
Abstract
Background:
To date, the clinical management of advanced hepatocellular carcinoma (HCC) patients remains challenging and the mechanisms of E2F transcription factor 1 (E2F1) underlying HCC are obscure.
Materials and Methods:
Our study integrated datasets mined from several public databases to comprehensively understand the deregulated expression status of E2F1. Tissue microarrays and immunohistochemistry staining was used to validate E2F1 expression level. The prognostic value of E2F1 was assessed. In-depth subgroup analyses were implemented to compare the differentially expressed levels of E2F1 in HCC patients with various tumor stages. Functional enrichments were used to address the predominant targets of E2F1 and shedding light on their potential roles in HCC.
Results:
We confirmed the elevated expression of E2F1 in HCC. Subgroup analyses indicated that elevated E2F1 level was independent of various stages in HCC. E2F1 possessed moderate discriminatory capability in differentiating HCC patients from non-HCC controls. Elevated E2F1 correlated with Asian race, tumor classification, neoplasm histologic grade, eastern cancer oncology group, and plasma AFP levels. Furthermore, high E2F1 correlated with poor survival condition and pooled HR signified E2F1 as a risk factor for HCC. Enrichment analysis of differentially expressed genes, coexpressed genes, and putative targets of E2F1 emphasized the importance of cell cycle pathway, where CCNE1 and CCNA2 served as hub genes.
Conclusions:
We confirmed the upregulation of E2F1 and explored the prognostic value of E2F1 in HCC patients. Two putative targeted genes (CCNE1 and CCNA2) of E2F1 were identified for their potential roles in regulating cell cycle and promote antiapoptotic activity in HCC patients.
Introduction
According to the latest cancer statistics, the incidence of liver cancer is increasing most rapidly compared with any other forms of tumor in America. 1 Globally, liver cancer ranks sixth among the most common cancers and fourth among the causes of cancer-related deaths. 2 Hepatocellular carcinoma (HCC) comprises more than three-fourths of liver cancer cases and threatens lives of people in most regions of the world, especially eastern Asia. 2 At present, the diagnosis of HCC predominantly relies on histological analysis and contrast-enhanced computed tomography (CT). 3 Surgery, chemotherapy, and radiotherapy are conventional treatment regimens for HCC. 4 –9 Regarding progressed HCC patients, chemotherapy is considered first-line care. 10 Nevertheless, a large body of HCC patients are asymptomatic in the early stage and the situation confronting advanced HCC patients remain grave owing to agent resistance and other uncontrollable factors. 11,12 Therefore, more effective markers are urgently needed to promote the screening of early-stage HCC patients, and more potent therapeutic methods in line with the concept of precision medicine are encouraged to optimize clinical management and extend life expectancy of advanced HCC patients.
E2F family transcriptional factors, whose activity depends on the phosphorylation status of retinoblastoma (RB), are imperative for G1-S phase transition in cell cycle and act as a trigger in tumor cells proliferation. 13,14 As a result of evolutionary adaptation to malignancies, E2F transcription factor 1 (E2F1) serves as an activator in regulating downstream targets, mediating both cell proliferation and apoptosis process. 15 Several small sample size studies reported the upregulation of E2F1 in HCC patients. 13,16,17 The upregulation of E2F1 not only drives HCC in a dose-dependent manner and facilitates tumor growth, but also correlates to poor outcomes of HCC patients. 18 –21 However, the role that E2F1 plays in HCC remains controversial, and the clinical implication of E2F1 in HCC has not been fully evaluated thus far. Some researchers support that upregulated E2F1 expression in HCC expedites tumor cells apoptosis. 22 On the contrary, others have it that E2F1 acts as an important antiapoptotic factor in liver cancer by attenuating c-Myc-mediated cell apoptosis. 23 Moreover, genetic alteration landscapes and the mechanisms of E2F1 underlying HCC are obscure, and the transcriptional targets of E2F1 in HCC were largely unknown. In colorectal cancer, E2F1 promote cell cycle by activating its targeted gene DNAJA1 (DnaJ heat shock protein family member A1). 24 In addition, it was reported that E2F1 activated lncRNA small nucleolar RNA host gene 3, thus promoting the proliferation and migration of nonsmall cell lung cancer cells. 25 Nonetheless, the transcriptional targets of E2F1 in HCC have not been intensively studied. More studies on E2F1 are required to dig out the underlying causes of HCC and to identify novel therapeutic targets in-depth.
Given the aforementioned situation, we sought to comprehensively understand the deregulated expression status of E2F1 in HCC patients at the mRNA and protein levels. In addition, the prognostic value of E2F1 in HCC patients was estimated. More importantly, we were devoted to addressing the predominant targets of E2F1 and to shedding light on their potential roles in HCC.
Materials and Methods
MRNA expression levels and potential clinical applications of E2F1 in HCC tissues by in silico analysis
Overall expression status of E2F1 in HCC patients compared with non-HCC controls
In this study, microarrays and mRNA-sequencing (RNA-seq) data submitted before February 5, 2020 were searched and downloaded from public databases, namely Gene Expression Omnibus (GEO), The Cancer Genome Atlas (TCGA), and the Genotype-Tissue Expression (GTEx), using the following search query: (tumor OR carcinoma OR adenocarcinoma OR neoplasm OR cancer) AND (hepatocellular OR hepar OR liver OR hepatic OR HCC). Duplicated dataset was removed and the latest version was preserved. The initially screened datasets were carefully assessed for eligibility. The enrolled datasets satisfied the standards as follows: (i) the organism should be Homo sapiens; (ii) the experimental specimens should be tissues dissected from HCC patients rather than cell lines; (iii) the matrices should contain expression values of E2F1; and (iv) non-HCC control group should be set up to be compared with HCC patients. Other datasets were excluded because the experimental group could not be judged whether it was HCC or intrahepatic cholangiocarcinoma without sufficient information.
A standardized mean difference (SMD) statistic was calculated based on the included datasets using STATA v12.0. Randomized-effect model was selected in case significant heterogeneity existed (I 2 > 50%). Otherwise, fixed-effect model was considered. Sensitive analysis was applied to probe the source of heterogeneity. We also exploited subgroup analysis to compare the differentially expressed levels of E2F1 in HCC patients from different perspectives. Begg's funnel plot was drawn to determine whether significant publication bias existed (p < 0.05). If publication bias could not be ignored, trim and fill method was used. Summary receiver operating characteristic (sROC) curve along with area under the curve (AUC) were generated and sensitivity, specificity, diagnostic likelihood ratio positive (DLR P), diagnostic likelihood ratio negative (DLR N), diagnostic score, and diagnostic odd ratio (DOR) were calculated to evaluate the discriminatory value of E2F1 in HCC patients. An AUC value between 0.7 and 0.9 and a DOR value >1 indicated moderate discriminatory capacity.
Associations between E2F1 expression and clinicopathological parameters
Clinicopathological information of HCC patients based on TCGA cohort was queried and downloaded using TCGAbiolinks package in R v3.6.1 software. HCC patients were divided into different groups according to their responding clinicopathological parameters. The expression levels of E2F1 in HCC patients with various characteristics were compared.
Prognostic value of E2F1 in HCC based on microarrays, RNA-seq data, and literature extraction
Survival analysis was performed using survival package in R v3.6.1. KM curve was plotted and log-rank test was conducted to judge whether the survival condition varied from low to high E2F1 expression groups in HCC patients. Literatures published up to February 15, 2020 were retrieved from databases inclusive of PubMed, Highwire, Wiley Online Library, EBSCO, Springer, ScienceDirect, and several Chinese literatures databases, namely Chinese Biological Medical Database (CBMdisc), China National Knowledge Infrastructure (CNKI), ChongQing VIP, WanFang, and ChaoXing. The search query was as follows: (tumor OR carcinoma OR adenocarcinoma OR neoplasm OR cancer) AND (hepatocellular OR hepar OR HCC OR liver OR hepatic) AND (E2F1 OR E2F-1 OR RBAP1 OR RBBP3 OR RBP3). Studies were included if: (i) HR and responding 95% confident interval (CI) of E2F1 in HCC patients were provided; or (ii) KM curve of high and low E2F1 groups in HCC patients was plotted and the data could be extracted using Engauge Digitizer. Other studies were excluded because of (i) lack of sufficient data on the prognostic value of E2F1 in HCC patients; (ii) articles as review, report, news, or conference abstracts. Eventually, the effect size HR data were pooled using STATA12.0.
