
Abstract
Objective:
To explore the role of miR-498 in the radiotherapy resistance of esophageal cancer (EC) and its underlying mechanism.
Methods:
In vivo models of EC tissues with radioresistance or radiosensitivity were isolated from 72 EC patients who received radiotherapy. In vitro models were established after irradiation of KYSE30 cells. Quantitative reverse transcription polymerase chain reaction (qRT-PCR) and Western blot were employed to measure the expression levels of miR-498 and DNMT3b in EC cells sensitive or resistant to irradiation. Then, protein expression of DNMT3b was verified by immunohistochemistry. The cell viability, colony formation rate, and cell apoptotic rate of EC were correspondingly assessed by CCK-8, colony formation assay, and Annexin V/PI (propidium iodide) double staining. Western blot was utilized to perform the expression levels of PI3K, p-PI3K, AKT, and p-AKT in EC cell lines after irradiation.
Results:
Highly expressed DNMT3b and lowly expressed miR-498 were found in EC tissues. EC tissues with radiosensitivity had higher miR-498 level and lower DNMT3b expression than EC tissues with radioresistance. Overexpression of miR-498 or knockdown of DNMT3b enhanced the radiosensitivity of EC cells. DNMT3b was a target gene of miR-498. DNMT3b diminished the radiosensitization of miR-498 in EC cells.
Conclusions:
MiR-498 enhances the sensitivity of EC cells to radiation by DNMT3b inhibition, and exerts biological functions by inactivating the PI3K/AKT signaling pathway.
Introduction
To date, esophageal cancer (EC) has emerged as one of the prominent reasons for cancer-related fatality worldwide. 1 The esophageal adenocarcinoma and esophageal squamous cell carcinoma are deemed as the two primary subtypes of EC, which occupy ≥95% of esophageal malignancies. 2 Patients with operable EC are frequently intervened by surgical resection, but the prevalence of recurrence rate exists after surgical resection, which encourages the advancement of multimodality therapies that combine esophageal resection with radiotherapy, chemotherapy, and chemoradiotherapy. 3,4 Radiotherapy is one of the major therapy modalities for patients with EC, nevertheless, distant metastasis and local recurrence may herald radiation therapy failure in EC caused by radioresistance. 5,6 Thus, the exploration of effective sensitizers to augment tumor response to radiation is urgently necessary for EC patients. Besides, understanding the mechanisms of radioresistance is also essential to perfect strategies of radiation therapy.
MicroRNAs (miRNAs) are small noncoding RNAs of 19–23 nucleotides that can interact with multiple messenger RNAs (mRNAs) for negative mediation of gene expression, bringing about mRNA degradation or mRNA translation suppression. 7 Some studies have proposed that miRNAs directly affect radioresistance through modifying specific manners, including cell proliferation, migration, 8 and apoptosis. 9 As an miRNA few researched in EC, miR-498 can be mediated by long noncoding RNA TUG1 to interfere with EC cell proliferation and invasion. 10 However, the specific functional mechanism of miR-498 in EC and whether miR-498 is a key regulator for the sensitivity of EC cells to radiotherapy remain to be further investigated. DNA methyltransferases (DNMTs) are a class of enzymes processing DNA methylation, which are divided into three types: DNMT3a/3b/3l, DNMT1, and DNMT2. 11 DNMT3b is the chief de novo DNMT expressed and active during the early stage of embryonic development. 12 Interestingly, X-ray irradiation can induce demethylation of the whole genome through DNMT repression. 13 Moreover, knockdown of DNMT3b has been proven to sensitize esophageal and prostate cancer cells to radiation. 14,15 In this study, it was predicted that DNMT3b was a target gene of miR-498. However, the relationship between miR-498 and DNMT3b in the EC therapy needs to be elucidated.
The PI3K/AKT signaling pathway interferes with the mediation of cell growth, metabolism, and proliferation, and dysregulation of PI3K/AKT pathway components is responsible for ∼30% human cancers. 16 In this study, the profile of miR-498 in vitro and in vivo was assessed. Its potential utility in sensitivity and resistance for radiotherapy in EC was also focused. It was proposed that miR-498 potentiates cell apoptosis, represses cell proliferation, and enhances radiosensitivity for EC cells by directly targeting DNMT3B through the PI3K/AKT pathway.
Materials and Methods
Ethical statement
The study was carried out according to the Declaration of Helsinki. The study protocol concerning human was approved by the Second Affiliated Hospital of Wenzhou Medical University. All the patients provided their written informed consent.
Subjects
A total of 72 EC patients who received radiotherapy were recruited consecutively from the Department of Gastroenterology in the Second Affiliated Hospital of Wenzhou Medical University since March 2016 to March 2019. Esophageal tumor tissues (EC group) and paracarcinoma tissues (Normal group) were collected and stored in liquid nitrogen. The patient underwent enhanced chest computed tomographic (CT) and barium esophagogram 1 month after radiotherapy. The 22 cases with radiotherapy resistance and 50 cases with radiotherapy sensitivity were selected after evaluation of radiotherapy efficacy according to X-ray evaluation criteria. Inclusion criteria were (1) treatment-naive patients who are diagnosed with EC by pathological confirmation, X-ray image, and CT scan and (2) patients who have accepted and completed the standard radical radiotherapy according to the treatment standard of Radiotherapy Department of Peking University International Hospital. The therapy scheme is 1.8–2.0 Gy/time, 1 time/d and 5 times/week, and the total radiotherapy dose is 60–70 Gy. Exclusion criteria were (1) patients who do not agree with the collection of specimens, (2) patients who are intolerant to surgery, and (3) patients with immune system diseases.
