
Abstract
Background:
Our research is designed to explore the role of 5-FU and Pulsatilla decoction (PD) through modulation of Immunogenic cell death (ICD) for the co-treatment of Colorectal cancer (CRC).
Materials and Methods:
Cell viability was evaluated by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazol-3-ium bromide (MTT) assays. Cell apoptosis was assessed using flow cytometry. Phosphorylation of STAT3 and expression of Mcl-1 and Bcl-xl were measured by Western blot assays. The levels of ATP and HMGB1 in the supernatants of the culture medium were analyzed by ATP assays and the HMGB1 enzyme linked immunosorbent assay kit. The cell surface levels of CRT were measured by immunofluorescence assays. The tumor growth was analyzed in mice.
Results:
PD increased 5-FU-induced ICD in CRC cells, as demonstrated by the extracellular levels of adenosine triphosphate (ATP) and high-mobility group box 1 (HMGB1), and the surface levels of calreticulin (CRT). Our mechanism study showed that PD promoted 5-FU-induced ICD by inactivating signal transducer and activator of transcription 3 (STAT3). Furthermore, the co-treatment of 5-FU and PD further promoted 5-FU-induced CRT expression and T cell infiltration in vivo. Tumorigenicity analysis revealed that 5-FU combined with PD notably reduced tumor growth.
Conclusion:
This study indicated that PD enhances 5-FU-induced ICD and anti-tumor effect in CRC by inactivating STAT3. The combined application of 5-FU with PD may improve the anti-tumor activity of 5-FU in CRC.
Introduction
Colorectal cancer (CRC) is the third most prevalent malignancy globally and serves as a primary cause of tumor-associated deaths in both women and men. 1 The number of recently diagnosed CRC patients increased from an estimated 1.4 million in 2012 to 1.8 million in 2018, and deaths due to cancer from 0.7 million to ∼0.9 million, respectively, currently accounting for ∼10% of all cancer occurrences and deaths globally with the growing trend of young individuals. 2,3 Although prevalence of CRC has unwaveringly decreased because of partially improved diagnostic and therapeutic strategies, and reduced exposure to risk factors, the 5-year survival rate of 64.9% was not acceptable, emphasizing the need for innovative detection and treatment applications. 4,5
Immunogenic cell death (ICD) is characterized by the chronic presentation in the tumor microenvironment (TME) with damage-associated molecular patterns (DAMPs), stimulating the antitumor immune system. 6 The potential factors that trigger an antitumor immune response rely on the regulation of DAMPs, such as high-mobility group box 1 (HMGB1), adenosine triphosphate (ATP), and calreticulin (CRT), which are essential for the modulation of ICD. 7 In tumor treatment, this is beneficial as cancer cells undergoing ICD stimulate an antitumor immune response specific to that cancer. 8 Therefore, developing therapies that provoke ICD and promote the practical cooperation of the patient's adaptive immune system, provide the opportunity to improve the clinical outcomes of cancer therapeutics. 9 –11 ICD plays a critical role in the immunotherapy for CRC. 12,13 Accordingly, ICD is a potential target for the CRC therapy and the ICD-targeted treatment may improve the outcomes of current therapeutic strategy as the promising candidate. However, the therapeutic strategy targeted to ICD in CRC is extremely limited until now.