Potential molecular mechanisms of E2F1 underlying HCC by gene expression analysis
Evaluation of E2F1 genetic alterations and mutation types in HCC patients
Genetic alteration landscapes of E2F1 in HCC patients were addressed using cBioPortal for Cancer Genomics. E2F1 alterations were queried based on 360 HCC patients (TCGA, Firehose Legacy), and a method of mRNA Expression z-Scores (RNA Seq V2 RSEM) was applied, setting a z-score threshold at ±2.0. In addition, the relevance between E2F1 RNA Seq V2 RSEM and methylation was further probed. The overall and disease/progression-free survival condition between E2F1 altered and unaltered groups was compared, respectively. Furthermore, E2F1 mutation types in HCC patients were clarified with the aid of the Catalogue of Somatic Mutations in Cancer (COSMIC). mSignatureDB was used to learn the mutation landscapes of E2F1in HCC.
Identification of DEGs, CEGs, and putative targets of E2F1
Based on the microarrays and RNA-seq data, differentially expressed genes (DEGs) in HCC patients, meeting |log2Fold Change| > 1 with an adjusted value of p < 0.05, were obtained using Limma Voom package in R software v3.6.1. Coexpressed genes (CEGs) of E2F1 were identified by calculating Pearson's correlation coefficients using R software v3.6.1. E2F1 positively related CEGs possessed Pearson's correlation coefficients >0.3 (p < 0.05). On the contrary, E2F1 negatively related CEGs possessed Pearson's correlation coefficients less than −0.3 (p < 0.05). Putative targets of transcriptional factor E2F1 were downloaded from Cistrome Data Browser (Cistrome DB), filtered by scores >2.0. Eventually, upregulated DEGs, E2F1 positively related CEGs, and putative targets of E2F1 were intersected. Downregulated DEGs, E2F1 negatively related CEGs, and putative targets of E2F1 were also intersected for further mechanism exploration.
Functional analysis of E2F1 underlying HCC by GO, KEGG, DO, and PPI
The intersected genes were utilized to perform Gene Ontology (GO), the Kyoto Encyclopedia of Genes and Genomes (KEGG), and Disease Ontology (DO) enrichment analyses using clusterProfiler package in R software v3.6.1, respectively. An adjusted value of p < 0.05 signified statistical significance. Protein-to-protein internet (PPI) was constructed based on the significantly clustered pathways using STRING. Hub genes were identified using Cytoscape v3.6.1. Hub genes were checked further according to chromatin-immunoprecipitation coupled with sequencing (ChIP-seq) peak intensities from Cistrome DB. Promoter sequences in the hub genes matched to the motifs of E2F1 were also identified using the MEME Suite. 26
Protein expression level of E2F1 in HCC by IHC
A total of 315 HCC specimens and 194 non-HCC tissues were purchased from Pantomics, Inc. (Richmond, CA). This study was approved by ethics committee of The First Affiliated Hospital of Guangxi Medical University, Nanning, People's Republic of China. Recombinant anti-E2F1 antibody was purchased from Abcam website (Cambridge, MA). The streptavidin–peroxidase method was used for immunohistochemical staining. Two professional pathologists took double-blind readings without any recognizable clinical information of each specimen. Total 20 high magnification fields (400 × ) were chosen for each slice to observe and determine the staining intensity. The tissue microarray manufacturing process and immunohistochemical staining method followed the instructions of manufacturers.
Statistical analysis
Independent-samples t-test was used to determine the difference of E2F1 expression between HCC and non-HCC patients. One-way analysis of variance (ANOVA) was selected to compare E2F1 levels among three or more groups with clinicopathological parameters. Unless otherwise stated, the statistical results are presented as mean (M) ± standard deviation (SD). A value of p < 0.05 indicates statistical significance.
Results
Elevated expression of E2F1 in HCC patients compared with non-HCC controls based on microarrays and RNA-seq data
The details about population study, including quality control, population structure, inclusion criteria, and exclusion criteria are given in Supplementary Figure S1. The basic information of the enrolled datasets is given in Table 1. Because high heterogeneity existed (I 2 = 90.4%, p < 0.001), randomized-effect model was exploited. An SMD value of 1.11 (95% CI: 0.92–1.31) (Fig. 1) signified significant elevated E2F1 expression in liver tissues of HCC patients compared with that of control groups (i.e., normal, hepatitis, cirrhotic, dysplastic, or para-carcinoma liver tissues). Sensitive analysis indicated that the difference among the included datasets could be ignored (Supplementary Fig. S2A). No significant publish bias existed (Supplementary Fig. S2B). The upregulation levels of E2F1 in HCC patients were independent of age (Supplementary Fig. S3A), gender (Supplementary Fig. S3B), ethnicity (Supplementary Fig. S4), Barcelona Clinic liver cancer (BCLC) class (Supplementary Fig. S5A), tumor recurrent status (Supplementary Fig. S5B), TNM tumor stage (Supplementary Fig. S5C), and specimen types of control groups (Supplementary Fig. S6), because E2F1 was significantly upregulated in all those subgroups. sROC curve indicated a moderate discriminatory capacity of E2F1 in differentiating HCC patients from non-HCC controls (Figs. 2 and 3).
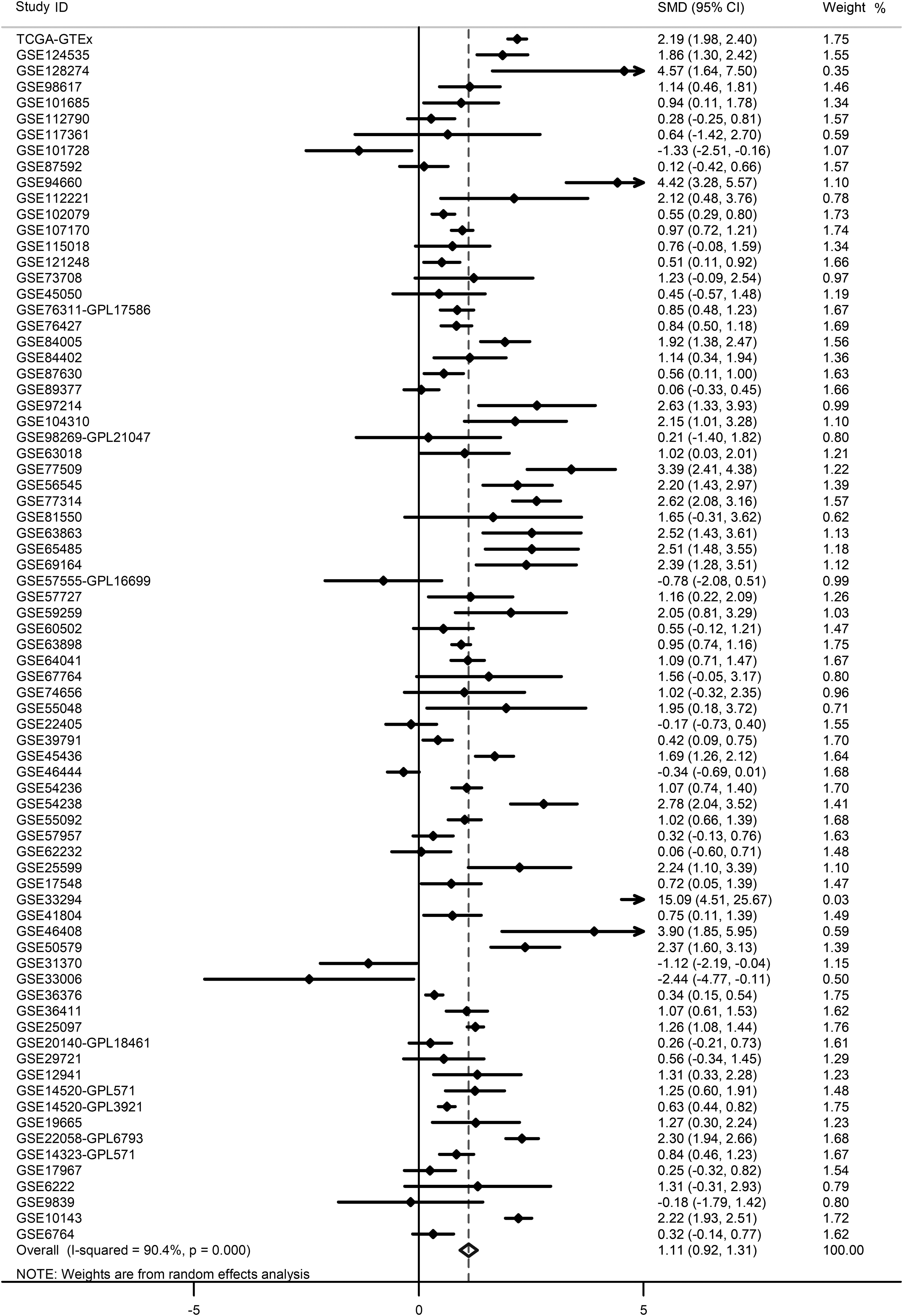
SMD of E2F1 expression level in HCC and non-HCC groups. Forest plot indicated an elevated trend of E2F1 expression in HCC patients compared with non-HCC controls. SMD, standard mean deviation; HCC, hepatocellular carcinoma.