Immunohistochemistry
Immunohistochemical staining was employed to determine the expression of DNMT3b in EC. In brief, sections were subjected to xylene for deparaffinization and hydration, and then orderly underwent gradient ethanol dehydration. Antigen retrieval was carried out by immersing sections in 10 mmol/L sodium citrate solution (pH 6.0) for 10 min of heating at 100°C and 20 min of cooling, which was repeated for three times. The sections were washed in phosphate-buffered saline (PBS, pH 7.2) and blocked in 3% hydrogen peroxide and goat serum at room temperature for 15 min to avoid unspecific response. After that, sections were incubated with monoclonal antibody against DNMT3b (ab16049, 1:300; Abcam, Cambridge, MA) overnight at 4°C, and subsequently incubated with biotin-labeled secondary antibody at 37°C for 30 min. Before color development with DAB, sections were given to avidin conjugated to horseradish peroxidase (avidin-HRP) at 37°C for 20 min of incubation. After that, sections were rinsed with running water for 15 min to terminate the color reaction. Sections were counterstained with 1% hematoxylin, and then dehydrated and sealed.
Cell line
The mucosal epithelial cells (human esophageal epithelial cell [HEEC] group) were isolated from normal mucosal tissues adjacent to EC tissues. EC cell lines (KYSE150, KYSE30, KYSE510, and KYSE410) were purchased from the Cell Resource Center of Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences. The EC cell lines were incubated in Dulbecco's modified eagle medium or RPMI-1640 medium (Gibco) containing 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 U/mL streptomycin at 37°C with 5% CO2.
Construction of cell lines resistant to radiotherapy
KYSE30 cells in the logarithmic growth phase were irradiated at 2 Gy by a 6 MV linear accelerator. Immediately after irradiation, cells were aspirated to a new RPMI-1640 medium to observe cell viability. The active cells were irradiated at increased radiation dose and subcultured until they could survive stably under the dose of 6 Gy. After that, radioresistant EC cell line KYSE30R was obtained for subsequent experiments.
Cell transfection
Cells (KYSE140 and KYSE140R) were transfected with miR-498 mimic, mimic NC, si-DNMT3b, si-NC, miR-498 mimic+OE-DNMT3b, and miR-498 mimic+OE-NC, respectively, employing the Lipofectamine 2000 transfection reagent (Invitrogen, Inc.). On the first day after transfection, cells were seeded onto a standard six-well cell culture plate at a density of 1 × 106 cells/well. Then cells in each well (cell density: 50%–70%) received 1.5 mL medium without antibiotics. The above plasmids were synthesized and constructed by Ruibao Biological Co., Ltd. (Guangzhou, China).
CCK-8
After transfection for 24 h, KYSE30 and KYSE30R cells were digested with 0.25% trypsin, followed by resuspension and cell counting. After cells were diluted to 20 cells/μL, 100 μL of cell suspension was aspirated to a 96-well plate (2 × 103 cells/well). Cells were irradiated at 6 Gy after adherence, and the cell proliferation rate was measured at 0, 12, 24, and 48 h after irradiation. Cells in each well were incubated with 10μL CCK-8 solution for 3 h before optical density (OD) at the wavelength of 450 nm was measured using a microplate absorbance reader. OD620nm was reckoned as reference wavelength. Cell survival rate = [OD (Irradiation group) − OD (Blank group)]/[OD (Control group) − OD (Blank group)] × 100%. The experiment was repeated three times.
Colony formation assay
Cells were digested with 0.25% trypsin, before centrifugation and cell counting. Transfected KYSE30 and KYSE30R cells were seeded onto a six-well plate that were pretreated with a medium containing 10% FBS, and then cells in the plates were incubated overnight. Cells were grouped into 0 Gy group (500 cells/well), 2 Gy group (1000 cells/well), 4 Gy group (2000 cells/well), 6 Gy group (3000 cells/well), and 8 Gy group (4000 cells/well). Three wells were set for each group. After that, cells correspondingly received X-ray irradiation at 0, 2, 4, 6, and 8 Gy and were incubated for 10 d. The culture was terminated once the cloned cell in the plate was visible to the naked eye (∼50 cells were counted under a microscope). Then cells were fixed in 4% paraformaldehyde (5 mL) for 15 min and stained with crystal violet solution (1 mL) for 15 min before being observed. The experiment was repeated three times.
Annexin V/propidium iodide double staining
Cells (3 × 105) were seeded into a six-well plate and exposed to X-ray irradiation at 0 or 6 Gy for 24 h when cells reached 50%–60% confluence. Single cell suspension with a density of 1 × 106 cells/mL was obtained and incubated with 5 μL of Alexa Fluor 488 annexin V and 1 μL of 100 μg/mL propidium iodide (PI) at room temperature for 15 min. After that, cells received 400 μL of 1 × Annexin V Binding Buffer and were placed on ice in darkness. The cell apoptotic rate was analyzed by flow cytometer (Beckman Coulter, Inc., Fullerton, CA). The experiment was conducted thrice.
Quantitative reverse transcription polymerase chain reaction (qRT-PCR)
The total RNA was extracted from cells and tissues by utilizing TRIzol (Takara Co., Ltd., Dalian, China), and then the concentration and purity of RNA were determined. The complementary DNA (cDNA) template was synthesized by TaKaRa reverse transcription reaction (Thermo Fisher Scientific). The reaction conditions were consistent with the instructions of the TaKaRa RT-qPCR kit. The test was done in triplicate. GAPDH was regarded as internal reference for polymerase chain reaction amplification, and the 2−ΔΔCt method was employed for data analysis. All operations were conducted following the manufacturer's instructions. All primers were synthesized by Ribo Biotechnology Co., Ltd. (Guangzhou, China), and the amplified primer sequences of each gene are presented in Table 1.
