
Abstract
Background:
Gallbladder cancer (GBC) is the most common biliary tract malignancy. Long noncoding RNA urothelial carcinoma-associated 1 (UCA1) and MicroRNA-613 (miR-613) have been reported to be involved in the progression of various cancers. However, the regulatory mechanism between UCA1 and miR-613 in GBC is unclear.
Materials and Methods:
The expression levels of UCA1, miR-613, and secreted protein/osteonectin, cwcv, and kazal-like domains proteoglycan 1 (SPOCK1) mRNA were detected using quantitative real-time polymerase chain reaction. Cell proliferation, migration, invasion, and apoptosis were determined with MTT, transwell, or flow cytometry assays. The levels of SPOCK1 protein, Bax, cleaved-casp-3, and Bcl-2 were determined by Western blot analysis. The relationship between miR-613 and UCA1 or SPOCK1 was verified through dual-luciferase reporter and/or RNA immunoprecipitation assays. Xenograft assay was performed to verify the role of UCA1 in vivo.
Results:
UCA1 and SPOCK1 were upregulated, whereas miR-613 was downregulated in GBC tissues and cells. UCA1 silencing decreased tumor growth in vivo and impeded proliferation, migration, invasion, and induced apoptosis of GBC cells in vitro. Notably, UCA1 acted as a sponge for miR-613, which targeted SPOCK1 in GBC cells. Moreover, UCA1 enhancement reversed the repressive impact of miR-613 mimic on the malignancy of GBC cells. UCA1 regulated SPOCK1 expression through adsorbing miR-613. Furthermore, SPOCK1 elevation overturned UCA1 silencing mediated the malignant behaviors of GBC cells.
Conclusion:
UCA1 knockdown suppressed GBC progression through downregulating SPOCK1 via sponging miR-613, providing an evidence for UCA1 as a target for GBC treatment.
Introduction
Gallbladder cancer (GBC), which originates from biliary tract cells, is a malignant tumor. 1 The 5-year survival rate of advanced GBC is only about 5%, which is mainly due to the low specificity of tumor diagnosis, and thus miss the optimal treatment time. 2,3 Therefore, it is urgent to further study the molecular mechanism of GBC development to provide effective targets for the diagnosis and treatment of GBC.
Long-noncoding RNAs (lncRNAs), a class of transcripts without protein-coding potential, are important regulatory factors in a variety of biological processes. 4 Numerous researches have shown that the dysregulation of lncRNAs may be related to the advancement of cancers. 5 lncRNA urothelial carcinoma-associated 1 (UCA1), an oncofetal gene located on chromosome 19p13.12 (2314 bp), was primitively found in human bladder cancer cells. UCA1 was reported to be involved in the development of various cancers. 6 –8 For instance, Nie et al. indicated that UCA1 regulated ERBB4 expression through sponging miR-193a-3p, which contributed to the progression of nonsmall-cell lung cancer. 6 Report of Cai et al. discovered that high UCA1 expression was associated with the tumor size, lymph node metastasis, and tumor–node–metastasis (TNM) stage of GBC patients. 9 Also, GBC patients with higher UCA1 expression had a shorter survival rate. 9 Additionally, UCA1 overexpression accelerated GBC cell growth in vitro and mouse models. 9 However, the molecular mechanisms by which UCA1 modulates GBC progression are unclear.
MicroRNAs (miRNAs) are a type of short noncoding RNAs that regulate gene expression through binding to the 3′ untranslated region (UTR) of target mRNAs. 10 Studies have shown that miRNAs play a crucial role in human physiological and pathological processes. 11,12 A large amount of miRNAs have been proved as potential biomarkers and targets for various cancer diagnosis and treatment. 13,14 MicroRNA-613 (miR-613) was demonstrated to play important roles in tumor progression. For instance, miR-613 suppressed tumor progression in triple-negative breast cancer, 15 bladder cancer, 16 and laryngeal squamous cell cancer. 17 To date, the role of miR-613 in GBC is indistinct.
Cysteine-rich acidic secreted protein/osteonectin, cwcv, and kazal-like domains proteoglycan 1 (SPOCK1), a member of ca2+-binding proteoglycan family, is involved in cell proliferation, adhesion, and migration. 18 A series of studies have shown that SPOCK1 acts as an oncogene in multiple cancers. 19 –21 In addition, Shu et al. reported that SPOCK1 was associated with the development of GBC. 22 Up to now, it is unclear whether SPOCK1 can be regulated by UCA1 and miR-613 in GBC.
Hence, the study aimed to explore the role and regulatory mechanism of UCA1 in GBC, and the results manifested that UCA1 knockdown impeded GBC progression through the miR-613/SPOCK1 axis.
Materials and Methods
GBC samples
Thirty paired GBC tissue samples and adjacent normal tissue samples were collected from GBC patients who accepted a radical resection (2 cm margin around tumor tissue) at the Loudi Central Hospital of Hunan. The histology and pathology of fresh samples were verified by at last two pathologists. The TNM staging of GBC was classified according to the International Anti-Cancer Association (UICC). GBC patients did not receive chemotherapy, radiotherapy, or other anticancer drugs before surgery. This study was approved by the Ethics Committee of Loudi Central Hospital of Hunan. All patients had signed the informed consents before undergoing surgical resection.
Cell culture and transfection
Human GBC cell lines (SGC-996, GBC-SD, and NOZ) and human intrahepatic bile duct epithelial cells (HIBEpiC) were used in the study. SGC-996, GBC-SD, and HIBEpiC cells were obtained from Jingkang Biological Engineering Co., Ltd. (Shanghai, China). NOZ cells were obtained from Bena culture collection (Suzhou, China). These cells were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS) (Thermo Fisher Scientific) and 100 U/mL penicillin/streptomycin (Corning, Inc., Corning, NY, USA) in an atmosphere with 5% CO2 at 37°C.