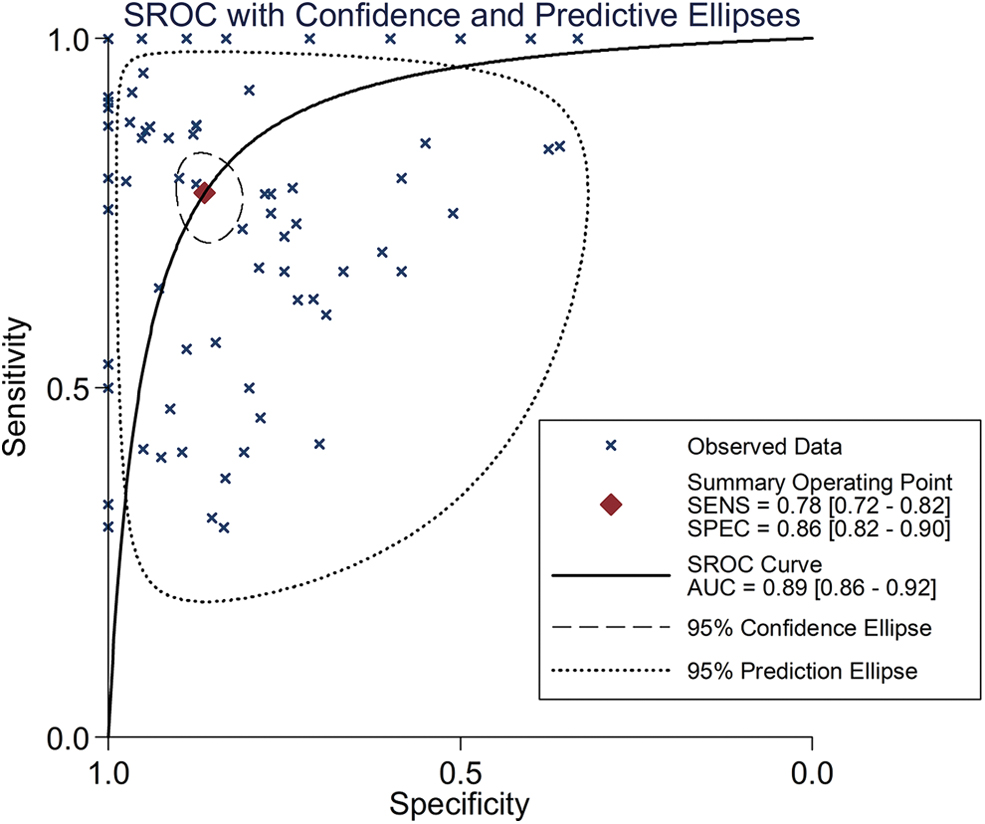
SROC curve. An AUC of 0.89 indicates moderate discriminatory capacity of E2F1 in differentiating HCC patients from non-HCC controls. SROC, summary receiver operating characteristic; AUC, area under the curve. Color images are available online.
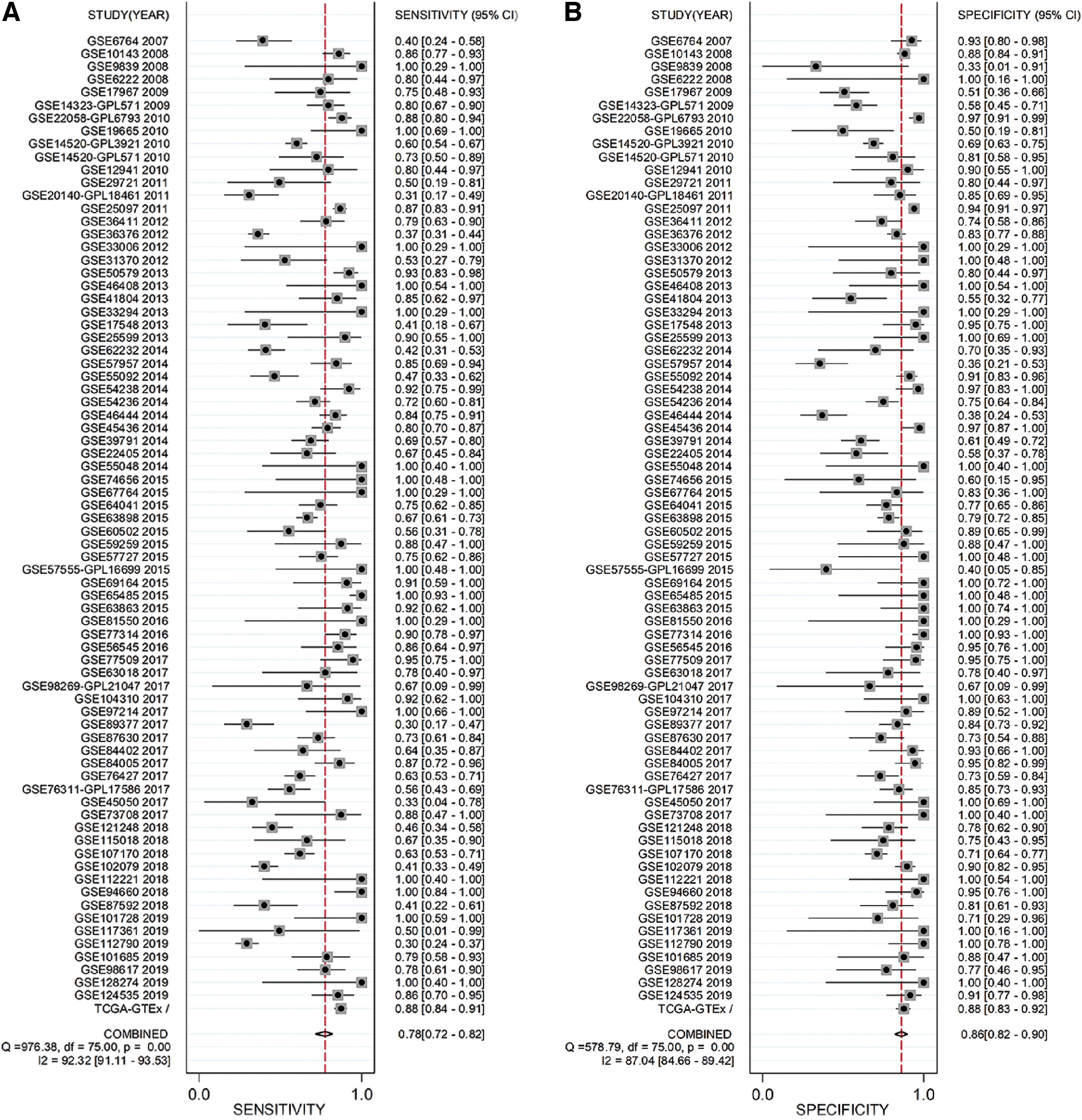
Calculation of sensitivity, specificity, DLR P, and DLR N. Panels

Basic Information of the 76 Enrolled Datasets
Associations between E2F1 expression and clinicopathological parameters
The relevance between E2F1 expression and several clinical parameters of HCC patients is given in Table 2. E2F1 expression level in HCC patients correlated with race (p = 0.001) (Fig. 4A), relative family cancer history (p = 0.043) (Fig. 4B), pathological stage (p < 0.001) (Fig. 4C), tumor classification (p < 0.001) (Fig. 4D), neoplasm histologic grade (p < 0.001) (Fig. 4E), eastern cancer oncology group (p = 0.018) (Fig. 4F), and AFP levels (ng/mL; p < 0.001) (Fig. 4G). Of interest, elevated E2F1 in HCC patients was relevant to Asian race, higher tumor classification, higher neoplasm histologic grade, higher eastern cancer oncology group, and higher plasma AFP levels.