Multiple chemotherapeutic agents, such as oxaliplatin and 5-fluorouracil (5-FU), have been identified as inducers of ICD. 14,15 5-FU serves as a pyrimidine analog, widely employed in cancer therapy and particularly efficient in the treatment of head and neck cancers, breast, stomach, and CRC. 16 –18 As an antitumor agent, 5-FU presents multiple drawbacks, such as short plasma half-life and high toxicity due to its water solubility. 19 Correspondingly, it has been recognized that the combination therapy can enhance chemotherapy-induced ICD, improving the chemosensitivity of cancer and relieving its toxic and side effects. 15,20,21
Pulsatilla decoction (PD) is a classical prescription of traditional Chinese medicine (TCM) that is initially described nearly 1800 years ago by Zhang Zhongjing in “Shang Han Lun.” 22 PD comprises of four herbs, Bai Tou Weng (Pulsatillae radix (Bge.) Regel), Huangbai (Phellodendron chinense Schneid), Huanglian (Coptis chinensis Franch), and Qinpi (Fraxinus rhynchophylla Hance). 22 PD has presented valuable biomedical activity for multiple diseases, such as inflammation and colitis. 22 Meanwhile, several investigations have identified the role of Pulsatilla chinensis saponins in inducing cell apoptosis at physiological conditions. 23,24 It has been well identified that immune activation caused by inflammation is well known to increase cancer risk and tumor necrosis factor (TNF) that is closely correlated with ICD. 25,26 PD is able to attenuate colitis by inhibiting inflammation and epithelial barrier disruption in mice. 22 PD inhibits interleukin-1 (IL-1) α, TNF-α, ET-1, and NO secretion in intestinal microvascular endothelial cells of LPS-induced rat. 27 These studies identify the significant role of PD in modulating inflammation and implies the potential correlation of PD with ICD. However, the role of PD in cancer progression and 5-FU-induced ICD remains unclear.
In this study, they aimed to explore the role of 5-FU combined with PD in the progression and ICD of CRC. They identified that PD enhanced 5-FU-induced ICD and antitumor effect on CRC by inactivating signal transducer and activator of transcription 3 (STAT3).
Materials and Methods
Cell culture and treatment
The human CRC, CT26, and HCT-116 cells were purchased from the American Type Tissue Culture Collection. The cells were cultured in the medium of Dulbecco's modified Eagle's medium (Gibco, China) containing 10% fetal bovine serum (Gibco), 0.1 mg/mL streptomycin (Gibco), and 100 U/mL penicillin (Gibco) at a condition of 37°C with 5% CO2. PD was obtained from the Affiliated Hospital of Southwest Medical University. The PD was diluted to 1, 5, and 10 mg/mL to select the appropriate concentration of PD in the cells. The 5-FU was purchased from Sigma-Aldrich (St. Louis, MO). The 5-FU and PD were applied at the dose of 0.8 mM and 10 mg/mL in the cells, respectively. The pcDNA3.1-STAT3 overexpression vector was synthesized and obtained (GenScript, China). The transfection in the cells was performed by using Lipofectamine 3000 (Invitrogen, Waltham, MA) according to the manufacturer's instructions.
3-(4,5)-dimethylthiahiazo (-z-y1)-3,5-di-phenytetrazoliumromide assays
The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazol-3-ium bromide (MTT) assay was used to measure the cell viability of CT26 and HCT-116 cells after 48 h of treatment. Briefly, about 1 × 104 CT26 and HCT-116 cells were planted in 96 wells and treated as indicated for 24 h, respectively. After 48 h of treatment, the cells were added with a 10 μL MTT solution (5 mg/mL) (Sigma) and cultured for an extra 4 h. Discarded medium, and 150 μL/well dimethyl sulfoxide (Thermo) was used to treat the wells. An enzyme-linked immunosorbent assay (ELISA) browser was applied to analyze the absorbance at 570 nm (Bio-Tek EL 800).
Analysis of cell apoptosis
The apoptosis of CT26 and HCT-116 cells was analyzed by using the Annexin V-FITC Apoptosis Detection Kit (CST, Boston, MA) according to the manufacture's instruction after 24 h of treatment. Briefly, ∼2 × 105 CT26 and HCT-116 cells were plated on six-well dishes, respectively. The treated cells were washed with phosphate-buffered saline and resuspended with 100 μL of binding buffer. Then, 5 μL of Annexin V-FITC and 10 μL of propidium iodide were added into the suspensions and incubated at room temperature for 10 min in the dark. The apoptosis rate was analyzed using flow cytometry (FACSCalibur, BD).