Primer Sequence Information
F, forward primer; R, reversed primer.
Western blot analysis
The proteins were obtained from cells and tissues in each group, and the protein concentration was measured by a BCA kit. The proteins were treated with 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis before membrane transference (300 mA, constant current). After 1 h of inactivation in 5% nonfat dried milk, the membranes were incubated with the primary antibodies against DNMT3b (ab239893, 1:2000; Abcam), PI3K (ab32089, 1:1000), p-PI3K (ab154598, 1:1000), AKT (ab8805, 1:1000), p-AKT (ab81283, 1:1000), and GAPDH (ab181602, 1:1000) overnight at 4°C. After washing thrice with Tris Buffered Saline Tween (TBST), the membranes were cultured with the HRP marked secondary antibody (Abcam) at 37°C for 1 h. The chemiluminescent reagent was applied for color development after the membranes were washed with TBST. The gray value of the target protein band was analyzed by ImageJ software. The test was done in triplicate.
Dual-luciferase reporter gene assay
StarBase was employed to predict the binding site of miR-498 and DNMT3b. The reporter vectors of mutated type sequence and wild-type sequence (MUT-DNMT3b and WT-DNMT3b) were constructed. Then the vectors were cotransfected with miR-498 mimic or mimic NC into KYSE30 cells for 48 h at 37°C with 5% CO2. Cells were washed in PBS and then lysed by PLB. Cells in 10 μL of supernatant were placed in a 96-well plate, and received 10 μL of Luciferase Assay Reagent II. The luciferase reaction intensity was inspected.
RNA immunoprecipitation assay
The RNA immunoprecipitation (RIP) assay was carried out by using a Magna RIP™ RNA-Binding Protein Immunoprecipitation Kit (Millipore, Billerica, MA). The KYSE30 cells were lysed with RIP buffer before incubation with magnetic beads conjugated with human anti-Ago2 (Millipore) or NC normal mouse anti-IgG (Millipore). Then, cells were incubated with Proteinase K to digest proteins, and the immunoprecipitated RNA was isolated for qRT-PCR analysis.
Statistical analysis
For statistical analysis of the data, SPSS 18.0 (IBM Corp., Armonk, NY) and GraphPad Prism 6.0 (GraphPad Software, Inc.) were utilized. Data are displayed as mean ± standard deviation. t-Test was adopted for the comparison between two groups. Comparisons among multiple groups were analyzed by one-way analysis of variance followed by Bonferroni t-test. The correlation between miR-498/DNMT3b and clinicopathological features was performed using chi-square test. p-Values of significance were p < 0.05, unless indicated otherwise in figure legend.
Results
Highly expressed DNMT3b and lowly expressed miR-498 in EC tissues resistant to radiotherapy
Radiotherapy is a frequently used method for EC, however, the application and development of this therapy is not positive due to radioresistance. To explore the molecular mechanism of radioresistance in EC and a possible radiotherapy sensitization strategy, a total of 72 EC patients who received radiotherapy were recruited, and EC tissues as well as paracarcinoma tissues were isolated for subsequent experiments. The expression levels of miR-498 and DNMT3b in EC tissues and paracarcinoma tissues by qRT-PCR and Western blot were examined. The results manifested that compared with the expression levels in the paracarcinoma tissues, the miR-498 level was lowly expressed and DNMT3b level was highly expressed in EC tissues (p < 0.05, Fig. 1A, B).
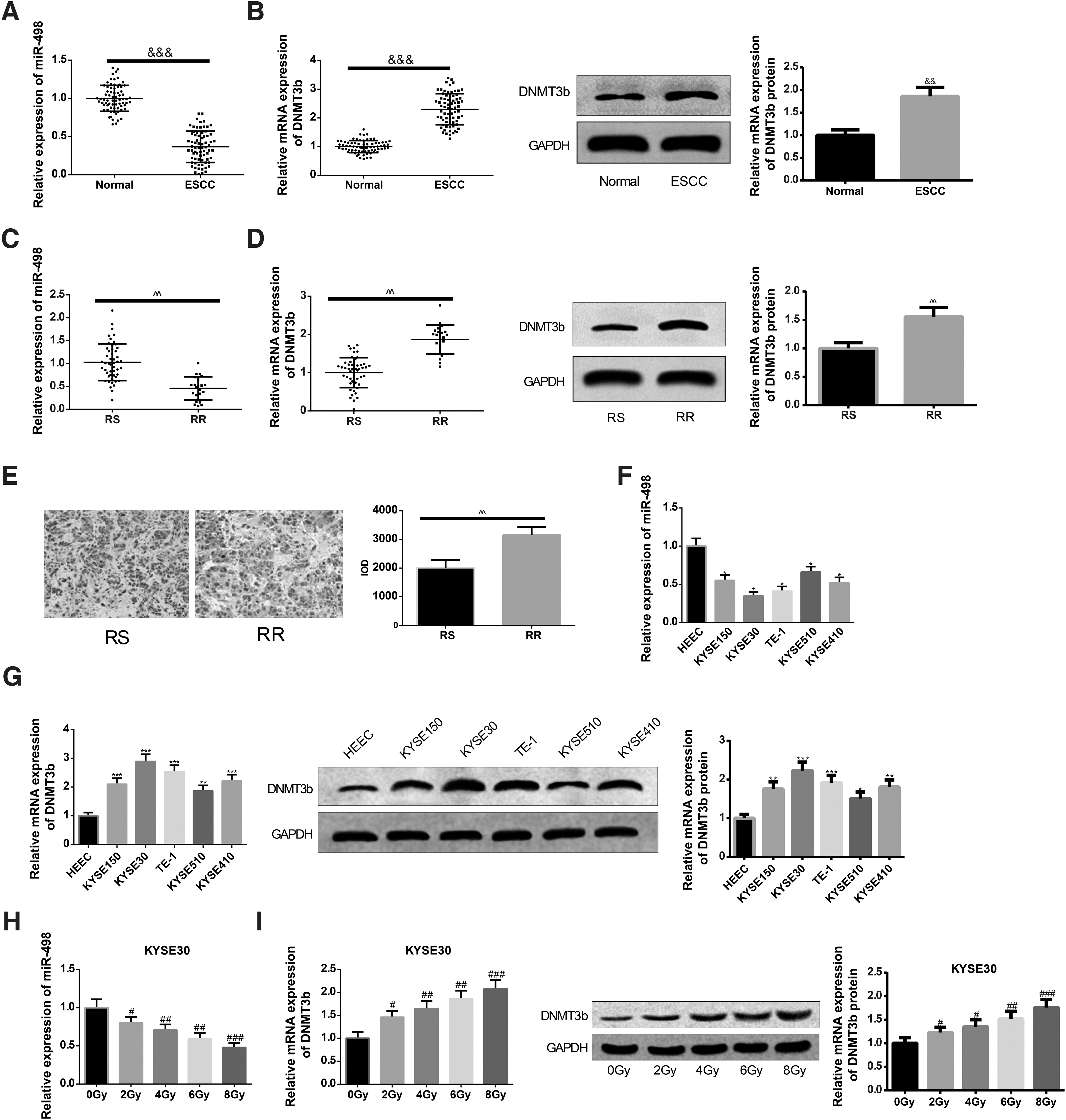
Differentially expressed miR-498 and DNMT3b in EC tissues. The expression levels of miR-498
