhibition of miR-4319 reversed the inhibitory effect of circ_0058058 knockdown on AML progression
To explore the mechanism of circ_0058058 and miR-4319 in AML cells, si-circ_0058058#3 and anti-miR-4319 were co-transfected into KG-1 and HL-60 cells. As exhibited in Figure 4A, miR-4319 inhibition attenuated the promotion effect of circ_0058058 knockdown on miR-4319 expression. Then, the authors found that miR-4319 inhibitor could reverse the inhibition of si-circ_0058058#3 on cell proliferation in KG-1 and HL-60 cells (Fig. 4B–D). The apoptosis rate was promoted by circ_0058058 knockdown, while this effect was weakened by co-transfection of anti-miR-4319 in the two AML cells (Fig. 4E). Similarly, transwell assay elucidated that the number of migration and invasion cells was reduced in si-circ_0058058#3-transfected KG-1 and HL-60 cells, which was inversely augmented by miR-4319 inhibitor (Fig. 4F, G). Meanwhile, the effects of circ_0058058 knockdown on the levels of Bcl-2, cyclinD1, and cleaved caspase-3 could be neutralized by miR-4319 depletion in KG-1 and HL-60 cells (Fig. 4H, I). These above results revealed that circ_0058058 knockdown repressed cell proliferation, migration, and invasion, but promoted cell apoptosis in AML cells by increasing the expression of miR-4319.