Western blot analysis
Western blots were carried out as previously described with the following modifications. 23 Total proteins were extracted from the cells or tumor tissues of the mice with radioimmunoprecipitation assay buffer (CST). Protein concentrations were measured by using the BCA Protein Quantification Kit (Abbkine, California). The same concentration of protein was divided by sodium dodecyl sulfate/polyacrylamide gel electrophoresis (12% polyacrylamide gels), transferred to polyvinylidene difluoride membranes (Millipore) in the subsequent step. The membranes were blocked with 5% milk and incubated overnight at 4°C with the primary antibodies for tyrosine phosphorylation of STAT3 (Y705) (CST), STAT3 (CST), Mcl-1 (CST), Bcl-xl (CST), and β-actin (CST). β-actin served as the control. Then, the corresponding secondary antibodies (1:1000) (Abcam, UK) were used to incubate the membranes 1 h at room temperature, followed by the visualization using an Odyssey CLx Infrared Imaging System. The results of Western blot analysis were quantified using ImageJ software.
Analysis of ICD markers
The CT26 and HCT-116 cells were treated with PD, 5-FU, or cotreated with 5-FU and PD for 24 h, respectively. The levels of ATP and HMGB1 in the supernatants of the culture medium were analyzed by ATP assays (Beyotime Biotechnology, China) and the HMGB1 ELISA Kit (Chondrex), respectively. The cell surface levels of CRT were measured by immunofluorescence assays as previously described. 28 Briefly, the cells were solidified with 4% paraformaldehyde for 30 min and treated with Triton X-100 (0.2%) for 10 min. The slides were incubated with the anti-CRT antibody (Abcam, Cambridge, MA) overnight at 4°C, then incubated with secondary antibodies (Abcam, Cambridge, MA) for 1 h at 37°C. The nuclei were stained with DAPI. A Nikon microscope (Tokyo, Japan) was utilized to analyze the immunofluorescence.
Analysis of ICD and tumorigenicity in vivo
To investigate the effect of the combination of PD and 5-FU on ICD in vivo, the BALB/c mice (male, 4-week-old, n = 15) were randomly divided into three groups (n = 5). Mice were subcutaneously injected with CT26 cells, followed by the treatment with 5-FU (25 mg/kg) or cotreatment of 5-FU (25 mg/kg) and PD (20 g/kg). The 5-FU administration was conducted by intravenous tail vein
Statistical analysis
Data are presented as mean ± standard deviations, and the statistical analysis was performed using SPSS software (version 18.0). The one-way analysis of variance was applied for comparing among multiple groups, and Tukey's post hoc test was used for post hoc analysis. p < 0.05 was considered as statistically significant.
Results
5-FU combined with PD inhibits the progression of CRC in vitro
To evaluate the cotreatment effect of PD and 5-FU on CRC cells, they first assessed the safety of PD in CT26 and HCT-116 cells. MTT assays revealed that PD failed to inhibit viability of CT26 and HCT-116 cells (Fig. 1A). Significantly, they found that 5-FU was able to reduce the cell proliferation of CT26 and HCT-116 cells, and the combination of PD with 5-FU (10 mg/mL) could enhance the reduction in the cells (Fig. 1B). Moreover, PD failed to promote apoptosis of CT26 and HCT-116 cells. The 5-FU increased cell apoptosis in the CT26 and HCT-116 cells, whereas PD and 5-FU cotreatment further promoted this phenotype in the system (Fig. 1C). Altogether, these data suggested that 5-FU combined with PD inhibits the progression of CRC in vitro.