Short hairpin RNA (sh-RNA) targeting UCA1 (sh-UCA1:5′-CCGGGGGCTTGGGACATTTCACTTTCTCGAGAAAGTGAAATGTCCCAAGCCCTTTTTG-3′) and negative control (sh-NC: 5′-UUCUCCGAACGUGUCACGUTT-3′) were synthesized by GenePharma (Shanghai, China). The sequences of UCA1 and SPOCK1 were cloned into the pcDNA3.1 vector (vector) (Invitrogen, Carlsbad, CA, USA) to construct the overexpression vectors for UCA1 and SPOCK1, respectively. MiR-613 mimic and inhibitor (miR-613 and anti-miR-613) and their corresponding negative controls (miR-NC and anti-miR-NC) were purchased from GenePharma. Oligonucleotides or plasmids were transfected into NOZ and GBC-SD cells using the Lipofectamine 2000 reagent (Thermo Fisher Scientific).
Quantitative real-time polymerase chain reaction
Total RNA was extracted from GBC tissues and cells by TRIzol reagent (TransGen Biotech, Beijing, China). Reverse transcription was executed using the All-in-One™ miRNA First-Strand cDNA Synthesis Kit (GeneCopoeia, Rockville, MD, USA) or the Prime Script™ RT Reagent Kit (TaKaRa, Shiga, Japan). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed using the Platinum SYBR Green qPCR SuperMix UDG (Invitrogen). The primer sequences were listed as below: UCA1 (forward, 5′-CTCTCCATTGGGTTCACCATTC-3′; reverse, 5′-GCGGCAGGTCTTAAGAGATGAG-3′), miR-613 (forward, 5′-CGTTTC TTCCTTGTAAGGA-3′; reverse, 5′-CCCAAGCTTGTTGGAGAACAGCAGCGAGGAC-3′), and SPOCK1 (forward, 5′-CACTGGGTTGGACCTTCGA-3′; reverse, 5′-CTTTGGTGGCTCAGGCTCT-3′), glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (forward, 5′-GACTCCACTCACGGCAAATTCA-3′; reverse, 5′-TCGCTCCTGGAAGATGGTGAT-3′), and U6 small nuclear RNA (snRNA) (forward: 5′-GCTCGCTTCGGCAGCACA-3′ and reverse: 5′-GAGGTATTCGCACCAGAGGA-3′). GAPDH served as an internal reference gene for UCA1 and SPOCK1. U6 snRNA was viewed as an internal control for miR-613. Relative expression was calculated using the 2−ΔΔCt method.
Western blot analysis
Total protein of GBC tissues and cells was extracted using RIPA protein extraction reagent (Beyotime, Shanghai, China). Total protein were isolated by electrophoresis on 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred onto the polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA, USA). The PVDF membranes were blocked in TBST buffer with 5% nonfat milk for 1 h. Then, the membranes were incubated with primary antibodies: anti-cleaved caspase-3 (ab32042, 1:1000; Abcam, Cambridge, MA, USA), anti-Bax (ab182733, 1:1000; Abcam), anti-Bcl-2 (ab32124, 1:1000; Abcam), anti-SPOCK1 (ab229935, 1:2000; Abcam), and anti-GAPDH (ab9485, 1:1000; Abcam). Subsequently, the membranes were incubated with a secondary antibody (goat anti-rabbit IgG [ab6721, 1:10,000; Abcam]). The immunoblot was visualized through the enhanced chemiluminescence solution (Beyotime).
Cell proliferation assay
The proliferation of NOZ and GBC-SD cells was evaluated by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) Kit (Sigma, St. Louis, MO, USA). Briefly, the transfected NOZ and GBC-SD cells (5000 cells/well) were seeded into 96-well plates for 24, 48, and 72 h. Then, MTT (20 μL, 5 mg/mL) was added to each well and kept for 4 h. Next, dimethyl sulfoxide (200 μL) was utilized for the dissolution of the formazan crystals. The optical density at 490 nm was measured using a Microplate Reader (Thermo Fisher Scientific).
Transwell assay
The migration of transfected NOZ and GBC-SD cells was evaluated through the transwell chamber (8 μm; BD Biosciences, San Jose, CA, USA). Briefly, the transfected NOZ and GBC-SD cells (5 × 104) were seeded to the upper of the chamber with RPMI 1640 medium, and the lower of the chamber was supplemented with RPMI 1640 containing FBS (10%). After culture for 24 h, the cells on the lower surface of the membrane were fixed through paraformaldehyde (4%, Sigma) and stained with Crystal Violet (0.1%, Solarbio, Beijing, China). The transwell chamber coated with Matrigel matrix (BD Biosciences) was used for invasion assay. The migrated or invaded cells were counted using the inverted microscope (Olympus, Tokyo, Japan).
Cell apoptosis assay
The apoptotic rate of transfected NOZ and GBC-SD cells was determined by the Annexin V-Fluorescein Isothiocyanate (FITC)/PI Apoptosis Detection Kit (BD Biosciences). Briefly, after transfection for 48 h, the transfected NOZ and GBC-SD cells (1 × 105) were resuspended in binding solution and then stained with Annexin V-FITC and PI. After incubating for 20 min in the dark, the apoptotic rate was determined using a FACScan® flow cytometry (BD Biosciences).
