
Abstract
Background:
Osteosarcoma (OS) is a common, aggressive primary sarcoma of bone. Drug resistance is a huge obstacle to chemotherapy for cancer. This study aimed to investigate the role and mechanism of circ_0002060 in OS resistance to doxorubicin (DOX).
Methods:
The levels of circ_0002060, miR-198, and ATP-binding cassette subfamily B member 1 (ABCB1) in OS tissues and DOX-resistant OS cells were measured by quantitative real-time polymerase chain reaction or Western blot assay. Kaplan–Meier analysis was performed to determine the relationship between circ_0002060 expression in OS tissues and overall survival of OS patients. The half-inhibitory concentration (IC50) of DOX was calculated using the Cell Counting Kit-8 (CCK-8) assay. Proliferation and apoptosis of DOX-resistant OS cells were assessed by colony formation assay and flow cytometry. The levels of apoptosis-related proteins in DOX-resistant OS cells were measured by Western blot assay. Xenograft assay was utilized to analyze the effect of circ_0002060 on DOX resistance in vivo. The interactions among circ_0002060, miR-198, and ABCB1 in DOX-resistant OS cells were confirmed by dual-luciferase reporter assay, RNA immunoprecipitation assay, or RNA pull-down assay.
Results:
circ_0002060 and ABCB1 were upregulated, while miR-198 was downregulated in OS tissues and DOX-resistant OS cells. circ_0002060 silencing reduced DOX resistance in vitro and in vivo. Moreover, circ_0002060 enhanced DOX resistance by sponging miR-198. Besides, miR-198 decreased DOX resistance by binding to ABCB1. In addition, circ_0002060 sponged miR-198 to upregulate ABCB1 expression.
Conclusions:
circ_0002060 promoted DOX resistance and OS progression by regulating the miR-198/ABCB1 axis, suggesting that circ_0002060 might be a promising biomarker for OS therapy.
Introduction
Osteosarcoma (OS) is a common malignant bone cancer that occurs in children and adolescents. 1 With the development of medical technology, surgical resection accompanied by the introduction of chemotherapy has improved the prognosis of OS patients, and the 5-year survival rate of OS patients has reached 70%. 2 However, chemoresistance is a major obstacle to improving OS prognosis, so overcoming drug resistance is critical to improving OS treatment effectiveness.
Circular RNAs (circRNAs), a new type of noncoding RNAs, can regulate tumorigenesis and development by participating in competitive endogenous RNA mechanisms (ceRNAs). 3 Increasing studies have revealed that several circRNAs that are abnormally expressed in OS are closely related to OS pathogenesis. 4,5 For instance, circ-0000285 was highly expressed in OS and facilitated OS development by targeting microRNA-599 and increasing TGFB2 expression. 6 circMMP9 sponged microRNA-1265 to upregulate CHI3L1, thus accelerating OS progression. 7 In osteoporosis, circ_0002060, a transcript of dynein cytoplasmic 1 heavy chain 1 (DYNC1H1), was significantly elevated and might be a potential therapeutic target. 8 Therefore, circ_0002060 expression was analyzed in OS, and the authors found that circ_0002060 was markedly upregulated, hinting its potential role in OS development.
Moreover, many microRNAs (miRNAs) have been identified as playing a vital role in OS progression. 9 For example, miR-150 exerted an anticancer effect on OS progression and enhanced the sensitivity of OS cells to doxorubicin (DOX) by regulating RUNX2. 10 Dong et al. presented that miR-193b could sensitize OS cells to epirubicin by modulating FEN1-induced autophagy. 11 Furthermore, previous research discovered that miR-198 was an underlying molecular target for OS treatment. 12 Bioinformatic analysis showed that miR-198 and circ_0002060 have complementary binding sites, so it is speculated that circ_0002060 may interact with miR-198 in OS.
ATP-binding cassette subfamily B member 1 (ABCB1) is strongly related to multidrug resistance (MDR). 13 In breast cancer, miR-381 targeted ABCB1 to reduce cisplatin resistance. 14 In OS, TIPE2 strengthened cisplatin sensitivity by decreasing ABCB1 expression. 15 However, the mechanisms of miR-198 and ABCB1 in DOX resistance of OS are still barely reported.
Hence, the authors further investigated the function and potential mechanisms of circ_0002060, miR-198, and ABCB1 in DOX resistance and tumor progression in OS.
Materials and Methods
Clinical samples
Forty samples of OS tissues and adjacent normal tissues were obtained from OS patients who underwent surgery at Weifang People's Hospital between January 2010 and December 2017. The inclusion criteria were as follows: preoperatively confirmed as OS by puncture or biopsy; no immunotherapy, chemotherapy, or radiotherapy before surgery; and complete clinical data. The exclusion criteria were as follows: suffering from other malignant tumors; suffering from other types of serious diseases; patients received treatment for other clinical diseases within 3 months before admission; and pregnant or breastfeeding. This research was conducted at Weifang People's Hospital and was approved by the Ethics Committee of Weifang People's Hospital. All participants signed a written informed consent. All tissues were immediately frozen in liquid nitrogen and then stored at −80°C. After treatment with DOX, OS patients were divided into a good response group (n = 20) and poor response group (n = 20) according to the response assessment criteria.