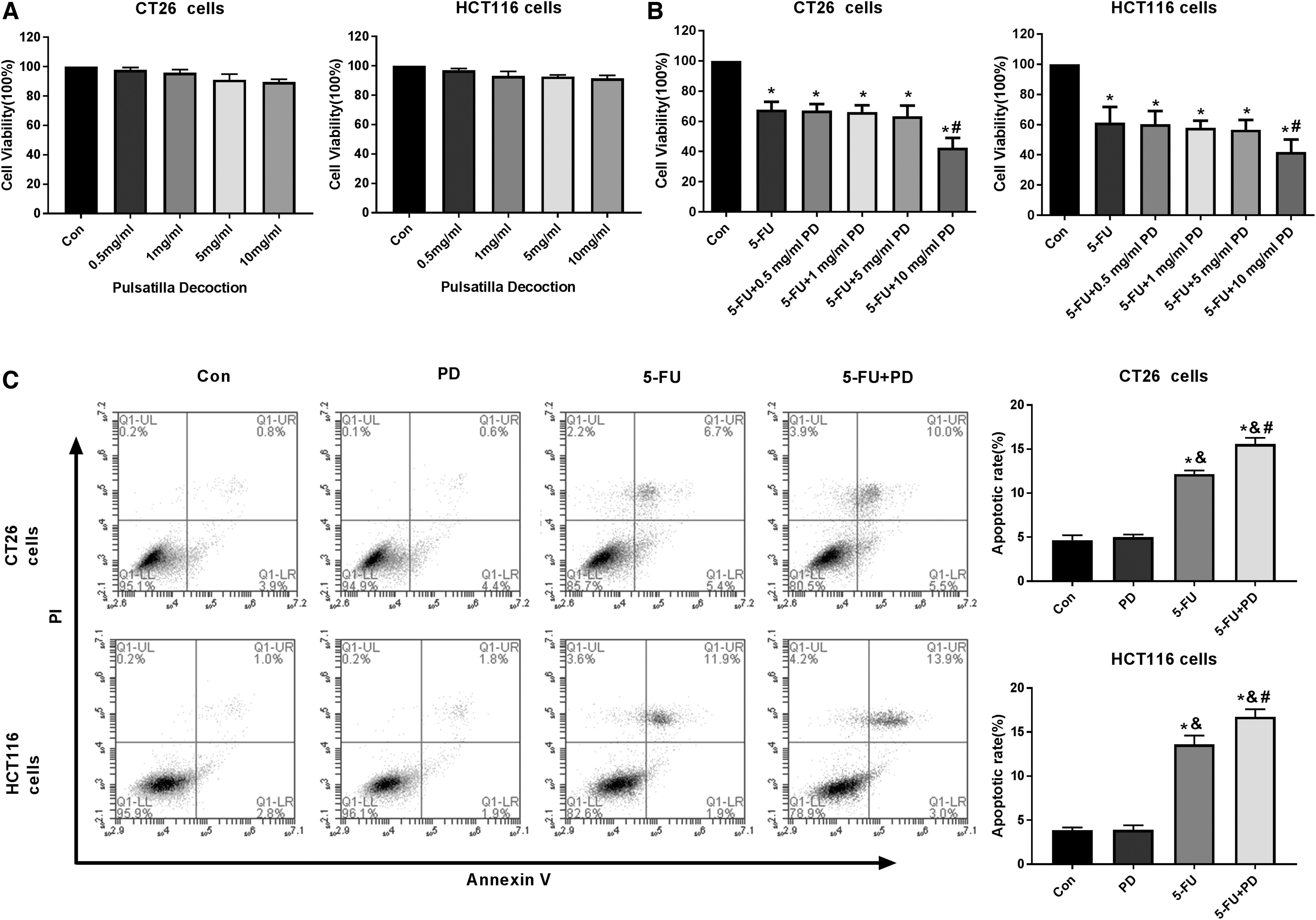
5-FU combined with PD inhibits the progression of CRC in vitro.
PD enhances 5-FU-induced ICD in CRC cells
ICD-related tumor-specific immune responses can affect the efficiency of antitumor therapy of conventional cytotoxic drugs. 29 It has been recognized that the chemotherapeutic agent can serve as the inducer of ICD in the immunotherapy of cancer. 14 Accordingly, the authors assessed the effect of the combination of 5-FU with PD on ICD in the CRC cells. Significantly, 5-FU treatment in the CT26 and HCT-116 cells increased the extracellular levels of ATP, and the cotreatment of 5-FU and PD reinforced the ATP levels in the system. However, the ATP levels did not change in the PD-treated CT26 and HCT-116 cells (Fig. 2A). Similarly, the secretion of HMGB1 was not changed by PD, whereas it was upregulated by 5-FU, and the combination of 5-FU and PD increased this upregulation in CT26 and HCT-116 cells (Fig. 2B). Furthermore, immunofluorescent assays revealed that cell surface levels of CRT were not changed by PD, but enhanced by 5-FU in the cells, whereas the combination of 5-FU and PD further increased the phenotype (Fig. 2C), indicating that PD promotes 5-FU-induced ICD in CRC cells.