According to the average relative expression levels of miR-498 and DNMT3b in EC tissues, they were divided into the low-expression group and the high-expression group. In 72 patients with EC, low expression of miR-498 and high expression of DNMT3b are correlated with tumor size, lymph node metastasis, and distant metastasis, but have nothing to do with the age and gender. The relative expression and clinicopathological features of EC patients are presented in Table 2.
The Relative Expression Levels of miR-498 and DNMT3b in Addition to Clinicopathological Features of Esophageal Cancer Patients
The enumeration data are expressed in the form of percentage or rate, and chi-square test was used for the analysis.
mRNA, messenger RNA.
Among 72 patients with EC, 22 patients were resistant to radiotherapy (RR group) and 50 patients were sensitive to radiotherapy (RS group). Then the authors sought to exploit whether miR-498 and DNMT3b were differentially expressed under the radiotherapy sensitivity condition. Toward this end, qRT-RCR and Western blot were applied to evaluate the expression levels of miR-498 and DNMT3b in EC tissues from the RS and RR groups. A pronounced increase in DNMT3b expression and decrease in miR-498 level were found in the RR group as compared with the RS group (p < 0.05, Fig. 1C, D). Subsequently, result of immunohistochemistry on DNMT3b expression in EC tissues sensitive to or resistant to radiation (Fig. 1E) was consistent with the findings obtained from qRT-RCR and Western blot. To further verify these findings, the expression levels of miR-498 and DNMT3b in different EC cell lines were analyzed in vitro. It was found that the KYSE150, KYSE30, KYSE510, and KYSE410 groups had elevated DNMT3b level and suppressed miR-498 expression in comparison with the HEEC group (p < 0.05, Fig. 1F, G). Cell line KYSE30 with more obvious decline in miR-498 expression was selected for subsequent experiments. Result of qRT-PCR unfolded that after irradiation of 0, 2, 4, 6, and 8 Gy, the mRNA expression levels of miR-498 in KYSE30 cells were repressed remarkably with elevation of irradiation dose (p < 0.05, Fig. 1H). Western blot analysis exhibited that after irradiation, the levels of DNMT3b in KYSE30 cells were heightened notably with the increase of irradiation dose (p < 0.05, Fig. 1I).
Upregulation of miR-498 associates with suppressed cell proliferation and colony formation abilities as well as heightened cell apoptotic rate
To probe the effect of miR-498 in EC cell proliferation ability under irradiation condition, KYSE30 and KYSE30R cells were transfected with miR-498 mimic. qRT-PCR measurement on miR-498 expression showed that EC cells transfected with overexpression of miR-498 had elevated expression of miR-498 as compared with cells in mimic NC group (p < 0.05, Fig. 2A). CCK-8 on cell viability yielded that miR-498 upregulation inhibited the proliferation ability of EC cells that are sensitive or resistant to irradiation (p < 0.05, Fig. 2B).

Upregulation of miR-498 enhances the sensitivity of EC cells to radiation. After KYSE30 and KYSE30R cells were transfected with miR-498 mimic, respectively, the miR-498 expression was measured by qRT-PCR
The colony formation assay was utilized to investigate whether enhanced miR-498 expression can increase the sensitivity of EC cells to radiation. The result addressed that overexpression of miR-498 reduced the colony formation rates of parental cell line KYSE30 as well as radioresistant EC cell line KYSE30R under 2, 4, 6, and 8 Gy of X-ray irradiation (p < 0.05, Fig. 2C), and there were significant differences between the miR-498 mimic group and mimic NC group in the colony formation rates under 8 Gy of irradiation (p < 0.05).