Dual-luciferase reporter assay
The starBase2.0 database was used to predict the binding sites between miR-613 and UCA1 or SPOCK1. The sequences of wild-type (WT) UCA1 (WT-UCA1), mutant (MUT) UCA1 (MUT-UCA1), WT SPOCK1 (SPOCK1 3′UTR-WT), mutant SPOCK1 (SPOCK1 3′UTR-MUT) were synthesized and inserted into the pMIR-REPORT vectors (Applied Biosystems, Foster, CA, USA). NOZ and GBC-SD cells were cotransfected with luciferase reporter vectors and miR-613 or miR-NC by Lipofectamine 2000 reagent, respectively. The luciferase activity was measured using the Luciferase Reporter Assay Kit (Promega) after transfection for 48 h.
RNA immunoprecipitation assay
The relationship between UCA1 and miR-613 was verified through the EZ-Magna RNA Immunoprecipitation (RIP) RNA-Binding Protein Immunoprecipitation Kit (Millipore, Billerica, MA, USA). NOZ and GBC-SD cells were lysed with RIP lysis buffer. Following this, the lysates were incubated with the RIP buffer containing magnetic beads conjugated with anti-IgG or anti-Ago2 antibodies (Millipore). Subsequently, the magnetic beads were treated with proteinase K (Sigma). The relative enrichment of UCA1 and miR-613 was analyzed by qRT-PCR.
Xenograft assay
Ten BALB/c nude mice (athymic, 3-week-old) were acquired from Shanghai Experimental Animal Center (Shanghai, China). The animal experiment was ratified by the Animal Ethics Committee of Loudi Central Hospital of Hunan. In short, NOZ cells (1 × 106) with lentivirus-mediated sh-UCA1 (RiboBio) or sh-NC were subcutaneously injected into the dorsal side of the nude mice. Ten mice were divided into two groups (5 mice/group), one group was injected with NOZ cells with sh-NC, and the other group was injected with NOZ cells with sh-UCA1. Tumor volume was figured through the equation: volume = (length × width 2 )/2. Tumor volume was measured every 5 d until day 35. After 5 weeks, the mice were euthanized for subsequent analysis.
Statistical analysis
All in vitro experiments were repeated at least three times. GraphPad Prism 5.0 (GraphPad, San Diego, CA, USA) and SPSS 22.0 software (SPSS, Chicago, IL, USA) were utilized for statistical analysis. Data were exhibited as mean ± standard deviation. Clinicopathological features were analyzed by chi-square tests. Person's correlation analysis was employed to determine the correlation between UCA1 and miR-613. Comparisons between two or among more groups were determined by Student's t test or one-way variance analysis (ANOVA). Differences with p < 0.05 were statistically significant.
Results
UCA1 was upregulated and miR-613 was downregulated in GBC tissues and cells
To verify the differential expression of UCA1 and miR-613 in GBC, the authors detected the expression of UCA1 and miR-613 in 30 pairs of GBC and adjoining normal tissues through qRT-PCR. The results presented that UCA1 expression was obviously upregulated in GBC tissues when compared with the adjoining normal tissues (Fig. 1A). As exhibited in Table 1, the high expression of UCA1 was associated with TNM stage and lymph node metastasis. Compared with adjoining normal tissues, miR-613 