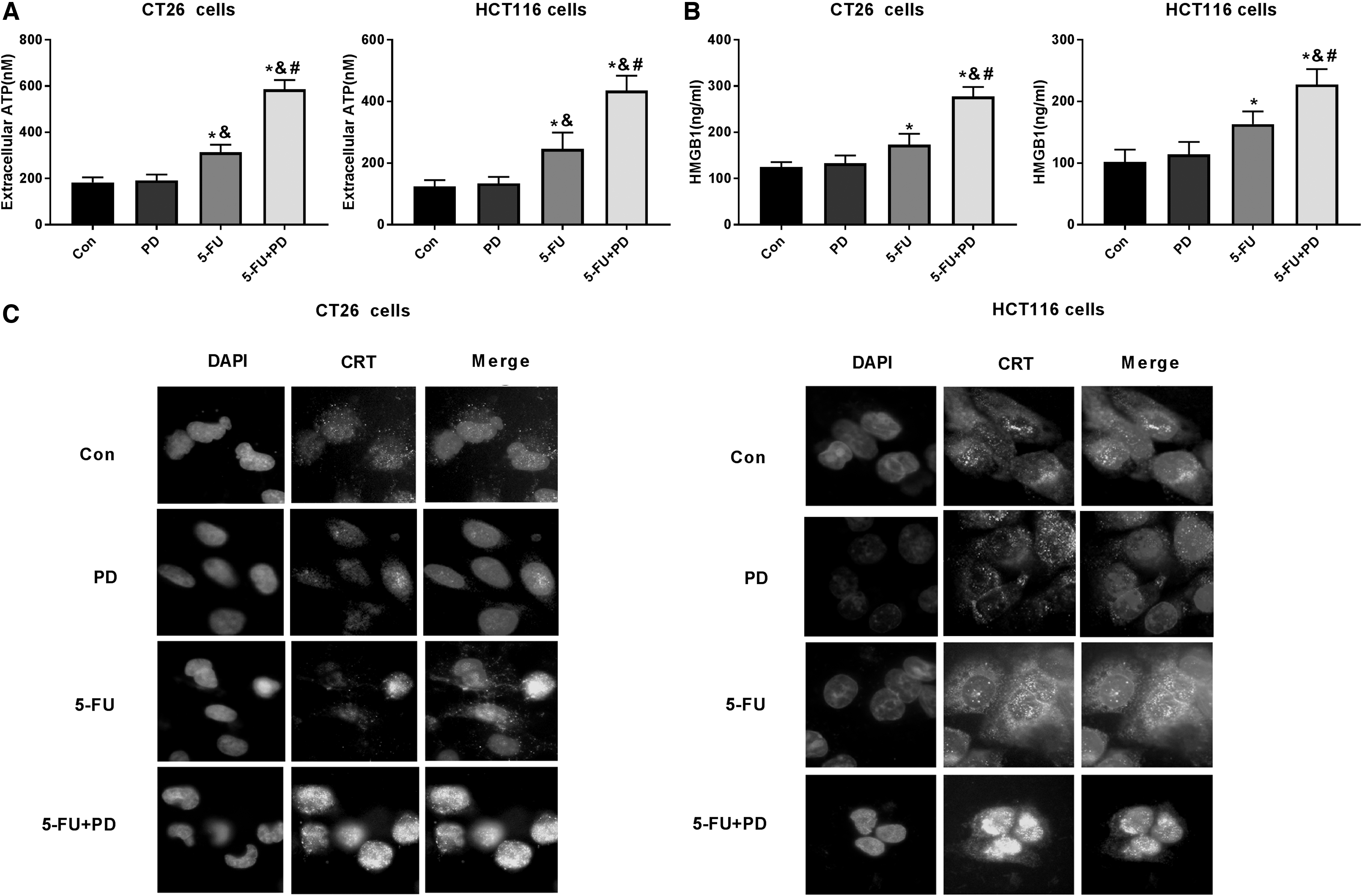
PD enhances 5-FU-induced ICD in CRC cells.