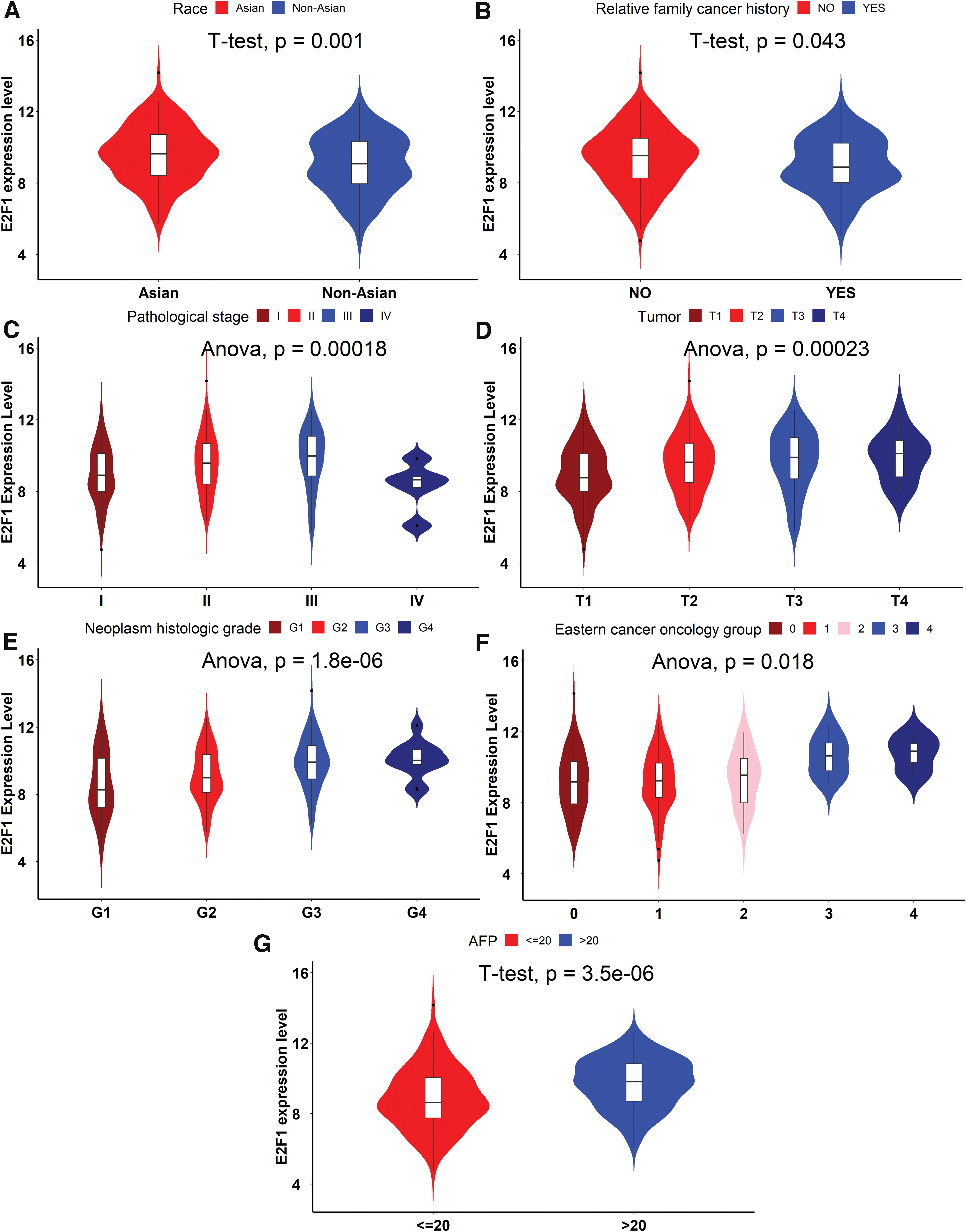
The relevance between E2F1 expression and clinical parameters of HCC patients. E2F1 expression level in HCC patients correlated with
The Relevance Between E2F1 Expression and Clinicopathological Parameters of Hepatocellular Carcinoma Patients
Independent samples t test was performed to identify the statistical significance of differences between two groups, whereas one-way analysis of variance (ANOVA) was selected for three or more groups.
A value of p < 0.05 signifies statistical significance. Of note, not all HCC patients had sufficient information.
E2F1 as a prognostic biomarker associated with poor outcomes of HCC patients based on microarrays, RNA-seq data, and literature extraction
KM curve based on TCGA cohort indicated that the survival condition of high E2F1 group was poorer in comparison with low E2F1 group for HCC patients (p = 0.044) (Fig. 5C). Four literatures studies and six datasets were included to conduct quantitative synthesis. 18,27 –29 Pooled HR indicated E2F1 as a risk factor in HCC patients (pooled HR = 1.61, 95% CI: 1.41–1.84) (Fig. 6A). Begg's funnel plot exhibited no significant publication bias (Fig. 6B) Sensitive analysis indicated no significant difference among the included studies (Fig. 6C).

Kaplan–Meier (KM) curve based on two microarrays and The Cancer Genome Atlas (TCGA) cohort.
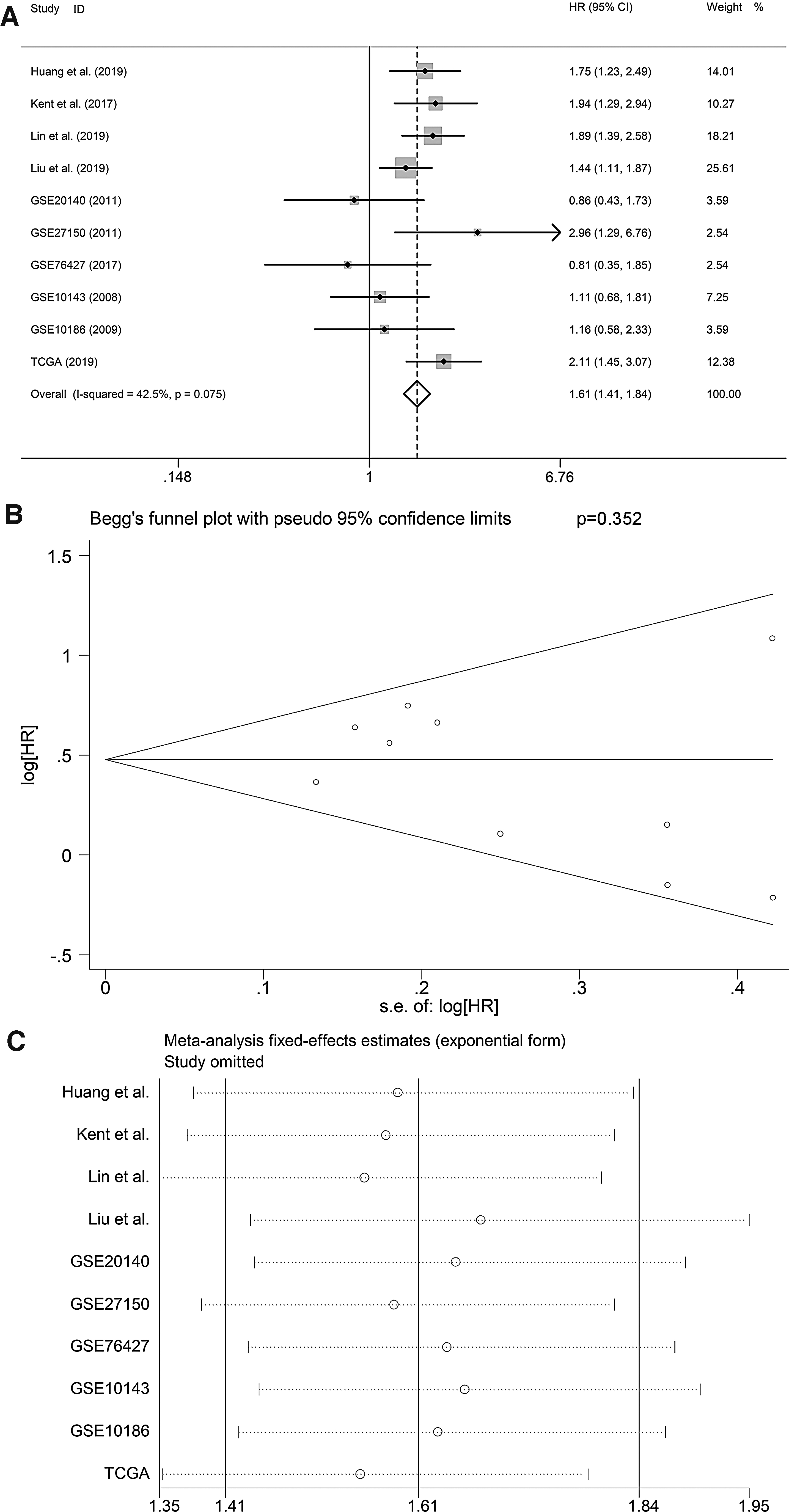
Pooled HR based on the enrolled literatures and three datasets.