was lowly expressed in GBC tissues (Fig. 1B). Moreover, UCA1 expression was negatively correlated with miR-613 in GBC tissues (Fig. 1C). Compared with the HIBEpiC cells, UCA1 expression was signally upregulated while miR-613 expression was markedly downregulated in GBC cells (SGC-996, GBC-SD, and NOZ), especially in GBC-SD and NOZ cells (Fig. 1D, E). Thus, they selected GBC-SD and NOZ cells for follow-up investigation. These results indicated that the aberrant expression of UCA1 and miR-613 might be involved in the development of GBC.
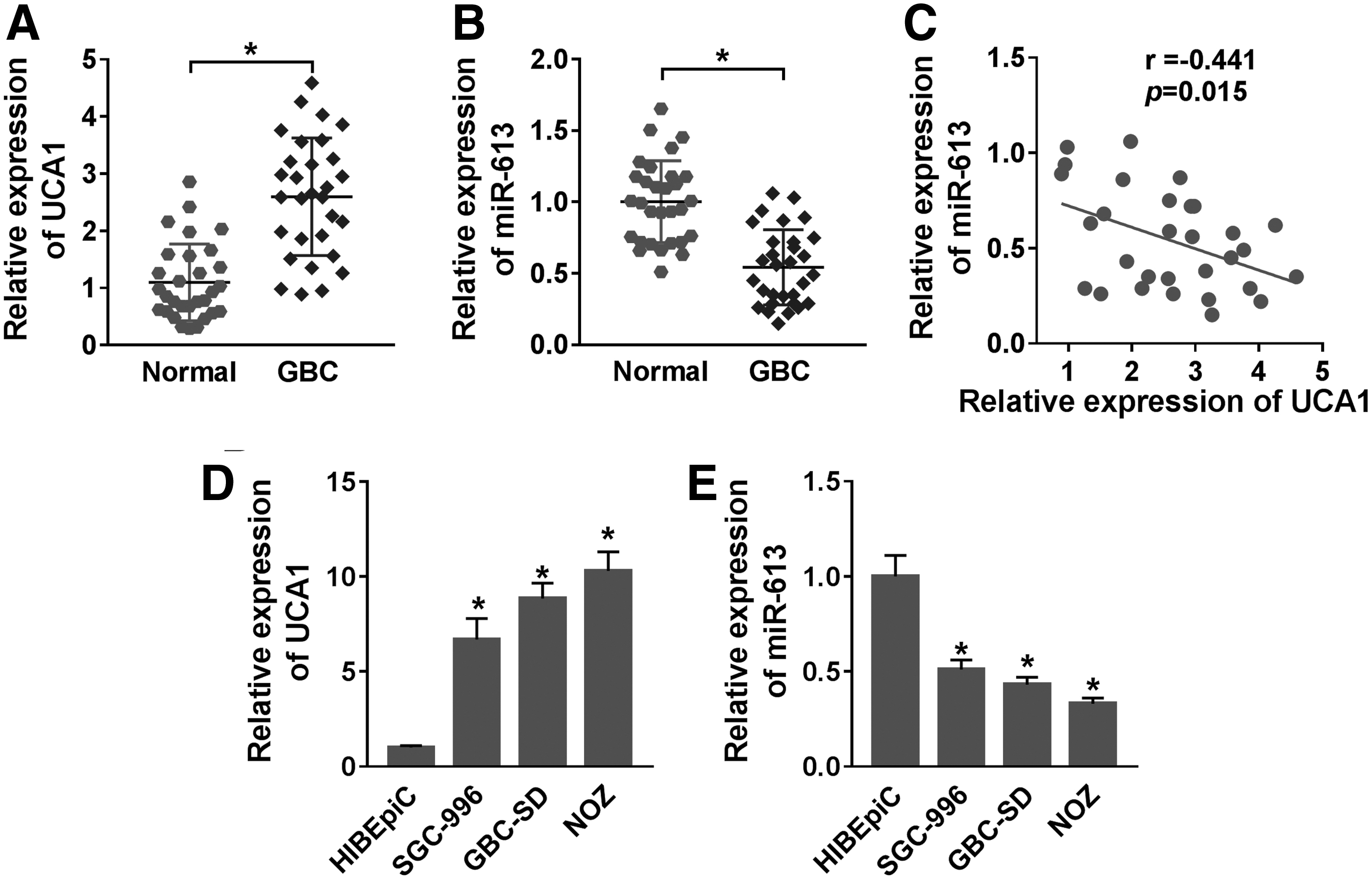
Expression levels of UCA1 and miR-613 in GBC tissues and cells.
Correlation of the Expression of Urothelial Carcinoma Associated 1 with Clinicopathological Features in Gallbladder Cancer Patients
TNM, tumor–node–metastasis; UCA1, urothelial carcinoma-associated 1.
Knockdown of UCA1 suppressed cell proliferation, migration, invasion, and promoted cell apoptosis in GBC cells
In consideration of the above results, they verified the role of UCA1 in GBC through loss-of-function experiments. The knockdown efficiency of UCA1 was displayed in Figure 2A. Moreover, MTT assays indicated that the proliferation of NOZ and GBC-SD cells were markedly repressed by UCA1 downregulation in comparison with the control group (Fig. 2B, C). Transwell assays manifested that the migration and invasion of UCA1-silenced NOZ and GBC-SD cells were curbed (Fig. 2D–G). Flow cytometry assays disclosed that the apoptotic rate of UCA1-suppressed NOZ and GBC-SD cells was conspicuously elevated (Fig. 2H, I). Also, Western blot analysis exhibited that UCA1 silencing downregulated the levels of Bax and cleaved-casp-3 and upregulated the level of Bcl-2 in NOZ and GBC-SD cells (Fig. 2J, K). These findings suggested that UCA1 knockdown inhibited cell proliferation, migration, invasion, and facilitated cell apoptosis in GBC cells.

Effects of UCA1 silencing on the proliferation, migration, invasion, and apoptosis of GBC cells. NOZ and GBC-SD cells were transfected with sh-UCA1 or sh-NC.
UCA1 severed as a sponge for miR-613 in GBC cells
To investigate the targeting relationship between UCA1 and miR-613 in GBC, they first predicted the potential binding sites between UCA1 and miR-613 using an online bioinformatics database (starBase2.0). The results exhibited that miR-613 had complementary bases paring with UCA1 (Fig. 3A). Subsequently, dual-luciferase reporter assay showed that miR-613 overexpression prominently impeded the luciferase activity of the WT-UCA1 reporter in NOZ and GBC-SD cells relative to the control group (Fig. 3B, C). Moreover, RIP assay presented that UCA1 and miR-613 were conspicuously enriched in Ago2-containing beads compared with IgG-harboring beads (Fig. 3D, E). Also, the expression of UCA1 was elevated in NOZ and GBC-SD cells after UCA1 transfection (Fig. 3F). In addition, miR-613 expression was repressed in UCA1-increased NOZ and GBC-SD cells and was increased in UCA1-silenced NOZ and GBC-SD cells (Fig. 3G, H). Collectively, these results manifested that miR-613 served as a target for UCA1 in GBC cells.