Cell culture
Two OS cell lines (U2OS and HOS) were purchased from American Type Culture Collection (ATCC, Manassas, VA). U2OS is a moderately differentiated OS cell line obtained from a 15-year-old girl with insulin-like growth factor expression. HOS is an OS cell line from a 13-year-old girl. DOX-resistant cell lines (U2OS/DOX and HOS/DOX) were produced by U2OS and HOS cells exposed to gradient doses of DOX (Solarbio, Beijing, China). All cells were incubated in Dulbecco's modified Eagle's medium (DMEM; Gibco, Carlsbad, CA) supplemented with 10% fetal bovine serum (FBS; Gibco) with 5% CO2 at 37°C.
Cell transfection
Small interfering RNA (siRNA) against circ_0002060 (si-circ_0002060#1, si-circ_0002060#2, and si-circ_0002060#3), siRNA control (si-NC), miR-198 mimics (miR-198), mimic control (miR-NC), circ_0002060 overexpression vector (circ_0002060), empty vector (pCD-ciR), miR-198 inhibitor (anti-miR-198), inhibitor control (anti-miR-NC), ABCB1 overexpression vector (ABCB1), and empty vector (pcDNA) were commercially obtained from GenePharma (Shanghai, China). The vectors and oligonucleotides were transfected into U2OS/DOX and HOS/DOX cells using Lipofectamine 3000 (Invitrogen, Carlsbad, CA).
Quantitative real-time polymerase chain reaction
After extracting RNA with TRIzol (Invitrogen), RNA was reverse transcribed using the HiScript II One Step RT-PCR Kit (Vazyme, Nanjing, China) or miRNA 1st Strand cDNA Synthesis Kit (Vazyme). Then, the quantitative real-time polymerase chain reaction (qRT-PCR) was carried out using the AceQ qPCR SYBR Green Master Mix (Vazyme). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and U6 were used as internal controls. Primers used are shown below: miR-198-F: 5′-GGTCCAGAGGGGAGAT-3′, miR-198-R: 5′-GAATACCTCGGACCCTGC-3′; ABCB1-F: 5′-CCCATCATTGCAATAGCAGG-3′, ABCB1-R: 5′-GTTCAAACTTCTGCTCCTGA-3′; GAPDH-F: 5′-TGCACCACCAACTGCTTAGC-3′, GAPDH-R: 5′-GGCATGGACTGTGGTCATGAG-3′; U6-F: 5′-CTCGCTTCGGCAGCACA-3′, U6-R: 5′-AACGCTTCACGAATTTGCGT-3′; and 18sRNA-F: 5′-AAACGGCTACCACATCCA-3′, 18sRNA-R: 5′-CACCAGACTTGCCCCTCCA-3′. The primers of circ_0002060 were purchased from GenePharma.
Cell counting kit-8 assay
Cells (3 × 103) were plated into 96-well plates and incubated with escalating doses of DOX for the indicated time. Subsequently, cells were reacted with 10 μL of Cell Counting Kit-8 (CCK-8) solution (Solarbio) for 2 h. Next, a Multi-Mode Reader (BioTek, Burlington, VT) was used to measure the optical density at 450 nm to assess cell viability. The half-maximal inhibitory concentration (IC50) of DOX is the concentration at which cell viability is reduced to 50%.
Colony formation assay
The treated cells were seeded into six-well plates and cultured in DMEM containing 10% FBS at 37°C for 14 d. After staining with 0.5% crystal violet, colonies were imaged and counted at least three times.
Flow cytometry
The treated cells were inoculated into six-well plates and washed with cold PBS. Then, annexin V-fluorescein isothiocyanate (annexin V-FITC)/propidium iodide (PI) apoptosis detection kit (Invitrogen) was used to detect cell apoptosis. Finally, the Attune NxT Flow Cytometer (Thermo Fisher Scientific, Waltham, MA) was utilized to monitor the apoptosis rate.
Western blot assay
After lysing cells with RIPA buffer (Solarbio), the extracted proteins were separated by polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA). Then, the membranes were probed with primary antibodies against caspase 3 (ab13847; Abcam, Cambridge, United Kingdom), B cell lymphoma 2 (Bcl-2) (ab196495; Abcam), Bcl-2-associated X protein (Bax) (ab53154; Abcam), ABCB1 (ab129450; Abcam), and GAPDH (ab9485; Abcam). Next, the membranes were treated with a secondary antibody (ab7090; Abcam). The signal intensity was measured using the enhanced chemiluminescence system (Millipore).
Xenograft tumor experiment
U2OS/DOX cells were transfected with the lentivirus carrying circ_0002060 short hairpin RNA (sh-circ_0002060) or the negative control (sh-NC). Subsequently, stably transfected cells (5 × 106) were subcutaneously injected into the right back of BALB/c nude mice (n = 6 per group). Then, DOX (15 mg/kg) was intravenously administered to nude mice twice a week. Tumor volume was monitored every week. After 5 weeks, mice were sacrificed, and xenografts were weighted. The xenograft assay was ratified by the Animal Research Committee of Weifang People's Hospital.