E2F1 genetic alterations and mutation types in HCC patients
According to the results from cBioPortal for Cancer Genomics, E2F1 was altered in 18 (5%) HCC patients (TCGA, Firehose Legacy), among which mRNA high was the predominant genetic alteration, followed by amplification (Supplementary Fig. S7A). In addition, E2F1 mRNA expression was negatively correlated with E2F1 methylation (Supplementary Fig. S7B). No significant difference was established in the overall and disease/progression-free survival condition between E2F1 altered and unaltered groups in HCC patients (Supplementary Fig. S7C, D). The mutation landscapes of HCC patients are given in Supplementary Figure S7E. Substitution-coding silent and substitution missense were dominant mutation types of E2F1 in HCC patients based on COSMIC (Table 3).
Mutation Types of E2F1 in Hepatocellular Carcinoma Patients Based on the Catalog of Somatic Mutations in Cancer
Identification of DEGs, CEGs, and putative targets of E2F1
A total of 1478 upregulated and 2267 downregulated DEGs, all of which appeared in no less than 8 datasets, were identified in HCC patients. In addition, a total of 55242 E2F1 positively related CEGs and 26501 E2F1 negatively related CEGs were acquired. Supplementary Figure S8A and S8B showed the top 10 E2F1 positively related and E2F1 negatively related CEGs of GSE14323-GPL5474 and GSE107170, respectively. A total of 9631 putative targets of transcriptional factor E2F1 were obtained from Cistrome DB (all with scores >2.0). Subsequently, 373 overlapping genes of upregulated DEGs, E2F1 positively related CEGs, and putative targets of E2F1 were identified, all of which appeared in no less than 2 ChIP-seq results. Similarly, 409 overlapping genes of downregulated DEGs, E2F1 negatively related CEGs, and putative targets of E2F1 were acquired, all of which also appeared in no less than 2 ChIP-seq results. The CEGs network is given in Supplementary Figure S9.
Potential mechanisms of E2F1 in HCC onset and progression by targeting CCNE1 and CCNA2
Further functional enrichment analysis indicated that the 373 overlapping genes of upregulated DEGs, E2F1 positively related CEGs, and putative targets of E2F1 were dominantly clustered in DNA replication, cell cycle G1/S phase transition, and regulation of transcription involved in G1/S transition of mitotic cell cycle in GO BP term (Fig. 7A and Table 4). Chromosomal region and DNA-binding transcription activator activity (RNA polymerase II specific) was the most enriched GO CC and MF term, respectively. In addition, cell cycle was the most clustered KEGG pathways, followed by other glycan degradation and DNA replication pathway (Fig. 7B and Table 4). As for DO enrichment analysis, the 373 overlapping genes dominantly participated in different kinds of cancer diseases (Fig. 7C and Table 4). On the contrary, 409 intersecting genes of E2F1 negatively related CEGs, downregulated DEGs, and putative target genes of E2F1 were found to be involved in several synaptic components in GO CC terms (Fig. 7D and Table 5), namely glutamatergic synapse, postsynaptic specialization membrane, and intrinsic component of synaptic membrane. Nonetheless, those genes were not significantly clustered in KEGG and DO enrichment analyses; therefore, the results were not presented.
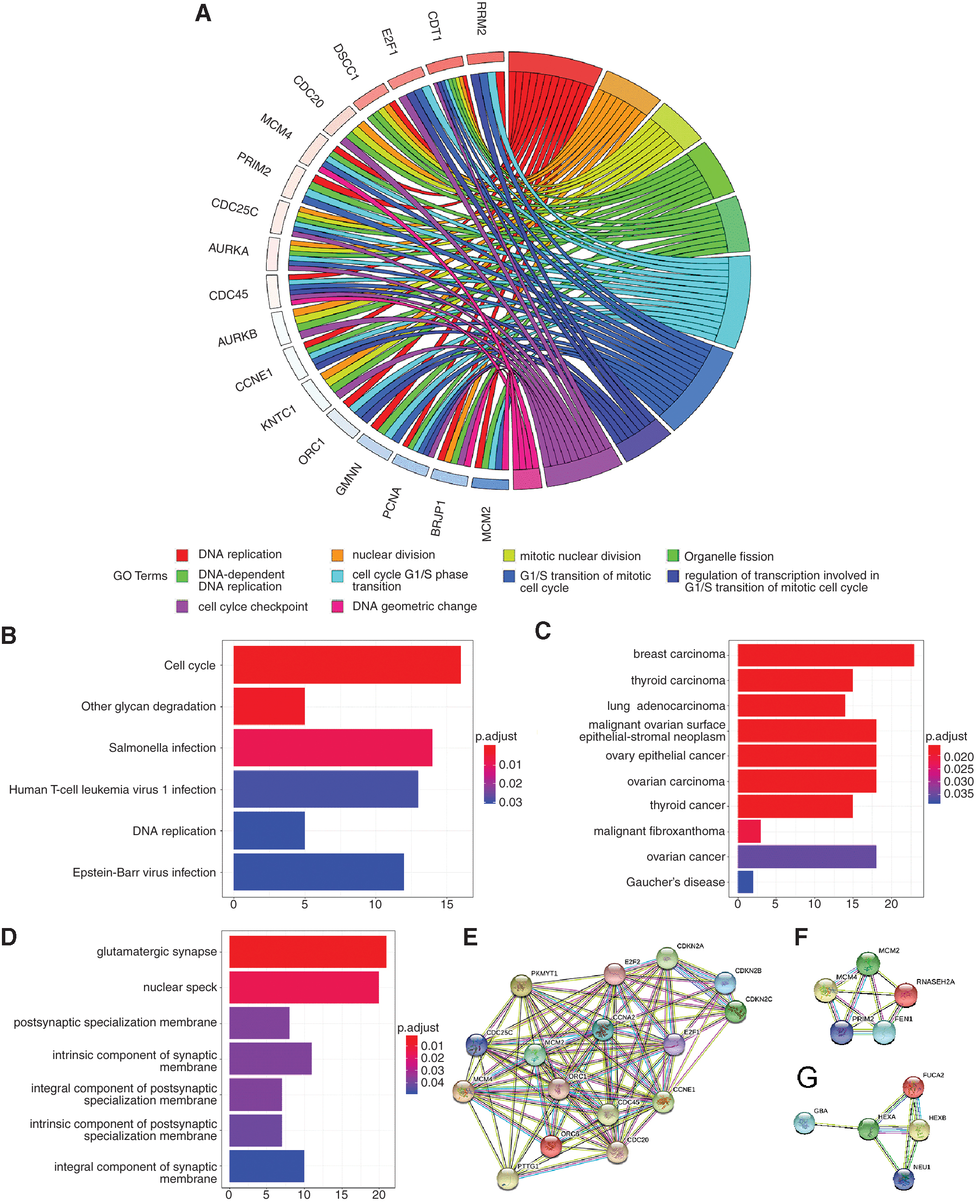
Enrichment analyses.
Enrichment Analysis Based on the 373 Intersecting Genes of Coexpressed Genes Positively Related to E2F1, Upregulated Differentially Expressed Genes, and Putative Target Genes of E2F1
Enrichment Analysis Based on the 409 Intersecting Genes of E2F1 Negatively Related Coexpressed Genes, Downregulated Differentially Expressed Genes, and Putative Target Genes of E2F1
These genes were not significantly clustered in terms of the Kyoto Encyclopedia of Genes and Genomes (KEGG) and Disease Ontology (DO) enrichment analysis, therefore, the results was not presented.