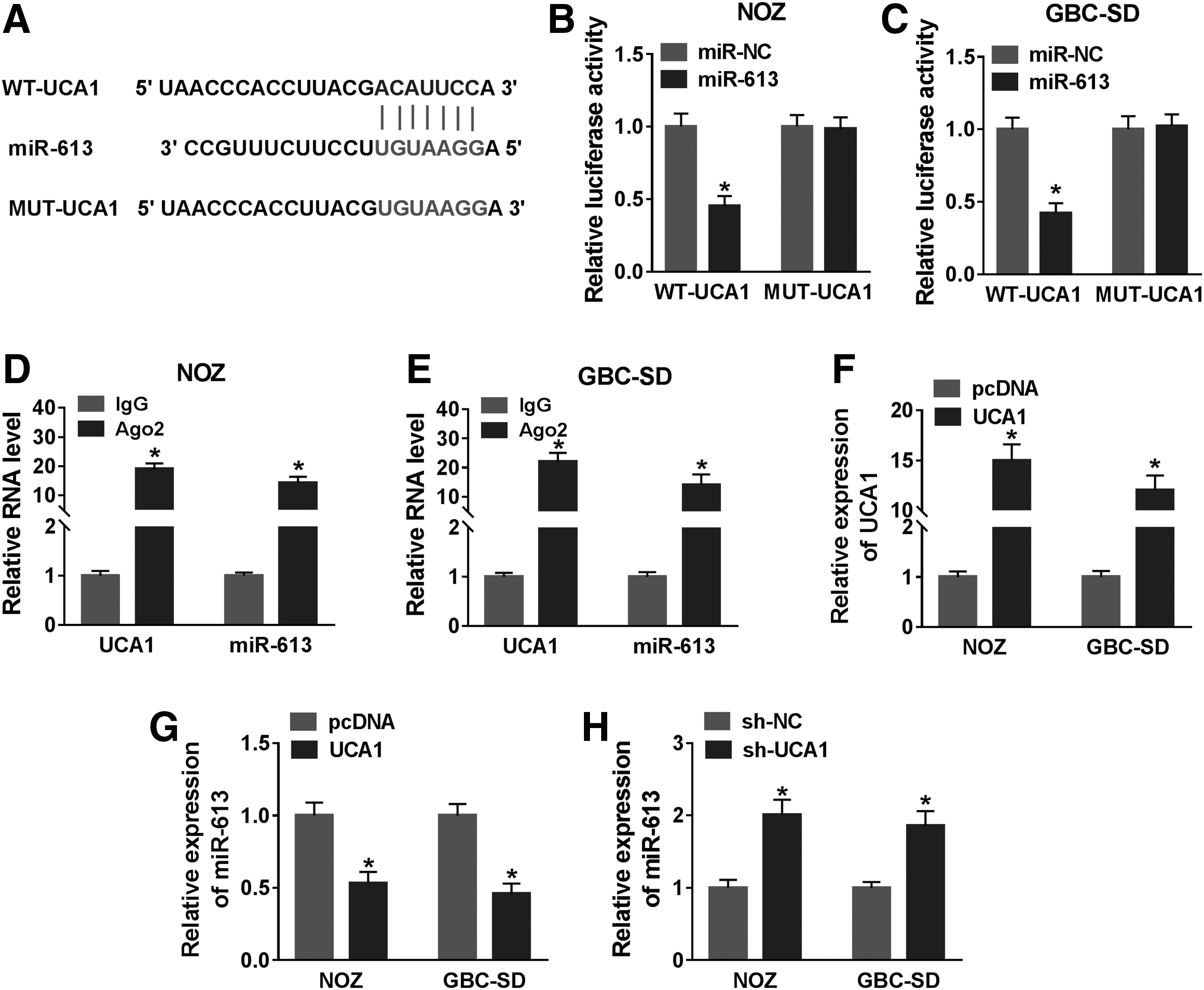
UCA1 served as a sponge for miR-613 in GBC cells.
UCA1 enhancement restored miR-613 upregulation-mediated effects on the proliferation, migration, invasion, and apoptosis of GBC cells
Next, they further explored the influence of UCA1 overexpression on the proliferation, migration, invasion, and apoptosis of GBC cells mediated by miR-613 mimic. qRT-PCR manifested that miR-613 expression was overtly increased in NOZ and GBC-SD cells after miR-613 mimic transfection (Fig. 4A). Moreover, miR-613 overexpression repressed proliferation, migration, and invasion of NOZ and GBC-SD cells, whereas this influence was overturned by UCA1 elevation (Fig. 4B–D). Also, the promoting effect of miR-613 upregulation on apoptosis of NOZ and GBC-SD cells was abolished by UCA1 overexpression (Fig. 4E). In addition, the upregulation of Bax and cleaved-casp-3 and the downregulation of Bcl-2 in NOZ and GBC-SD cells caused by miR-613 mimic were restored after UCA1 overexpression (Fig. 4F, G). These results showed that UCA1 regulated cell proliferation, migration, invasion, and apoptosis in GBC cells through miR-613.
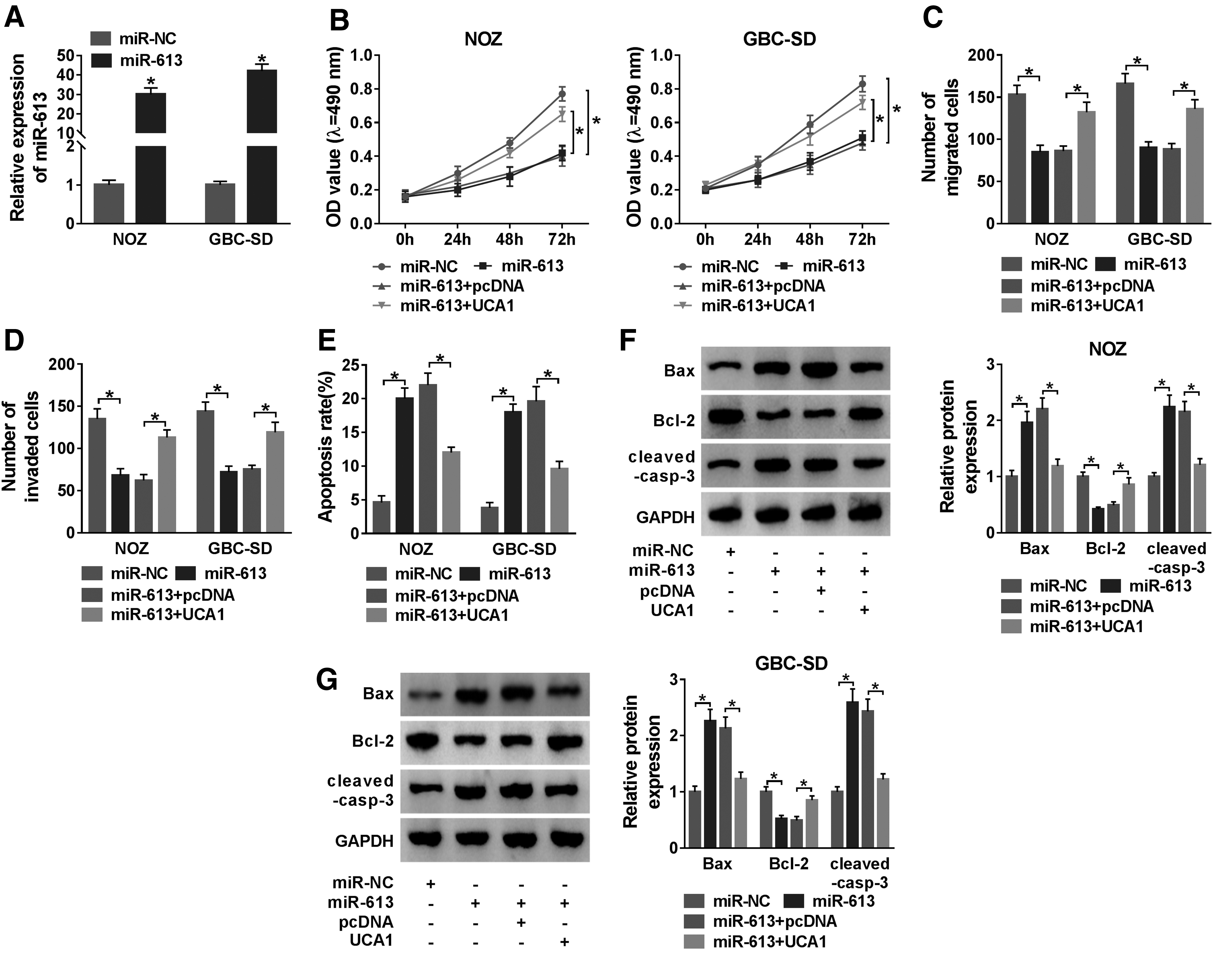
UCA1 upregulation reversed the impacts of miR-613 elevation on the proliferation, migration, invasion, and apoptosis of GBC cells.
SPOCK1 was verified as a target for miR-613 in GBC cells
In view of the above findings, they deduced that UCA1 might exert its function through acting as a ceRNA. As shown in Figure 5A, SPOCK1 was predicted as a target for miR-613. The luciferase activity of the SPOCK1 3′UTR-WT reproter in NOZ and GBC-SD cells transfected with miR-613 was effectively attenuated compared with the miR-NC, but there was no remarkable difference in the