Subsequently, Annexin V/PI double staining was employed to analyze the effect of miR-498 on EC cell apoptotic rate after irradiation. The finding exhibited that compared with the mimic NC group, the miR-498 mimic group had elevated apoptotic rate of EC cells that received 6 Gy of irradiation (p < 0.05, Fig. 2D). No remarkable change was noticed between the mimic NC group and miR-498 mimic group in apoptotic rate of EC cells that had not been irradiated (p > 0.05). Collectively, miR-498 hampers proliferation and potentiates apoptosis of EC cells resistant to radiotherapy.
Knockdown of DNMT3b facilitates EC cell apoptosis and restrains cell proliferation and colony formation
The EC cell lines KYSE30 and KYSE30R were transfected with si-DNMT3b, respectively, to perform the role of DNMT3b in EC. Analysis of qRT-PCR and Western blot proposed that knockdown of DNMT3b restrained the mRNA and protein expression levels of DNMT3b (p < 0.05, Fig. 3A, B). Result of CCK-8 illustrated that the cell viability of EC cells sensitive to radiation or resistant to radiation could be suppressed after transfection with si-DNMT3b (p < 0.05, Fig. 3C). The colony formation assay displayed that downregulation of DNMT3b diminished the colony formation rates of EC cells with radiosensitivity or radioresistance, and the difference between the si-NC group and the si-DNMT3b group in cell colony formation rate under 8 Gy of irradiation was obvious (p < 0.05, Fig. 3D).
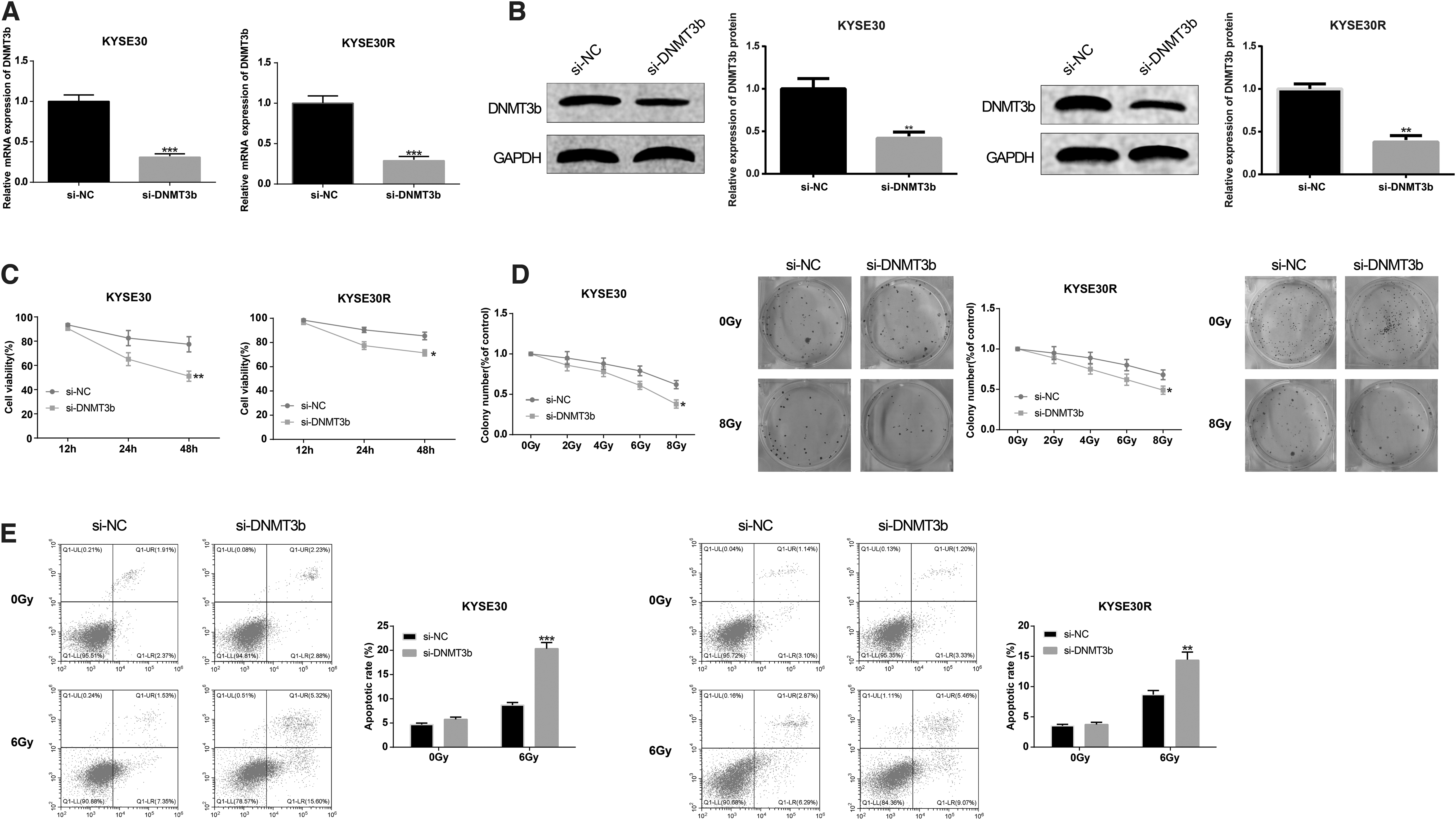
The sensitivity of EC cells to irradiation can be enhanced by DNMT3b suppression. KYSE30 and KYSE30R cells were transfected with si-DNMT3b, respectively. qRT-PCR
Annexin V/PI double staining was applied to explore the role of DNMT3b in EC cell apoptosis after irradiation. The result implicated that knockdown of DNMT3b caused increased apoptotic rate of EC cells that received 6 Gy of irradiation, whereas no significant difference was observed between the si-NC group and the si-DNMT3b group in apoptotic rate of EC cells that had not been irradiated (p > 0.05, Fig. 3E). These data indicated that DNMT3b downregulation enhanced the sensitivity of EC cells to irradiation.