Subsequently, PPI were constructed based on the top significantly enriched KEGG pathways. Two hub genes (i.e., CCNE1 and CCNA2) were identified in the pathway of cell cycle (Fig. 7E) and the relationships between E2F1 and CCNE1 or CCNA2 are given in Figure 8. In the DNA replication pathway, FEN1, MCM2, MCM4, PRIM2, and RNASEH2A were recognized as hub genes (Fig. 7F). HEXA was identified as hub gene in the other glycan degradation pathway (Fig. 7G). Furthermore, two hub genes (i.e., CCNE1 and CCNA2) identified from cell cycle pathway as important putative targets of E2F1 were checked according to ChIP-seq peak intensities from Cistrome DB, and the results confirmed CCNE1 to be an essential target gene of E2F1 (Fig. 9). Moreover, we found that promoter sequences of CCNA2 possessed a region that highly matched to the motif of E2F1 (motif ID: MA0024.2; sequence name: NC_000004.12:c121825883-121823784) by using the MEME Suite, and the matched sequence was GGGGCGGGAGA (p < 0.0001, q = 0.005). Eventually, the prognostic value of hub genes in essential KEGG pathways were probed using online tool Gene Expression Profiling Interactive Analysis (GEPIA) (Fig. 10), which is based on TCGA cohort. Consistent with E2F1, higher expression level of CCNE1, CCNA2, FEN1, MCM2, MCM4, PRIM2, and RNASEH2A correlated with poor outcomes of HCC patients.
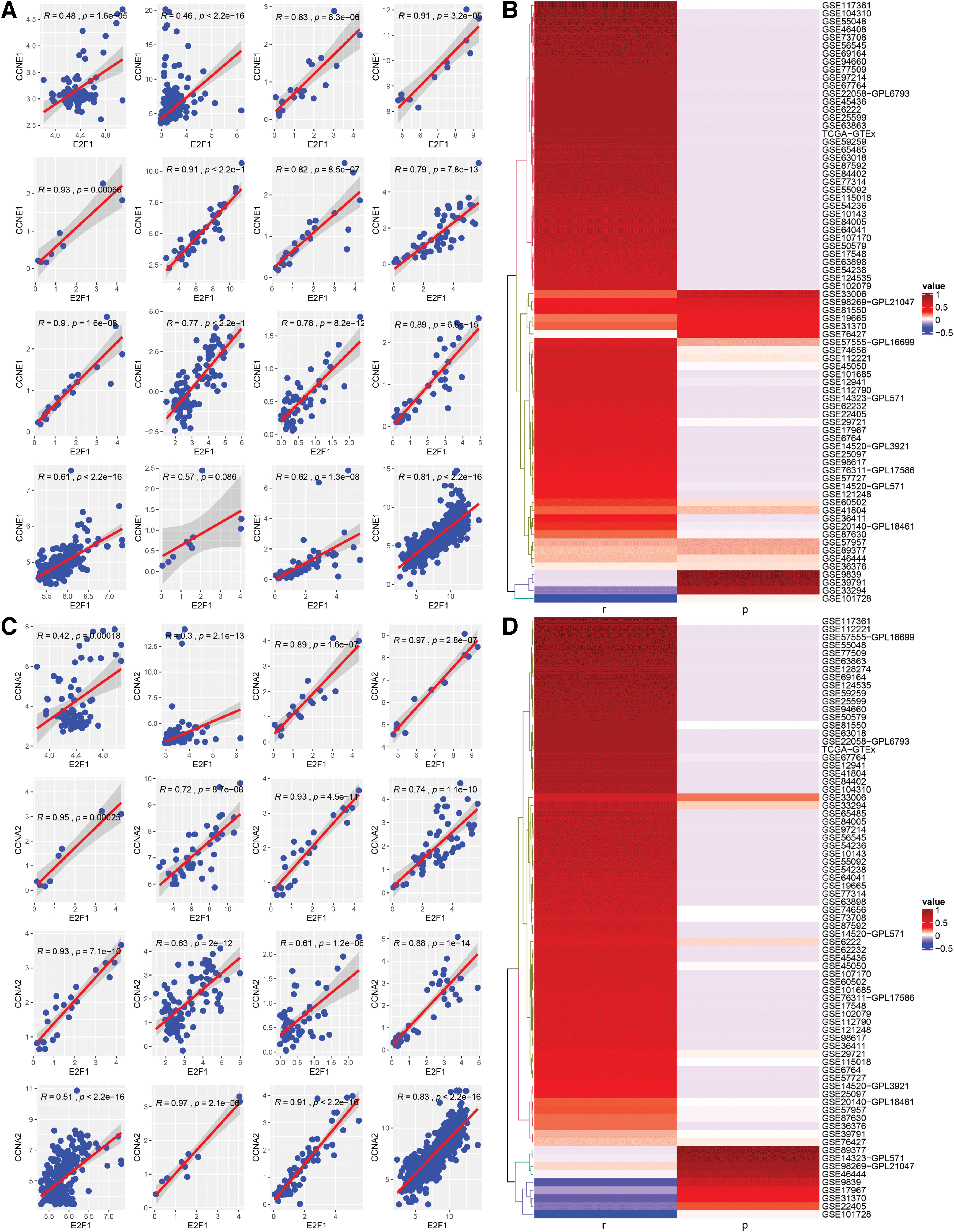
Relevance between E2F1 and two predominant putative targets (i.e., CCNE1 and CCNA2).

CCNE1 served as an essential putative target of E2F1.
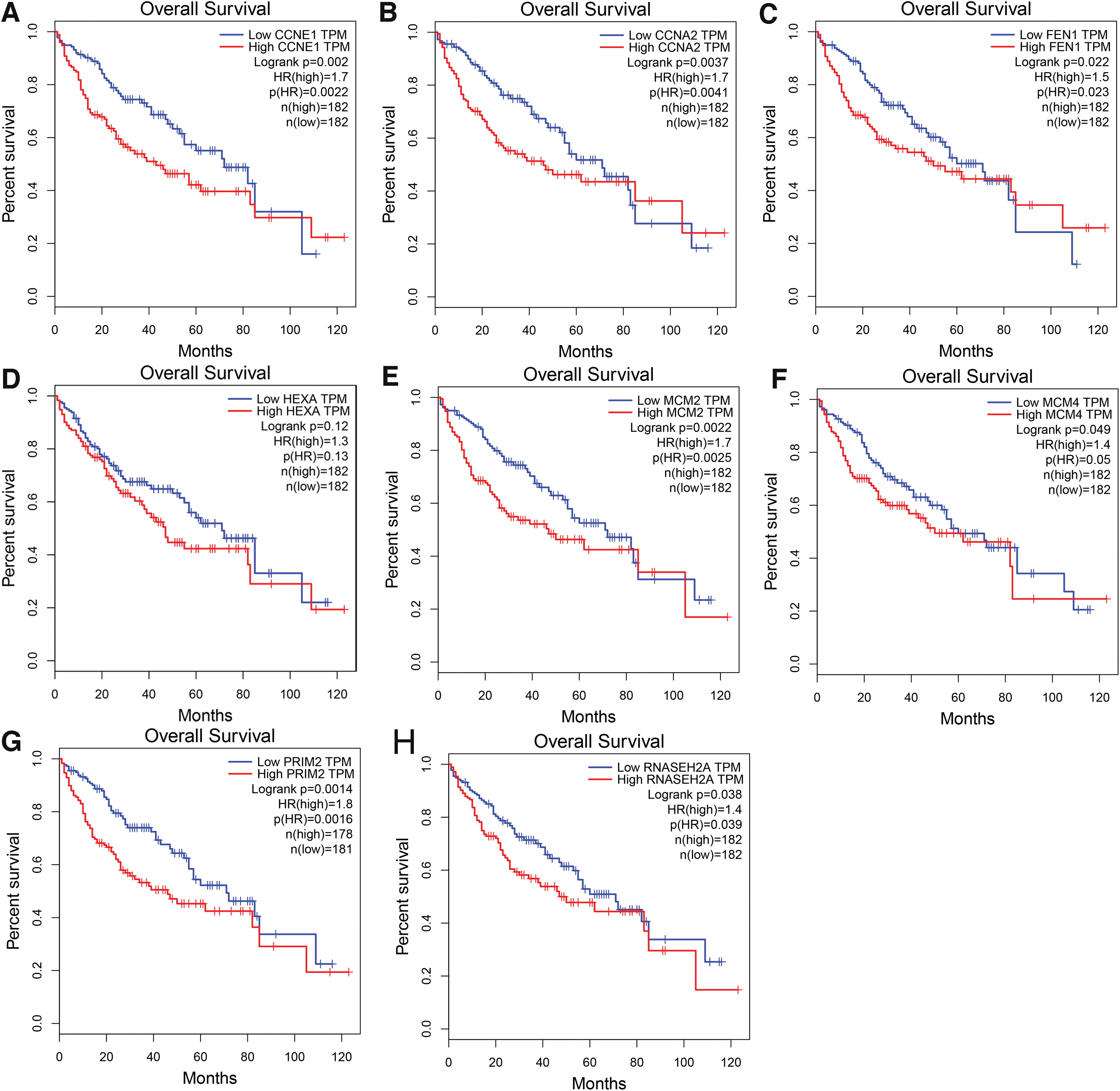
Prognostic values of hub genes. Higher expression levels of
Protein expression value of E2F1 in HCC
As given in Table 6, E2F1 protein expression was significantly higher in HCC in comparison with non-HCC, which was independent of various clinicopathological parameters. IHC staining results also qualitatively validated the upregulation of E2F1 in HCC patients (Fig. 11).