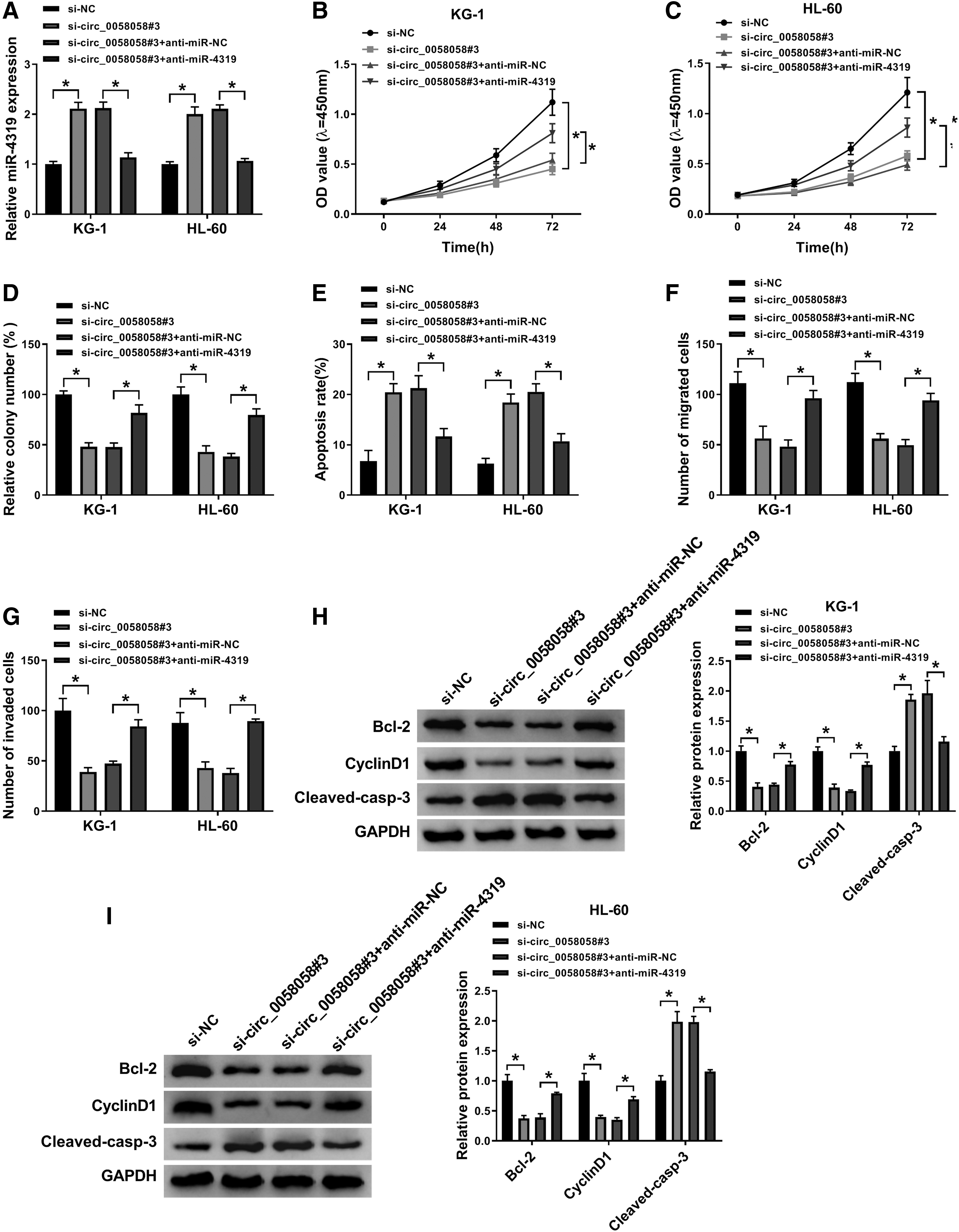
Inhibition of miR-4319 overturned the suppressive effect of circ_0058058 knockdown on AML progression. KG-1 and HL-60 cells were transfected with si-NC, si-circ_0058058#3, si-circ_0058058#3 + anti-miR-NC, or si-circ_0058058#3 + anti-miR-4319.
EIF5A2 was a target of miR-4319
The authors then screened the possible targets of miR-4319 by using StarBase v2.0 bioinformatics tool. As displayed in Figure 5A, there were complementary binding sites between miR-4319 and the 3′UTR of EIF5A2. To verify this prediction, dual-luciferase reporter assay was performed. The results manifested that the luciferase activity of EIF5A2 3′UTR-WT was prominently declined in KG-1 and HL-60 cells transfected with miR-4319, while the luciferase activity of EIF5A2 3′UTR-MUT had no significant change (Fig. 5B, C). The levels of EIF5A2 in AML patient samples and cells were elevated relative to the normal samples and cells (Fig. 5D–G). In addition, miR-4319 elevation significantly dwindled the mRNA and protein levels of EIF5A2 in KG-1 and HL-60 cells (Fig. 5H, I). Collectively, these data illuminated that miR-4319 could target EIF5A2 and negatively modulate EIF5A2 expression in AML cells.