DNMT3b is a target gene of miR-498
The bioinformatics database StarBase was utilized to predict the target gene of miR-498, and then it was found that DNMT3b may be the target gene of miR-498 (Fig. 4A). The mentioned result was verified by dual-luciferase reporter gene assay that cotransfection with WT-DNMT3b and miR-498 mimic had lower relative luciferase activity than cells cotransfected with WT-DNMT3b and mimic NC (p < 0.01, Fig. 4B). However, the relative luciferase activity in cells cotransfected with MUT-DNMT3b and miR-498 mimic was not statistically different from cells cotransfected with MUT-DNMT3b and mimic NC (p > 0.05). The results of RIP assay manifested that the expression of miR-498 or DNMT3b in Ago2 immunoprecipitation of KYSE30 cells was significantly enriched compared with IgG immunoprecipitation of KYSE30 cells (Fig. 4C). Then, these findings were corroborated in KYSE30R cells (Fig. 4D, E). Besides, analysis of qRT-PCR and Western blot exhibited that overexpression of miR-498 potentiated expression of miR-498, whereas restrained DNMT3b level in KYSE30 and KYSE30R cells (p < 0.05, Fig. 4F–I, miR-498 group vs. mimic NC group).
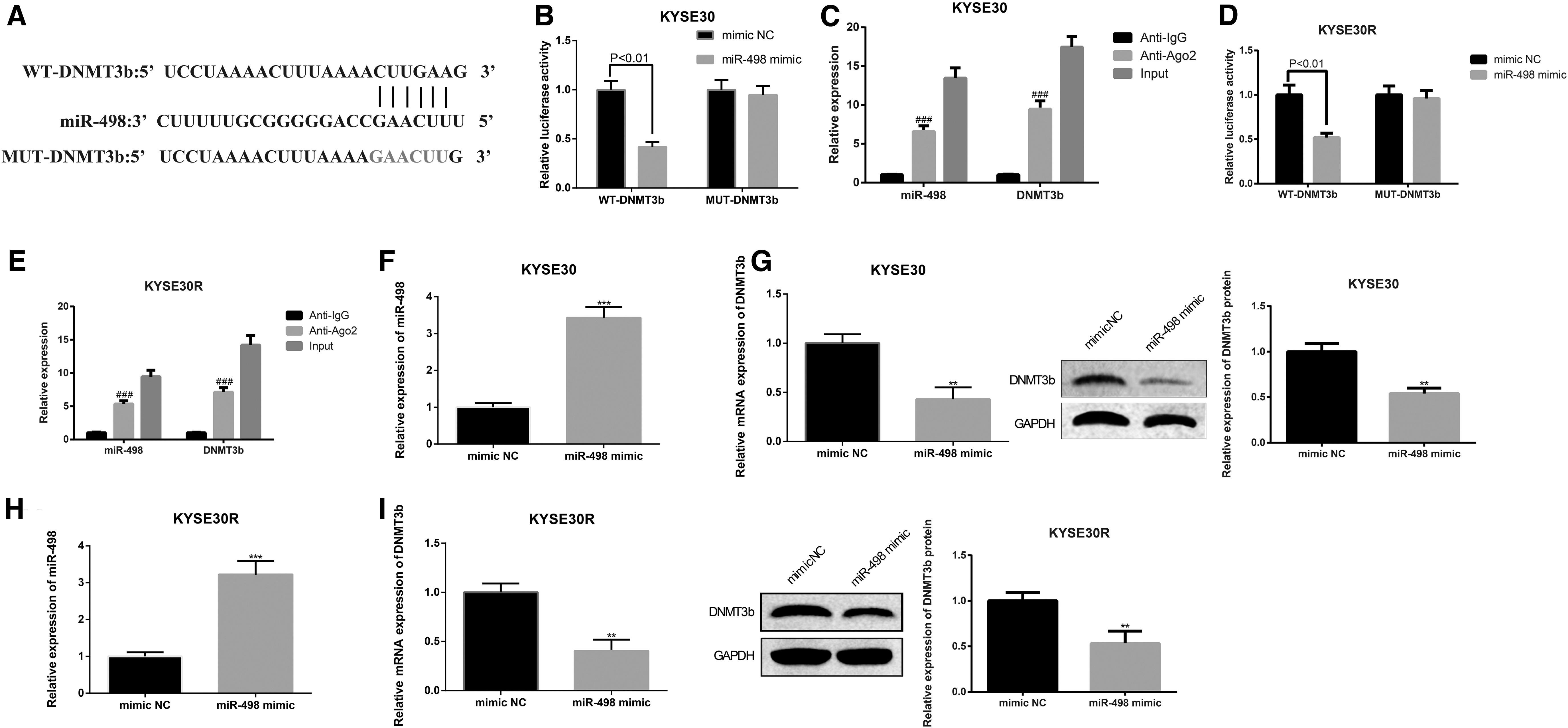
miR-498 negatively mediates DNMT3b. The StarBase predicted the binding site of DNMT3b and miR-498 (text in red indicate mutant site.)