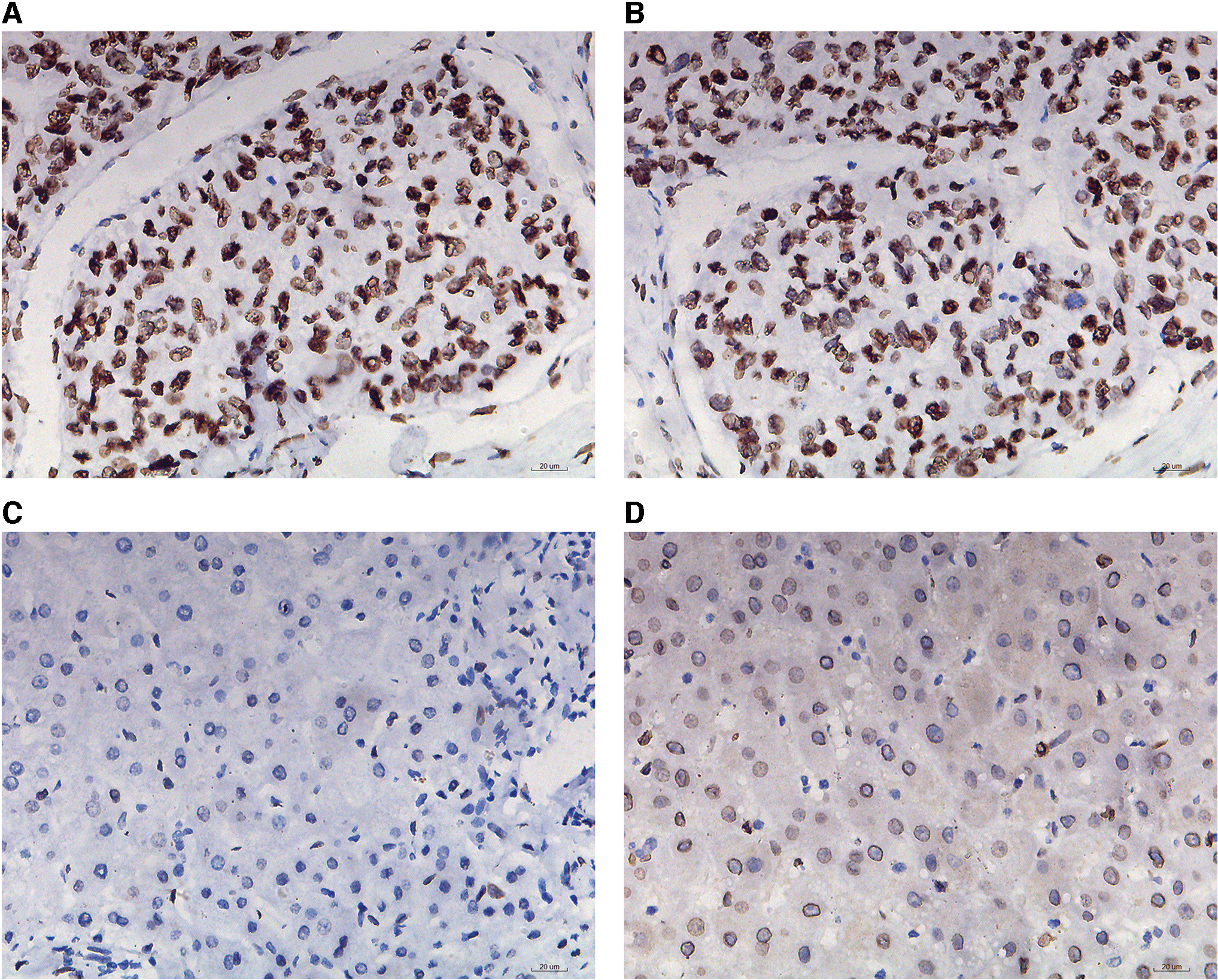
Immunohistochemistry staining presented higher protein expression of E2F1 in HCC tissues in comparison with normal liver tissues.
The Protein Expression Levels of E2F1 in Hepatocellular Carcinoma
Discussion
Despite the progress made in cancer treatments, the clinical management of advanced HCC patients remains tough. 30,31 Accumulated studies demonstrate that hepatitis viruses, cirrhosis, aflatoxin, alcohol consumption, diabetes, and drinking water pollution are risk factors for HCC. 32 –38 Nevertheless, the underlying causes of HCC onset and progression are unknown thus far. Therefore, our study is aimed at discovering novel biomarker for HCC early diagnosis and prognostic prediction, exploring its potential biological functions and concealed molecular mechanisms, and providing new direction for HCC disease coping.
Recently, transcriptional regulation has become a hotspot for cancer study. 39 –41 Transcriptional factor E2F1, which exerts profound biological functions by regulating cell cycle, plays different or even opposite roles in various types of cancer. E2F1 serves as a tumor oncogene in esophageal squamous cell carcinoma, whereas it exhibits tumor inhibitor activity in colorectal adenocarcinoma. 42,43 With regard to HCC, the role of E2F1 remains controversial for its double activities, either proapoptotic or antiapoptotic, which extremely limits the therapeutic application of E2F1 in HCC. Large bodies of studies report that E2F1 is upregulated in HCC patients, which is owing to the regulatory loops of E2F1, c-MYC, and miR-17-5p. 44,45 However, the clinical significance of E2F1 in HCC diagnosis and prognosis has not been extensively studied.
One of the advantages of this study is that we compressively analyze the E2F1 mRNA expression status and evaluated its clinical significance in HCC patients. We confirmed the elevated expression of E2F1 in HCC by integrating 76 studies covering 3856 HCC patients and 3309 non-HCC controls. In-depth subgroup analyses showed that the upregulated expression levels of E2F1 in HCC patients were independent of race, age, gender, specimen types of control groups, BCLC class, tumor recurrent status, and TNM tumor stage. The relevance between E2F1 and clinicopathological parameters of HCC patients were assessed for the first time, in which higher E2F1 was relevant to Asian race, higher tumor classification, neoplasm histologic grade, eastern cancer oncology group, and plasma AFP levels. Furthermore, high E2F1 correlated with poor survival condition and pooled HR signified E2F1 as a risk factor for HCC, thus indicating that E2F1 may well serve as a promising prognostic marker for HCC patients and effectively optimize clinical decision. The poor prognosis of E2F1 upregulation found in this study is consistent with the roles of E2F1 in promoting HCC progression from previous studies. 46,47
Moreover, we shed light on the biological functions and mechanisms of DEGs, E2F1, and its CEGs, as well as putative targets of E2F1 in HCC. Our results point to the key role of E2F1 in cell cycle regulation, especially G1–S period transition. Fortunately, this finding can be supported by scientific literature evidences. In an in vitro study, a novel polyplex combining α, β-poly-(N-2-hydroxyethyl)-d, L-aspartamide, diethylene triamine, polyethylene glycol, and galactose/siE2F1 was used to treat HCC cells. It was observed that the expressions of E2F1 and E2F1-related genes were significantly downregulated, all of which were aggregated in the promotion of G1/S transition, thus resulting in an increase in G0/G1 phase accumulation and an inhibition in cell proliferation. 48 As far as we are concerned, E2F1 transcriptional activity is suppressed by hypophosphorylated RB, which can bind to E2F1 during G1 phase in cell cycle. Provided that cyclin-CDK complex expedites the hyperphosphorylation of RB, E2F1 can isolate from RB and activates the transcription process by serving as essential regulatory points of G1 to S period transition. Therefore, deregulated E2F1 promotes the pathogenesis of cancers. According to Yi et al., overexpressed E2F8 activates E2F1/cyclin D1 signaling pathway and lead to HCC formation. 49 In addition, E2F1 induces proliferation of HCC cells by mediating IQGAP3 transactivation. 28 Furthermore, E2F1 has been found to correlate to several risk factors of HCC. 50 For example, AFB1 induced E2F1 overexpression and facilitated HCC cell growth. 51 E2F1 activation had roles in ATAD5 upregulation, which facilitated hepatitis B virus replication and protest tumor cells against treatment agents. 52 However, derivate isocorydine downregulates E2F1 and hampers HCC cell invasion. 53 More importantly, E2F1 drive cancer deterioration by recruiting pontin and reptin, which caused changes in the chromatin conformation of E2F1 target genes thus enriching transcriptional activity. 54 Our study found that the overlapping genes of upregulated DEGs, E2F1 positively related CEGs, and putative targets of E2F1 dominantly participated in various cancer diseases, which might confirm the roles that E2F1 plays in cancer.