Dual-luciferase reporter assay
The sequences of circ_0002060 or ABCB1 3′UTR containing miR-198 binding sites or mutant were amplified and then inserted into the pmirGLO vector (Promega, Madison, WI), named circ_0002060-wt, circ_0002060-mut, ABCB1 3′UTR-wt, or ABCB1 3′UTR-mut reporter. Then, the corresponding luciferase reporter and miR-198 or miR-NC were cotransfected into U2OS/DOX and HOS/DOX. Finally, the luciferase intensity was monitored using a Dual-Luciferase Reporter Assay Kit (Promega).
RNA immunoprecipitation assay
RNA immunoprecipitation (RIP) analysis was carried out using the EZ-Magna RIP kit (Millipore). Briefly, U2OS/DOX and HOS/DOX cells were lysed by RIP lysis buffer. Then, cell lysates were incubated with magnetic beads conjugated with Ago2 antibody or IgG antibody. The enrichment of circ_0002060, miR-198, or ABCB1 was measured by qRT-PCR.
RNA pull-down assay
The biotin-labeled circ_0002060 probe (Bio-circ_0002060) and control probe (Bio-NC) were purchased from GenePharma. Briefly, biotinylated probes were reacted with M-280 streptavidin Dynabeads (Invitrogen) at 37°C for 2 h to construct probe-coated beads. Next, the cells were lysed and incubated with probe-coated beads at 4°C for 3 h. Finally, the expression of miR-198 was measured by qRT-PCR.
Statistical analysis
All data are expressed as mean ± standard deviation using GraphPad Prism 7.0 software (GraphPad, San Diego, CA). Differences were tested using Student's t-test or one-way analysis of variance. When p < 0.05, the difference was considered statistically significant.
Results
circ_0002060 was upregulated in OS tissues and DOX-resistant OS cells
The authors detected the expression of circ_0002060 in OS tissues and adjacent normal tissues, and results show that circ_0002060 expression in OS tissues was strikingly increased compared with noncancer tissues (Fig. 1A). Besides, the circ_0002060 level in the poor response group was significantly higher than that in the good response group (Supplementary Fig. S1A). Kaplan–Meier analysis was performed to explore the relationship between circ_0002060 expression and overall survival in OS patients. The results showed that high circ_0002060 levels were closely related to poor prognosis in OS patients (Fig. 1B). In addition, the expression of circ_0002060 in U2OS/DOX and HOS/DOX cells was overtly higher than that in U2OS and HOS cells (Fig. 1C). Moreover, circ_0002060 was resistant to RNase R compared with its linear isoform, DYNC1H1 (Fig. 1D, E). In addition, DOX-resistant cells were treated with actinomycin D, and qRT-PCR showed that circ_0002060 had a half-life of more than 24 h, while DYNC1H1 mRNA had a half-life of less than 6 h (Fig. 1F, G). These data revealed that circ_0002060 might be an oncogene in OS.

circ_0002060 was upregulated in OS tissues and DOX-resistant OS cells.
circ_0002060 knockdown decreased DOX resistance in vitro and in vivo
To explore the function of circ_0002060 in OS cells, circ_0002060 was silenced by transfecting si-circ_0002060#1, si-circ_0002060#2, or si-circ_0002060#3 into U2OS/DOX and HOS/DOX cells. The knockdown efficiency of circ_0002060 was detected by qRT-PCR analysis, and the expression of DYNC1H1 was not affected (Fig. 2A, B). CCK-8 assay showed that downregulation of circ_0002060 reduced the IC50 value of DOX in U2OS/DOX and HOS/DOX cells (Fig. 2C, D). Colony formation assay demonstrated that circ_0002060 silencing and DOX treatment significantly decreased the proliferation ability of U2OS/DOX and HOS/DOX cells (Fig. 2E, F). Additionally, flow cytometry revealed that circ_0002060 knockdown and DOX stimulation markedly promoted the apoptosis of DOX-resistant OS cells (Fig. 2G–I). Consistently, circ_0002060 silencing and DOX stimulation resulted in a striking reduction in Bcl-2 expression and a significant increase in c-caspase 3/caspase 3 and Bax levels (Fig. 2J, K). Moreover, a xenograft mouse model was established to investigate the effect of circ_0002060 on DOX sensitivity in vivo. As shown in Figure 2L and M, tumor volume and weight were observably reduced in the sh-circ_0002060 group compared with the sh-NC group, and DOX treatment enhanced this effect. These results revealed that silencing of circ_0002060 reduced DOX resistance in vitro and in vivo.