The radiosensitization effect of miR-498 can be repressed by DNMT3b
The aforementioned results proved that DNMT3b was a target of miR-498, so the authors sought to inspect whether miR-498 confers biological effect on the regulation of DNMT3b. To this end, cell transfection was performed with cotransfection of miR-498 mimic and OE-DNMT3b in parental cell lines and radioresistant EC cell lines. qRT-PCR and Western blot on mRNA and protein levels of DNMT3b manifested that the miR-498 mimic+OE-DNMT3b group had enhanced expression of DNMT3b in comparison with the miR-498 mimic+OE-NC group (p < 0.05, Fig. 5A, B), whereas there was no obvious change in miR-498 expression (p > 0.05). Furthermore, results of CCK-8, colony formation assay, and Annexin V/PI double staining proposed that overexpression of miR-498 and DNMT3b increased cell viability (p < 0.05, Fig. 5C) and colony formation rate (p < 0.05, Fig. 5D), and decreased cell apoptotic rate (p < 0.05, Fig. 5E, vs. miR-498 mimic+OE-NC group). Taken together, DNMT3b upregulation can reverse the radiosensitization effect of miR-498 on EC cells.
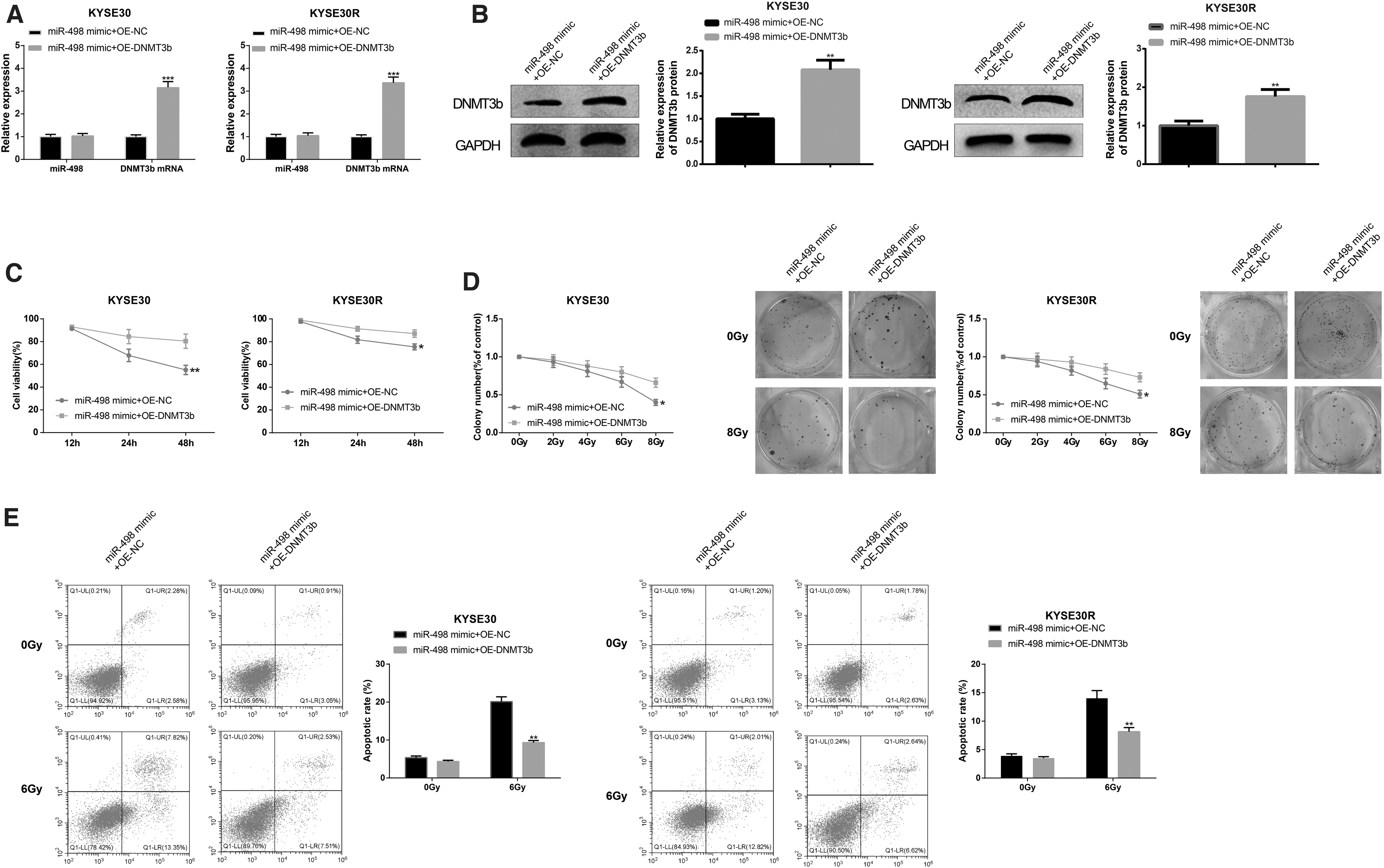
DNMT3b reduces the radiosensitization effect of miR-498 on EC cells. After cell transfection, the expression levels of miR-498 and DNMT3b were detected by qRT-PCR
miR-498 inactivates the PI3K/AKT pathway by inhibiting DNMT3b
Western blot was performed to determine the expression levels of PI3K/AKT pathway-related proteins. Transfection of miR-498 mimic (vs. mimic NC group) or si-DNMT3b (vs. si-NC group) was found to suppress protein levels of p-PI3K and p-AKT (p < 0.05, Fig. 6A, B), whereas cotransfection of miR-498 mimic and OE-DNMT3b heightened expression levels of p-PI3K and p-AKT (p < 0.05, Fig. 6A, B, vs. miR-498 mimic+OE-NC group). The mentioned findings suggested that miR-498 enhances the sensitivity of EC cells to irradiation by DNMT3b suppression, which is related to the inactivation of PI3K/AKT signaling pathway.