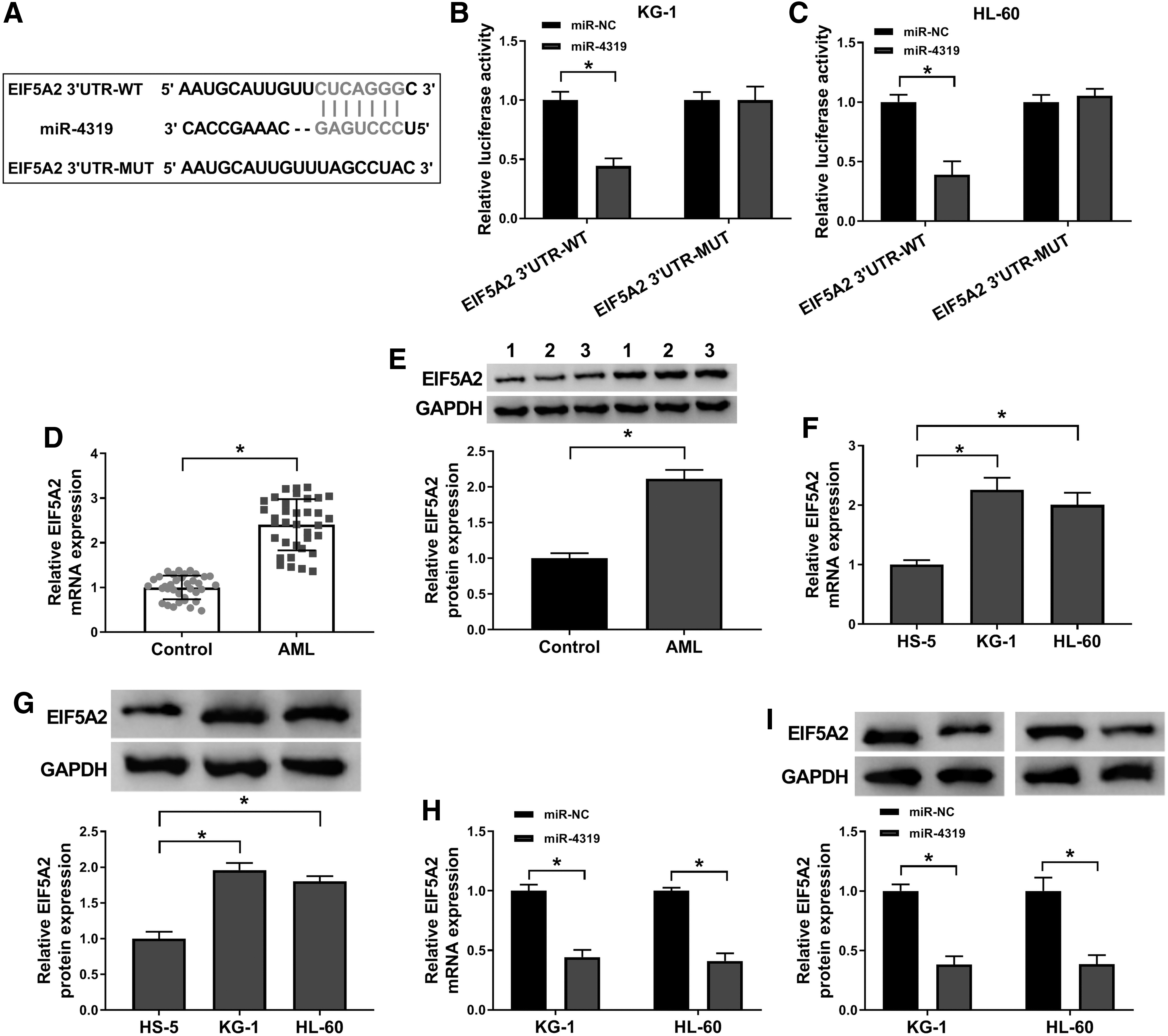
EIF5A2 was a target of miR-4319.
Overexpression of miR-4319 inhibited the progression of AML by targeting EIF5A2
To determine whether EIF5A2 could participate in the regulation of miR-4319 in AML progression, miR-4319 and pcDNA-EIF5A2 were co-transfected into AML cells. MiR-4319 overexpression downregulated EIF5A2 expression at mRNA and protein levels, while transfection of pcDNA-EIF5A2 promoted EIF5A2 expression in KG-1 and HL-60 cells (Fig. 6A–C). Moreover, overexpression of miR-4319 restrained cell proliferation (Fig. 6D–E), the number of colonies (Fig. 6F), migration (Fig. 6H), and invasion (Fig. 6I), but facilitated cell apoptosis (Fig. 6G) in KG-1 and HL-60 cells, while these effects could be blocked by overexpressing EIF5A2. Furthermore, upregulation of miR-4319 reduced Bcl-2 and cyclinD1 protein levels, and induced cleaved caspase-3 expression in KG-1 and HL-60 cells, which were reversed by EIF5A2 upregulation (Fig. 6J–K). These data indicated that miR-4319 could modulate AML progression by regulating EIF5A2.

Overexpression of miR-4319 inhibited the progression of AML by targeting EIF5A2. KG-1 and HL-60 cells were transfected with miR-NC, miR-4319, miR-4319 + pcDNA-NC, or miR-4319 + pcDNA-EIF5A2.