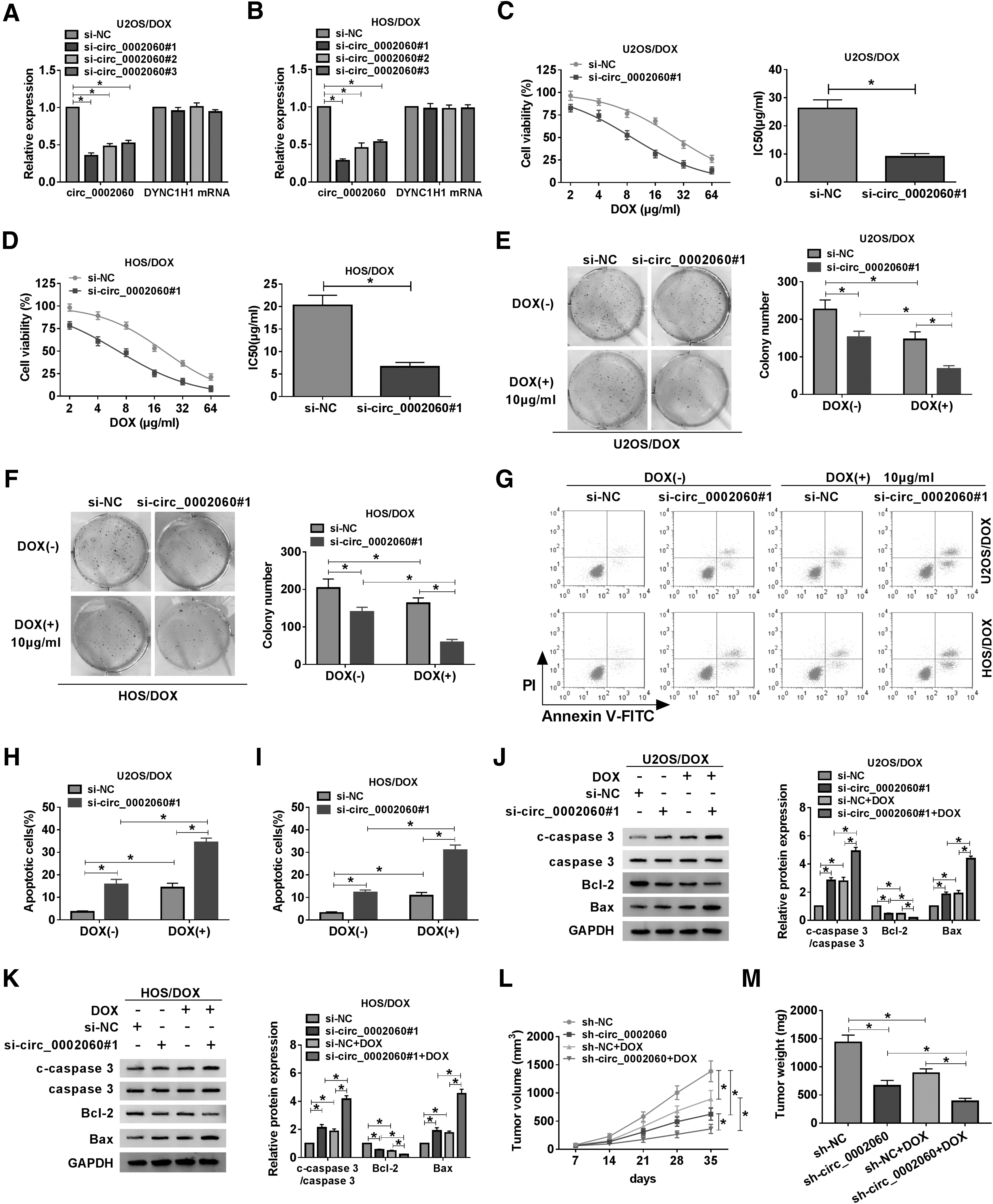
circ_0002060 knockdown decreased DOX resistance in vitro and in vivo.
circ_0002060 directly targeted miR-198
Next, the Circular RNA Interactome online database determined that circ_0002060 and miR-198 had complementary sequences (Fig. 3A). Besides, the expression of miR-198 in OS tissues was significantly lower than that in normal tissues (Fig. 3B). As shown in Supplementary Figure S1B, the miR-198 level in the poor response group was markedly reduced compared with the good response group. In OS tissues, circ_0002060 expression was inversely correlated with miR-198 expression (Fig. 3C). First, the dual-luciferase reporter assay suggested that miR-198 mimics inhibited the luciferase activity of circ_0002060-wt reporter (Fig. 3D, E). Then, the relationship between circ_0002060 and miR-198 was verified by RIP and RNA pull-down assays. As shown in Figure 3F and G, circ_0002060 and miR-198 were markedly enriched in the Ago2 group compared with the IgG group. Additionally, RNA pull-down assay revealed that miR-198 could be pulled down by Bio-circ_0002060, but not by Bio-NC (Fig. 3H). Compared with U2OS and HOS cells, the miR-198 level in U2OS/DOX and HOS/DOX cells was significantly reduced (Fig. 3I). In addition, circ_0002060 silencing strikingly elevated miR-198 expression, and circ_0002060 overexpression markedly decreased miR-198 expression (Fig. 3J, K). These data indicated that circ_0002060 was a sponge of miR-198.