luciferase intensity of the SPOCK1 3′UTR-MUT reporter (Fig. 5B, C). Also miR-613 expression was reduced in NOZ and GBC-SD cells after anti-miR-613 transfection (Fig. 5D). Furthermore, miR-613 mimic reduced the level of SPOCK1 protein NOZ and GBC-SD cells, but miR-613 inhibitor had an opposite result (Fig. 5E, F). Additionally, SPOCK1 mRNA and protein levels were drastically upregulated in GBC tissues and cells (Fig. 5G–I). Taken together, SPOCK1 served as a downstream target for miR-613 in GBC cells.

SPOCK1 acted as a target for miR-613.
SPOCK1 overexpression reversed UCA1 inhibition-mediated effects on the proliferation, migration, invasion, and apoptosis of GBC cells
Given that UCA1 acted as a sponge for miR-613, which targeted SPOCK1 in GBC cells. They surveyed whether UCA1 exerted its function through SPOCK1 in GBC cells. Silenced UCA1 expression downregulated the level of SPOCK1 protein in NOZ and GBC-SD cells, but this decrease was abrogated by miR-613 inhibitor (Fig. 6A, B). Moreover, the level of SPOCK1 protein was elevated in NOZ and GBC-SD cells transfected with SPOCK1 compared with the negative control vector (Fig. 6C). They also observed that SPOCK1 elevation overturned the repression of proliferation, migration, invasion of NOZ and GBC-SD cells caused by UCA1 silencing (Fig. 6D–G). Also, elevated SPOCK1 expression overturned the facilitation of apoptosis of NOZ and GBC-SD cells mediated by UCA1 knockdown (Fig. 6H). Furthermore, the upregulation of SPOCK1 reversed UCA1 knockdown-mediated effects on the levels of Bax, cleaved-casp-3, and Bcl-2 in NOZ and GBC-SD cells (Fig. 6I, J). Collectively, these results showed that UCA1 modulated cell proliferation, migration, invasion, and apoptosis through regulating SPOCK1.