Circ_0058058 acted as a miR-4319 sponge to regulate EIF5A2 expression in AML
The correlation among circ_0058058, miR-4319, and EIF5A2 was evaluated. As presented in Figure 7A, B, silencing circ_0058058 markedly degraded EIF5A2 expression at mRNA and protein levels, while miR-4319 inhibitor could promote EIF5A2 expression in circ_0058058#3-transfected KG-1 and HL-60 cells. More importantly, Pearson correlation analysis results showed that circ_0058058 or EIF5A2 expression was negatively correlated with miR-4319 expression in AML samples (Fig. 7C, D), while circ_0058058 expression was positively correlated with EIF5A2 expression (Fig. 7E). The authors' findings suggested that circ_0058058 modulated EIF5A2 expression by sponging miR-4319 in AML.

Circ_0058058 acted as a miR-4319 sponge to regulate EIF5A2 expression in AML.
Discussion
Increasing evidence has revealed that the aberrant expression of circRNAs was implicated in various cancers, including AML. 22,23 Importantly, the functions of circRNAs in AML are largely mediated by targeting miRNAs to release the mRNAs. 24 For instance, circRNA-100290 boosted AML cell growth and repressed cell apoptosis by targeting miR-203. 25 Yuan et al. reported that circ-0004136 induced the proliferation of AML cells by serving as a sponge for miR-142. 26 These data hinted that circRNAs might be the potential targets for the treatment of AML. In this research, GSE9459 dataset showed that circ_0058058 was identified as a novel circRNA that might be related to AML. A previous study reported that circ_0058058 was upregulated in AML. 27 Consistently, the authors found that circ_0058058 was evidently enhanced in AML samples and cells. Furthermore, the authors' data revealed that circ_0058058 deficiency retarded cell proliferation, migration, and invasion, and elevated cell apoptosis in AML cells, suggesting that circ_0058058 functioned as an oncogene in the development of AML.
Accumulating evidence indicated that circRNAs played a biological role by releasing the expression of miRNA-targeted genes as a miRNA sponge. 28 For example, circ_0009910 deletion repressed cell growth in AML by targeting miR-20a-5p. 29 To figure out the downstream gene of circ_0058058 in AML cells, the authors then screened the miRNAs that contained the binding sites of circ_0058058. The authors' data showed that miR-4319 was a target of circ_0058058. MiR-4319 was previously identified to play an anticancer role in gastric cancer 30 and thyroid cancer. 31 In this study, the authors revealed that miR-4319 was distinctly degraded in AML, which was in agreement with the previous reports. 32 More importantly, they observed that miR-4319 inhibitor could weaken the inhibition effects of circ_0058058 depletion on the progression of AML, implying that miR-4319 served as an inhibitor in AML. Taken together, these data supported that circ_0058058 boosted AML progression by targeting miR-4319.
The authors' data manifested that EIF5A2 was a target of miR-4319. EIF5A2 was proved to be upregulated in various cancers, including AML. 33,34 Consistently, they found that EIF5A2 was significantly increased in AML samples and cells. Moreover, Liu et al. reported that EIF5A2, as a target of miR-9, played a carcinogenic role in AML. 34 In this research, EIF5A2 elevation overturned the inhibition effects of miR-4319 overexpression on the progression of AML, suggesting that miR-4319 played an anticancer role by inhibiting the expression of EIF5A2. According to all findings in this study, the authors hypothesized that there was a circRNA-miRNA-mRNA regulatory axis in AML progression. As expected, the authors' data showed that circ_0058058 knockdown notably reduced EIF5A2 expression by sponging miR-4319. Overall, circ_0058058 was involved in the progression of AML by regulating miR-4319/EIF5A2 axis. However, this study has a limitation that the authors did not perform an in vivo experiment, and further studies are needed to determine the role of circ_0058058 in the pathogenesis of AML in mice.
In conclusion, circ_0058058 facilitated the progression of AML by sponging miR-4319 to regulate EIF5A2 expression, providing the potential therapeutic targets for the treatment of AML patients.
Footnotes
Acknowledgment
The authors sincerely appreciate all members who participated in this study.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work approved by Chengdu Municipal Health Commission Project “Toll-like receptor signaling pathway in the co-infection between influenza A virus and Streptococcus pneumoniae” (No: 2019107).