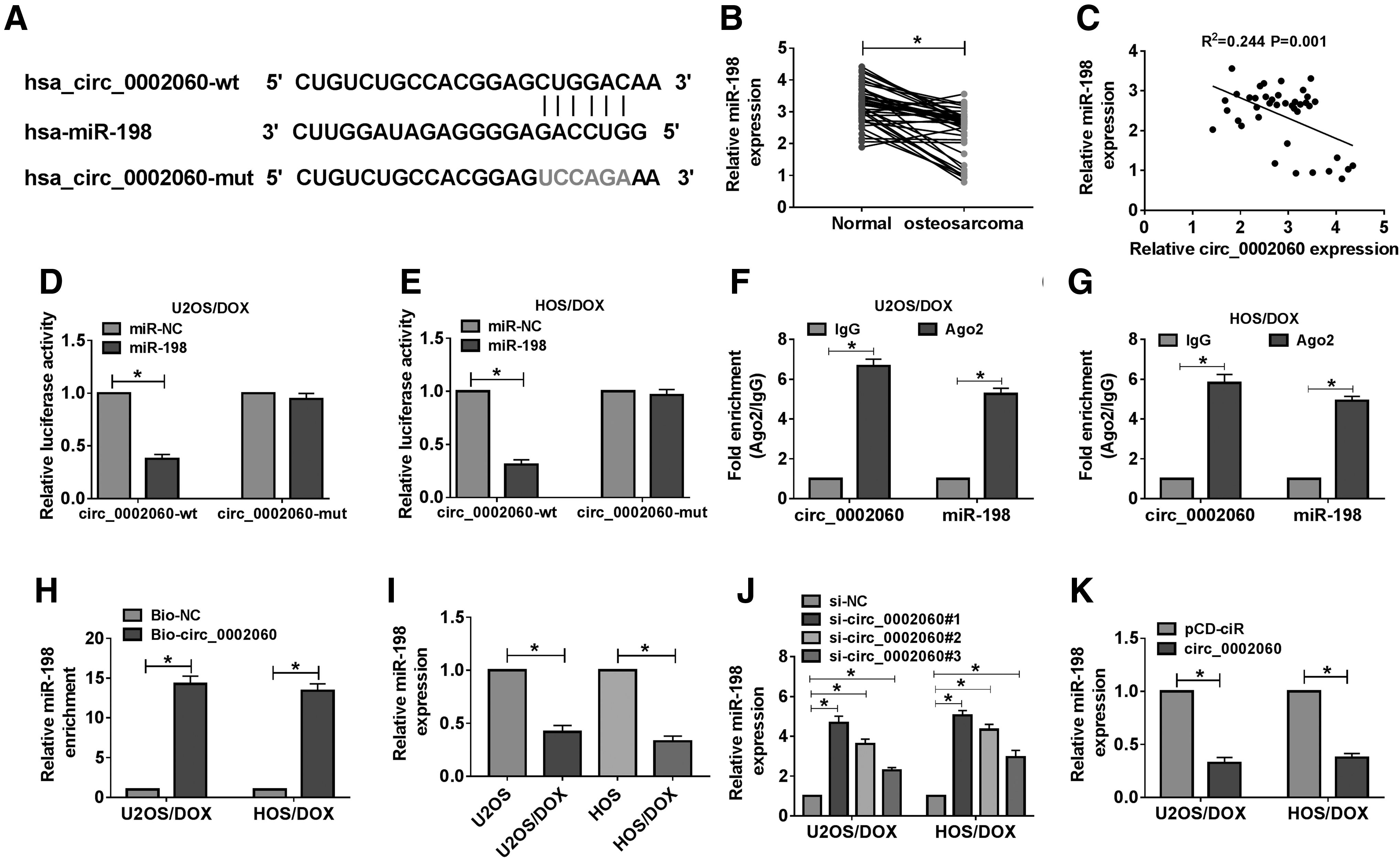
circ_0002060 directly targeted miR-198.
circ_0002060 regulated DOX resistance by sponging miR-198
To investigate the roles of circ_0002060 and miR-198 in DOX resistance, U2OS/DOX and HOS/DOX cells were transfected with si-NC, si-circ_0002060#1, si-circ_0002060#1+anti-miR-NC, or si-circ_0002060#1+anti-miR-198. The results showed that the addition of anti-miR-198 reversed the increase of miR-198 expression caused by circ_0002060 silencing (Fig. 4A). CCK-8 analysis revealed that inhibition of miR-198 eliminated the reduction in DOX resistance induced by circ_0002060 knockdown (Fig. 4B–E). Colony formation assay showed that downregulation of miR-198 partially attenuated the reduction in colony number caused by DOX treatment or circ_0002060 silencing (Fig. 4F, G). In addition, transfection with miR-198 partially reversed the effect of DOX stimulation or circ_0002060 depletion on cell apoptosis (Fig. 4H, I). Consistently, DOX treatment and circ_0002060 silencing increased the levels of c-caspase 3/caspase 3 and Bax and decreased the level of Bcl-2, while miR-198 knockdown partially abolished the effects (Fig. 4J–M). These data indicated that circ_0002060 modulated DOX resistance by sponging miR-198 in DOX-resistant OS cells.