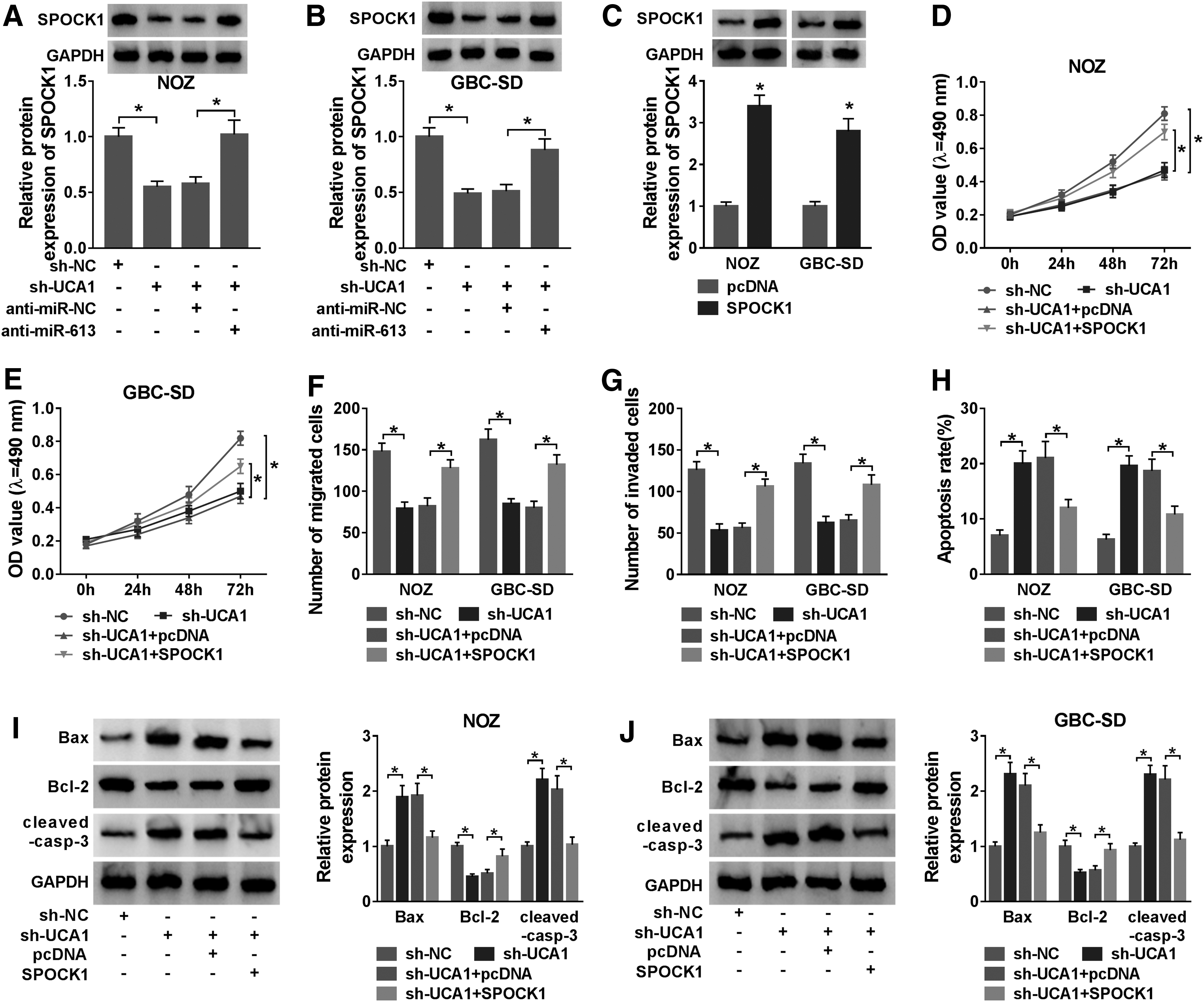
SPOCK1 elevation restored UCA1 silencing-mediated effects on the proliferation, migration, invasion, and apoptosis of GBC cells.
Silence of UCA1 reduced tumor growth in vivo
Knowing that UCA1 silencing could repress the malignant behaviors of GBC cells in vitro, they further verified the role of UCA1 in GBC in vivo through xenograft assay. They observed that UCA1 silencing evidently reduced tumor growth (volume) and weight in contrast to the control group (Fig. 7A, B). Also, UCA1 expression was decreased while miR-613 expression was elevated in mice tumor tissues in the sh-UCA1 group in contrast to the sh-NC group (Fig. 7C, D). Furthermore, the level of SPOCK1 protein was reduced in mice tumor tissues in the sh-UCA1 group compared with the control group (Fig. 7E). Altogether, these data indicated that UCA1 inhibition reduced tumor growth in vivo.
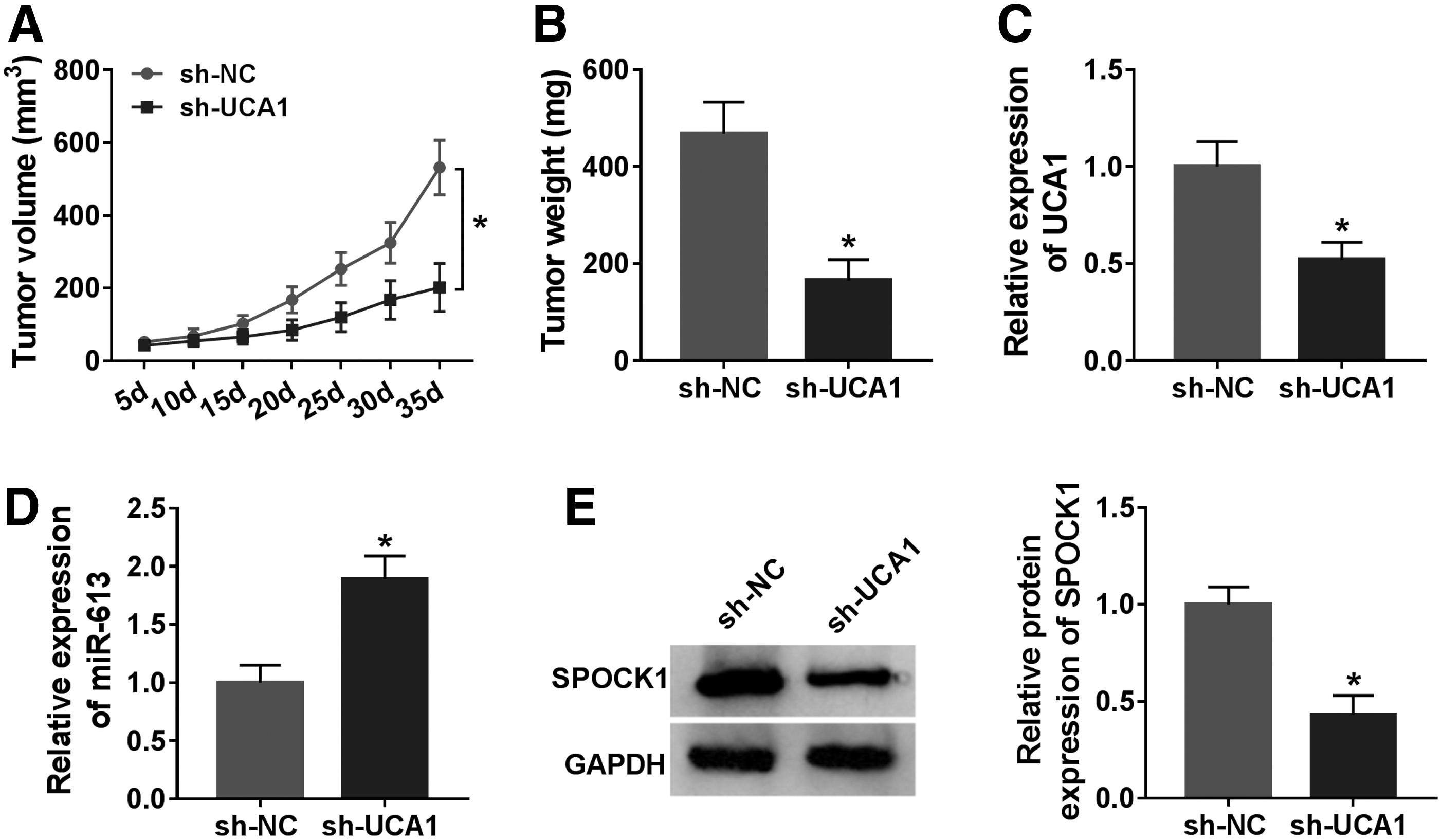
UCA1 silencing impeded tumor growth in vivo.
Discussion
At present, the specificity of GBC diagnosis is relatively low, and patients with advanced stage have poor prognosis and high mortality. It is well documented that lncRNAs and miRNAs are potential therapeutic targets for various cancers. 23 Herein, they aimed to explore regulatory mechanism between UCA1 and miR-613 in GBC. The results indicated that UCA1 exerted a promoting role in GBC progression. Knockdown of UCA1 repressed GBC progression through downregulation of SPOCK1 through sponging miR-613.
Convincing evidence showed that UCA1 played a vital role in the progression of various cancers. For example, UCA1 elevation promoted osteosarcoma progression and was correlated with poor prognosis of osteosarcoma patients. 24 Furthermore, UCA1 overexpression accelerated cell invasion and migration in pancreatic cancer through the hippo pathway. 25 Moreover, reduced UCA1 expression impeded cell invasion and proliferation through upregulating miR-206 in cervical cancer. 26 In the present study, UCA1 was upregulated in GBC tissues and cells. UCA1 knockdown decreased tumor growth in vivo and inhibited proliferation, migration, invasion, and promoted apoptosis of GBC cells in vitro. Previous study demonstrated that UCA1 promoted GBC advancement through recruiting enhancer of EZH2 and suppressing P21 and E-cadherin. 9 Therefore, these data manifested that UCA1 played a carcinogenic role in GBC.