The prominent highlight of this study is that CCNE1 and CCNA2 are identified as novel targets of E2F1 in HCC. Results of ChIP-seq peak intensities confirmed CCNE1 as an essential target gene of E2F1. In addition, CCNA2 promoter sequence segment (i.e., GGGGCGGGAGA) is highly matched to motif of transcriptional factor E2F1. To the best of our knowledge, CCNE1 and CCNA2 play pivotal roles in the pathway of cell cycle and their alterations has been utilized to define a homogenous form of aggressive HCC. 55 Lately, adeno-associated virus DNA insertions were identified in both CCNE1 and CCNA2, and those biological behaviors upregulated oncogenes through multiple mechanisms. 56 Cyclin E1 (CCNE1) is involved in HCC initiation and suppression of CCNE1 expression has been recognized as a promising therapeutic strategy. 57,58 Previous studies reported the association between E2F1 and CCNE1 in terms of expression level. 57 Furthermore, an in vitro and clinical study of Aurélie et al. evidenced that RIP140 attenuated transactivation of E2F1, which targeted at gene CCNE1 in breast cancer. 59 However, whether CCNE1 serve as a target gene of E2F1 in HCC has not been studied before. Our finding supports that CCNE1 might be also an essential target gene of E2F1 in HCC. E2F1 is most likely to accurately regulate G1–S transition in the process of cell cycle by targeting CCNE1, and its deregulation promotes dysplastic and invasive activity of tumor cells, thus causing carcinogenesis and deterioration of HCC.
On the contrary, deregulation of CCNA2 has been reported to participate in HCC development. 60 The study of Chiun et al. indicated that higher expression signatures of CCNA2 along with other cell cycle regulators correlated with poor survival condition HCC subclass, 61 which was in agreement with our survival analysis results. According to Tomoaki et al., CCNA2 might be a downstream target of B-Myb, and the latter is further regulated by E2F1. 17 Therefore, combined with our results, it is reasonable to suppose that CCNA2 might be also a pivotal target of transcriptional factor E2F1 in HCC. The biological and functional links between E2F1 and CCNE1, as well as CCNA2, are of paramount importance in S phase entry of cell cycle and HCC initiation or even progression. More importantly, CCNE1 and CCNA2 might serve as marvelous therapeutic targets for advanced HCC patients who establish resistance to sorafenib. 61
The paradox of E2F1's dual-directional role in HCC might be partially explained by its target gene CCNA2. Traditionally, E2F1 is thought to be involved in regulating cell apoptosis, especially when DNA is impaired, and this apoptotic process can be modulated by transcriptional cofactors. 62 –65 On the contrary, another voice has it that E2F1 induces antapoptosis by hampering c-Myc-mediated cell apoptosis. 22 Quantities of studies have explored the complicated reasons for opposite roles that E2F1 had in HCC. According to Roh et al., both Skp2 deletion and p27T187A mutation were capable of converting the biological effects of E2F1 in HCC, from proliferation to apoptosis. 66 Manickavinayaham et al. demonstrated that E2F1 expedited replication-stress-induced DNA damage and caused cells to start a p53-independent apoptotic process. 67 In addition, E2F1 might counteract c-Myc-induced apoptosis by activating several signaling pathways and downregulation of E2F1, caused by WIF1-Fc and sFRP1-Fc, promoted tumor cell apoptosis. 23 Based on our results, we raised the hypothesis that in the advanced stage of HCC, E2F1 might either directly modulate the antiapoptosis function of CCNA2 or initially upregulate B-Myb, which subsequently activates CCNA2's antiapoptotic activity. 17,61 However, further studies are needed in the future to confirm our conjecture.
When compared with the previous studies concerning E2F1 and HCC, our study exhibits unique advantages. We integrated datasets mined from several public databases and calculated the SMD of E2F1 expression level in HCC and non-HCC groups to comprehensively understand the elevated E2F1 expression in HCC patients. In addition, we underscored the moderate discriminatory capacity of E2F1 in differentiating HCC patients from non-HCC controls. Furthermore, in-depth subgroup analyses indicated the universally increased levels of E2F1 in HCC patients from various subgroups and survival analysis together with pooled HR recognized E2F1 as a valuable prognostic biomarker of poor outcomes in HCC patients. Moreover, CCNE1 and CCNA2 were identified as predominant targets of E2F1 in HCC onset and deterioration, which provides a promising therapeutic strategy for advanced HCC patients.
Nevertheless, several limitations that appear in our study should not be ignored. First, large scales of clinical practices are needed to validate the prognostic significance of E2F1 in HCC. Second, the complicated role that E2F1 plays in HCC, by targeting CCNE1 and CCNA2, should be validated in vivo and in vitro experiments in the future. Third, the associations between E2F1 and HCC require to be further studied and more evidences must be provided before it can be put into use in real life.
In conclusion, we confirmed the upregulation of E2F1 and explored the prognostic value of E2F1 in HCC patients. Two target genes (i.e., CCNE1 and CCNA2) of E2F1 were identified for their essential roles in regulating cell cycle and promote antiapoptotic activity in HCC patients.
Conclusions
We confirmed the upregulation of E2F1 and explored the prognostic value of E2F1 in HCC patients. Two putative targeted genes (CCNE1 and CCNA2) of E2F1 were identified for their potential roles in regulating cell cycle and promote antiapoptotic activity in HCC patients.
Footnotes
Authors' Contributions
Designing of the study, supervising all experiments, correcting the draft: H.-P.L., G.C., R.-Q.H., H.-Y.W., X.-G.Z., M.-H.R., L.-H.Y., S.-L.P., Y.-Y.P., and Y.-W.D.; performing experiment: W.-Y.Y.; performing gene chip, high-throughput sequencing datasets screening, statistical analysis, and drafting the article: J.-D.L., W.-Y.H., and Q.-Y.P. All authors have reviewed and approved the final version of the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
The authors thank the following funding agencies: Fund of National Natural Science Foundation of China (NSFC 81860717), Natural Science Foundation of Guangxi, China (2019GXNSFAA245074, 2019GXNSFAA245087), Fund of Natural Science Foundation of Guangxi, China (2017GXNSFAA198017), Promoting Project of Basic Capacity for Young and Middle-aged University Teachers in Guangxi (2018KY0123), Guangxi Zhuang Autonomous Region Health Committee Self-Financed Scientific Research Project (Z20190529), Guangxi Medical University Training Program for Distinguished Young Scholars, Medical Excellence Award Funded by the Creative Research Development Grant from the First Affiliated Hospital of Guangxi Medical University, Guangxi Degree and Postgraduate Education Reform and Development Research Projects, China (JGY2019050), Guangxi Higher Education Undergraduate Teaching Reform Project (2020JGA146), Guangxi Medical University Education and Teaching Reform Project (2019XJGZ04), Guangxi Medical University 2020 Undergraduate Innovation and Entrepreneurship Training Program (202010598002), The future academic star of Guangxi Medical University (WLXSZX20088). The authors also appreciated people who contributed to public databases, packages, and online tools.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.