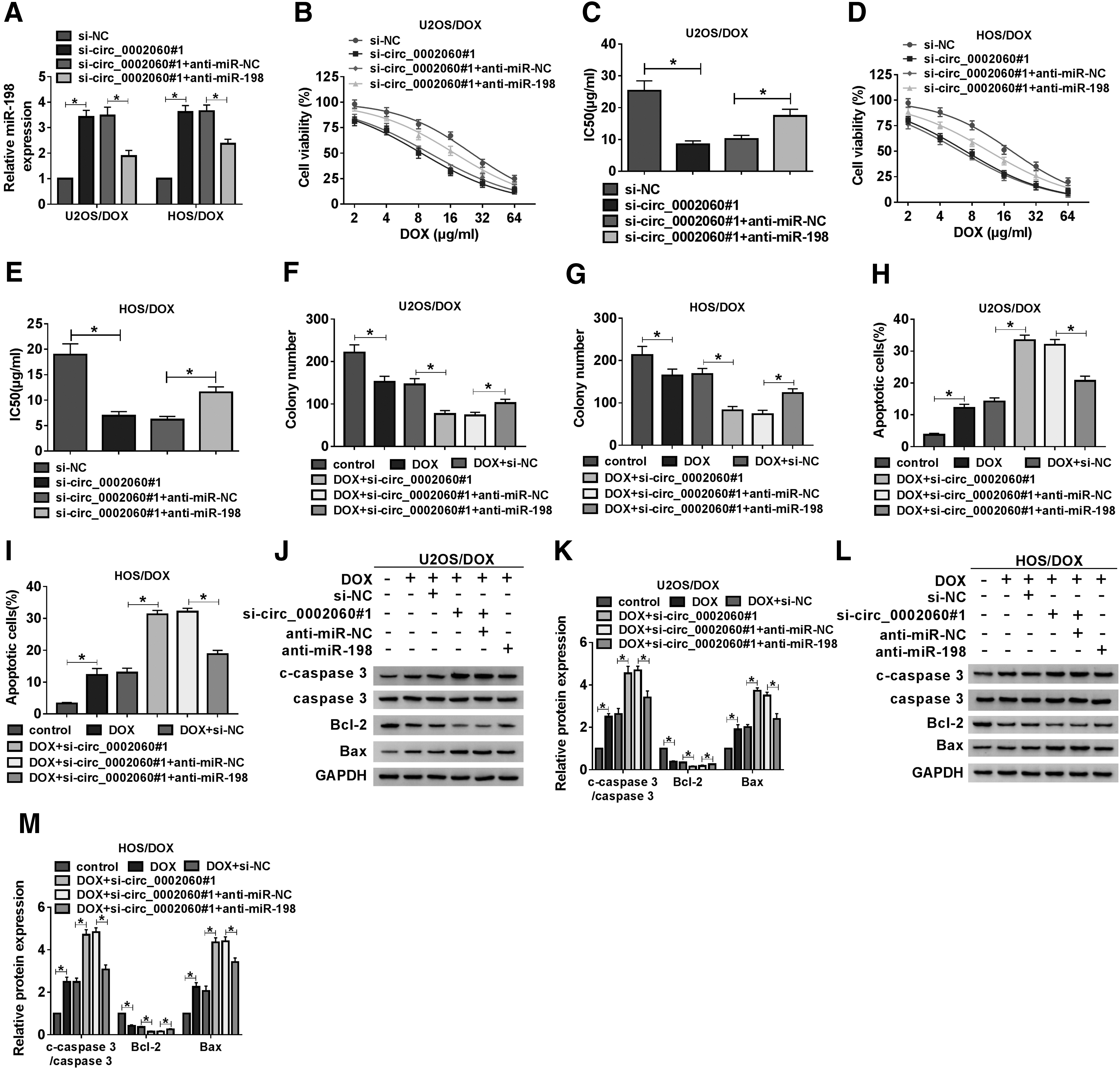
circ_0002060 regulated DOX resistance by sponging miR-198. U2OS/DOX and HOS/DOX cells were transfected with si-NC, si-circ_0002060#1, si-circ_0002060#1+anti-miR-NC, or si-circ_0002060#1+anti-miR-198, respectively.
ABCB1 was a target of miR-198
Furthermore, the TargetScan online database predicted that miR-198 and ABCB1 3′UTR had putative binding sites (Fig. 5A). First, qRT-PCR and Western blot assays suggested that mRNA and protein levels of ABCB1 were remarkably increased in OS tissues relative to normal tissues (Fig. 5B, C). Additionally, the ABCB1 mRNA level was strikingly elevated in the poor response group relative to the good response group (Supplementary Fig. S1C). In addition, miR-198 and ABCB1 levels were negatively correlated in OS tissues (Fig. 5D). Then, the dual-luciferase reporter assay demonstrated that mature miR-198 reduced the luciferase activity of ABCB1 3′UTR-wt reporter (Fig. 5E, F). RIP assay was performed to validate the interaction between miR-198 and ABCB1, and results showed that miR-198 and ABCB1 were significantly enriched in the Ago2 group compared with the IgG group (Fig. 5G, H). Compared with U2OS and HOS cells, mRNA and protein levels of ABCB1 in U2OS/DOX and HOS/DOX cells were significantly upregulated (Fig. 5I, J). Besides, miR-198 mimics suppressed ABCB1 mRNA and protein expression, while miR-198 knockdown has the opposite effect (Fig. 5K–N). These data revealed that ABCB1 was a target of miR-198.