Studies have shown that lncRNAs regulate gene expression by acting as miRNA sponges. 27 In the present study, UCA1 was proved as a sponge for miR-613 in GBC cells. It was reported that miR-613 mimic could repress Daam1 expression in triple-negative breast cancer, which impeded cell invasion and migration. 15 Yang et al. reported that lncRNA HOTAIR enhanced c-met expression through sponging miR-613, thereby facilitating tumor cell epithelial–mesenchymal transition in retinoblastoma. 28 However, Yang et al. pointed out that miR-613 accelerated colon cancer progression through repressing ATOH1 expression. 29 Also, miR-613 contributed to the advancement of cervical cancer through modulating PTPN9 expression. 30 Herein, miR-613 blocked cell proliferation, migration, invasion, and promoted cell apoptosis in GBC cells, but these impacts were restored by UCA1 overexpression. These indicated that UCA1 played its function through miR-613 in GBC.
In addition, they discovered that SPOCK1 was a downstream target for miR-613. Increasing evidence suggested that SPOCK1 was upregulated in multiple cancers. 19,21 Zhao et al. revealed that SPOCK1 silencing repressed cell invasion and proliferation through impeding the PI3K/AKT pathway. 31 Shu et al. also proved that SPOCK1 was upregulated in GBC tissues, and SPOCK1 overexpression promoted GBC progression through activation of the PI3K/AKT pathway. 22 In this study, SPOCK1 expression was elevated in GBC tissues and cells, and forced SPOCK1 expression overturned UCA1 silencing-mediated effects on proliferation, migration, invasion, and apoptosis of GBC cells. Furthermore, SPOCK1 was regulated by the UCA1/miR-613 axis. Therefore, they concluded that UCA1 modulated GBC progression through regulating SPOCK1 expression through miR-613.
Conclusion
In conclusion, UCA1 silencing impeded GBC advancement through downregulating SPOCK1 through sponging miR-613, implying that UCA1 might be a potential target for the treatment of GBC.
Availability of Data and Materials
The analyzed datasets generated during the present study are available from the corresponding author on reasonable request.
Footnotes
Authors' Contributions
All authors made substantial contribution to conception and design, acquisition of the data, or analysis and interpretation of the data; take part in drafting the article or revising it critically for important intellectual content; gave final approval of the revision to be published; and agree to be accountable for all aspects of the work.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.