MiR-498 enhances radiosensitivity of EC cells by DNMT3b repression through blocking PI3K/AKT pathway. The protein levels of p-PI3K and p-AKT in KYSE30
Discussion
EC is regarded as one of malignant tumors with the highest lethal rate in the world. 7 The optimal treatment for patients with advanced EC is a combination of radiotherapy and surgery, whereas patients with EC benefit little from radiotherapy due to radioresistance. 17 Hence, searching for effective radiation sensitizers and novel radiation resistance indicators is an urgent issue to improve radiation therapy. In this study, the expression patterns of miR-498 between radioresistant EC cell line KYSE30R and its parental cell line KYSE30 were characterized to investigate the mechanisms of acquired radioresistance of EC. It was revealed that the sensitivity of EC cells toward radiotherapy is mediated by miR-498 through targeting DNMT3b and inactivating the PI3K/AKT pathway.
Recent research has yielded repressed expression of miR-498 in EC cells and tissues. 18 –20 In this study, low expression of miR-498 was found in EC cell lines as well as tissues obtained from EC patients. These results prompted us to probe whether miR-498 elicits a regulatory effect on the progression of EC. An increasing body of evidence has proven that miRNAs can serve as tumor suppressors or promoters to control various basic cellular functions in EC. 21 –23 Here, results of CCK-8, colony formation assay, and Annexin V/PI double staining exhibited that miR-498 may act as a tumor suppressor in EC by modulating cellular functions as evidenced by restrained EC cell proliferation ability and facilitated cell apoptosis level after miR-498 upregulation. These data are in agreement with the earlier findings showing that miR-498 is implicated in hepatocellular carcinoma regarding cell proliferation, invasion, and migration. 24 In acute myeloid leukemia, miR-498 can inhibit malignant cell proliferation and induces cell apoptosis. 25 In addition, it was further discovered that tissues with radioresistance had a lower miR-498 level than tissues with radiosensitivity. A more detailed characterization of miR-498 through gain-of-function research revealed that miR-498 can serve as an EC suppressor, which not only participated in proliferation and apoptosis of EC cells sensitive toward irradiation but also implicated in EC cells resistant to irradiation, eventually impacting in EC response to radiotherapy. Altogether, the findings highlight that miR-498 can enhance the sensitivity of EC cells to radiation. Despite significant merits of miR-498 in the progression of EC have been expounded, the mechanisms responsible for the early cellular events and pathogenesis of EC remain unclear to date.
The overexpressed DNMT has been inspected in a variety of malignancies, including lung and colorectal tumors. 26,27 Hence, the authors focused on the profile and potential function of DNMT3b in EC. Initially, highly expressed DNMT3b was found in EC tissues and cells in vivo and in vitro, and the radioresistance group possessed a higher level of DNMT3b than the radiosensitivity group. Based on these observations, it was speculated that DNMT3b may diminish the sensitivity of EC cells to radiotherapy. As expected, transfection with si-DNMT3b inhibited EC cell proliferation and induced cell apoptosis. Consistent evidence was found in a former study that DNMT inhibitors radiosensitize human cancer cells by suppressing DNA repair activity. 28 Furthermore, the dual-luciferase reporter gene assay verified that DNMT3b is a target gene of miR-498. Subsequently, the follow-up experiments were undertaken to seek further evidence for the potential synergistic effect of DNMT3b on the regulation of EC radiotherapy resistance. DNMT3b can reverse the radiosensitization effect of miR-498 on EC cells as evidenced by elevated cell viability and colony formation rate, and repressed cell apoptotic ability after overexpression of miR-498 and DNMT3b. Taking these data together, miR-498 mitigates EC by targeting DNMT3b to enhance the sensitivity of EC cells to radiotherapy.
In the PI3K/AKT pathway, PI3K activation can catalyze the phosphorylation of 3,4,5-phosphatidylinositol triphosphate, which, in turn, activates protein kinase AKT to promote cell growth. 29 Moreover, the PI3K/AKT signaling pathway is essential for cancer cell survival, apoptosis, proliferation, and invasion, and crucial mediators in this pathway can be altered by DNMT3b. 30 In this study, the PI3K/AKT pathway was identified and found to be involved in the proliferation and apoptosis of EC cells, which was negatively mediated by miR-498/DNMT3b. These findings suggest that miR-498 represses DNMT3b expression and inactivates the PI3K/AKT signaling pathway to enhance the sensitivity of EC cells toward irradiation, thereby ameliorating EC.
In summary, this study highlights that upregulation of miR-498 inactivates the PI3K/AKT signaling by targeting DNMT3b, leading to increased radiosensitivity of EC cells. In addition, targeting this novel miR-498/DNMT3b axis in EC patients with radioresistance holds great promise in the prevention therapy resistance and the formation of distant metastases of EC. Owing to the limited sample size and experiment period, an extended investigation into the inhibition effects of miR-498 on human EC is required in the future to further improve the survival rate of EC patients.
Footnotes
Disclosure Statement
The authors declare there is no conflict of interests.
Funding Information
Y.J. was supported by grants from the Scientific Research Fund of Traditional Chinese Medicine of Zhejiang Province (Grant No. 2019ZB075) and the Foundation of Wenzhou Science and Technology Bureau (Grant No. Y20190603).