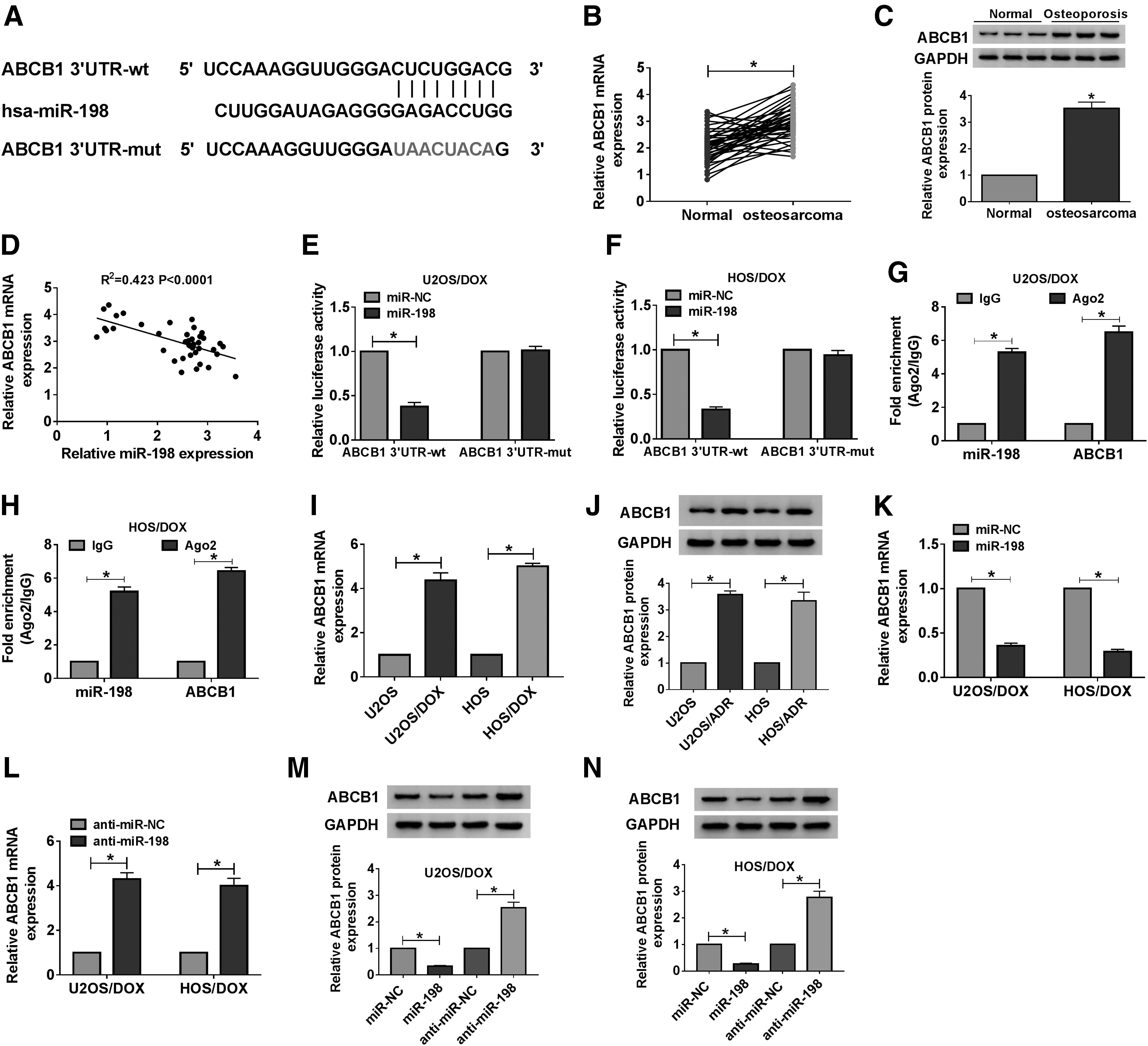
ABCB1 was a target of miR-198.
miR-198 reduced DOX resistance by targeting ABCB1
To explore the relationship between miR-198 and ABCB1 in DOX resistance, U2OS/DOX and HOS/DOX cells were transfected with miR-NC, miR-198, miR-198+pcDNA, or miR-198+ABCB1. First, transfection with ABCB1 reversed the decrease in ABCB1 expression caused by miR-198 overexpression (Fig. 6A–C). Second, upregulation of miR-198 decreased the IC50 value of DOX, whereas the impact was abolished after transfection with ABCB1 (Fig. 6D–G). Colony formation assay revealed that overexpression of ABCB1 partially abrogated the decrease in colony number induced by DOX stimulation or miR-198 upregulation (Fig. 6H, I). Additionally, introduction of ABCB1 partially rescued the promoting effect of DOX treatment or miR-198 overexpression on cell apoptosis (Fig. 6J, K). Similarly, DOX stimulation and miR-198 upregulation resulted in increased c-caspase 3/caspase 3 and Bax expression and decreased Bcl-2 expression, while these changes were partially reversed by overexpressing ABCB1 (Fig. 6L–O). These data showed that miR-198 decreased DOX resistance by targeting ABCB1 in U2OS/DOX and HOS/DOX cells.

miR-198 reduced DOX resistance by targeting ABCB1. U2OS/DOX and HOS/DOX cells were transduced with miR-NC, miR-198, miR-198+pcDNA, or miR-198+ABCB1, respectively.
circ_0002060 modulated ABCB1 expression by regulating miR-198
First, Spearman's correlation analysis illustrated that circ_0002060 and ABCB1 levels were positively correlated in OS tissues (Fig. 7A). Then, to elucidate the regulatory mechanisms of circ_0002060, miR-198, and ABCB1, U2OS/DOX and HOS/DOX cells were transduced with si-NC, si-circ_0002060#1, si-circ_0002060#1+anti-miR-NC, or si-circ_0002060#1+anti-miR-198. As displayed in Figure 7B–D, inhibition of miR-198 reversed the decrease of ABCB1 expression caused by circ_0002060 knockdown. These results concluded that circ_0002060 silencing reduced ABCB1 expression by sponging miR-198.
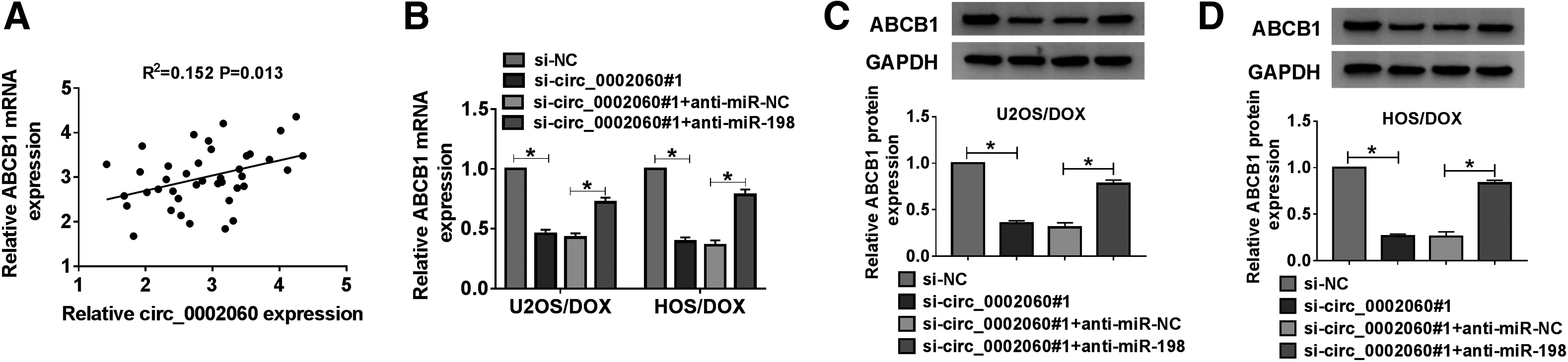
circ_0002060 modulated ABCB1 expression by regulating miR-198.
Discussion
Recently, the occurrence of chemoresistance has become a major stumbling block in OS treatment. 16 Besides, adriamycin has significant medicinal properties in adjuvant chemotherapy of localized high-grade OS. 17 Additionally, large amounts of abnormally expressed noncoding RNAs (ncRNAs) play a crucial role in antitumor drug resistance. 18 Therefore, the authors strengthened the study of circRNAs in DOX resistance of OS.
Accumulating evidence demonstrated that circRNAs could mediate the expression of target mRNAs by serving as ceRNAs or miRNA sponges, thereby regulating chemotherapy resistance. 19 For example, circKDM4C potentiated DOX sensitivity and hindered tumorigenesis in breast cancer by sponging miR-548p and upregulating PBLD. 20 Additionally, Hu et al. presented that circ-LARP4 attenuated cisplatin and DOX resistance in OS by binding to miR-424. 21 The authors' research demonstrated that circ_0002060 was remarkably upregulated in OS tissues and DOX-resistant OS cells, and circ_0002060 silencing reduced DOX resistance in OS, indicating that circ_0002060 was an oncogene in OS. Furthermore, the authors revealed the negative regulation and binding relationship between circ_0002060 and miR-198.
Several research reports corroborate miR-198 as an inhibitor of various tumors, including gastric cancer, 22 colorectal cancer, 23 and breast cancer. 24 Furthermore, mining of miRNAs provides a new pathway for early diagnosis of OS. Zhang et al. revealed that miR-198 functioned as a potential target for OS treatment by regulating ROCK1. 12 In addition, several studies demonstrated that miRNAs could regulate chemotherapy resistance of OS by mediating multiple potential molecular mechanisms such as DNA damage response, autophagy induction, and signal transduction. 25 Chen et al. found that miR-198 weakened temozolomide resistance by modulating MGMT in glioblastoma cells. 26 Huang et al. discovered that miR-198 reduced the increase of cisplatin resistance caused by circAKT3 by targeting PIK3R1 in gastric cancer. 27 Nevertheless, the effect of miR-198 on DOX resistance of OS remains unclear. In this study, the authors showed that miR-198 expression was remarkably decreased and circ_0002060 sponged miR-198 to enhance DOX resistance in OS.
Meanwhile, the existence of MDR has made tumor treatment more difficult. 28 ABCB1 (also known as MDR1) is considered to be the main factor inducing MDR. 29 In hepatocellular carcinoma, miR-122 improved oxaliplatin sensitivity by repressing ABCB1 expression to inactivate the Wnt/β-catenin pathway. 30 In colorectal cancer, overexpression of ABCB1 attenuated oxaliplatin sensitivity, which might be regulated by miR-302c-5p. 31 In addition, many studies discovered that ABCB1 was remarkably elevated in OS, 32 and ABCB1 inhibitor undermined OS resistance to DOX and paclitaxel. 33 The authors demonstrated that ABCB1 was a target of miR-198 in DOX-resistant OS cells and miR-198 targeted ABCB1 to regulate DOX resistance in OS.
In conclusion, circ_0002060 enhanced DOX resistance and facilitated OS progression by modulating the miR-198/ABCB1 axis. These findings provided a promising, new therapeutic target for OS patients.
Footnotes
Disclosure Statement
The authors declare that they have no conflicts of interest.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.