The combination of 5-FU and PD inhibits the STAT3 activation
Next, the authors further explored the mechanism by which PD enhanced 5-FU-induced ICD. Inhibition of STAT3 has been reported to promote ICD in cancer cells. 30 Meanwhile, STAT3 signaling is involved in 5-FU-induced antitumor function. 31 Hence, they evaluated whether STAT3 signaling participates in the combined effect of PD and 5-FU on ICD. Western blot analysis demonstrated that the tyrosine phosphorylation of STAT3 (Y705) was reduced by PD and 5-FU, and the cotreatment of PD and 5-FU further decreased the phosphorylation in the CT26 and HCT-116 cells (Fig. 3A). As expected, the expression levels of downstream targets of STAT3, including Mcl-1 and Bcl-xl, were downregulated by the PD and 5-FU cotreatment in the cells (Fig. 3B), suggesting that the combination of PD and 5-FU inhibits the activation of STAT3.

The combination of 5-FU and PD inhibits the STAT3 activation.
PD promotes 5-FU-induced ICD by inactivating STAT3
Then, the authors' data revealed that cotreatment of 5-FU and PD promoted the extracellular levels of ATP and HMGB1 in CT26 and HCT-116 cells, while the overexpression of STAT3 blocked the phenotype in the system (Fig. 4A, B). Similarly, STAT3 overexpression reversed the surface expression of CRT enhanced by the combination of PD and 5-FU (Fig. 4C). Altogether these data suggest that PD enhances 5-FU-induced ICD by inactivating STAT3.

PD promotes 5-FU-induced ICD by inactivating STAT3.
5-FU combined with PD enhances ICD and inhibits the progression of CRC in vivo
Next, they further investigated the effect of the combination of PD and 5-FU on ICD in vivo. To this end, the BALB/c mice were injected with CT26 cells, followed by the treatment of 5-FU, PD, or cotreatment of 5-FU and PD. Tumorigenicity analysis showed that there was no statistical difference in tumor volume and weight between the PD and control groups. PD failed to inhibit tumor growth of CT26 cells (Supplementary Fig. S1A–C). 5-FU combined with PD notably reduced tumor volume (Fig. 5A), tumor size (Fig. 5B), and tumor weight (Fig. 5C) in the mice, indicating that the combination of 5-FU and PD inhibits the progression of CRC in vivo. Besides, immunohistochemical staining analysis revealed that the combination of 5-FU and PD remarkably promoted the infiltration of CD8+ T cells into tumor tissues (Fig. 5D), suggesting that PD is able to enhance 5-FU-induced ICD in vivo.

5-FU combined with PD enhances ICD and inhibits the progression of CRC in vivo.
Discussion
CRC is the third most prevalent cancer and affects nearly 700,000 individuals per year, making it one of the most lethal malignancies globally. 32,33 Several previous studies have reported several investigations about whether the combination treatment improves the effect of 5-FU on CRC progression. It has been reported that dihydroartemisinin promotes the antitumor function of 5-FU in the resistant CRC cells. 34 Esculetin or gefitinib improves the inhibitory impact of 5-FU on the progression of CRC. 35,36 However, the combination treatment of 5-FU with TCM is still limited. Moreover, PD has presented biomedical activity in multiple diseases. For example, PD suppresses Candida albicans adhesion and proliferation of vulvovaginal candidiasis by the Dectin-1 signaling in a mouse model. 37 Modified PD inhibits oxazolone-provoked colitis by suppressing epithelial barrier disruption and inflammation in mice. 22 PD is able to attenuate the dampness/heat diarrhea in rat model by controlling glycerin phospholipid metabolism-based lipidomic approach. 38 PD represses secretion of IL-1 α, TNF-α, endothelin-1, and nitric oxide in intestinal microvascular endothelial cells of lipopolysaccharide-induced rat. 27 In this study, their data showed that 5-FU combined with PD inhibited the progression of CRC in vitro and in vivo. Their data demonstrated a new function of PD involving in the antitumor activity of 5-FU. This is the first report of PD in the modulation of CRC progression, providing the valuable evidence of the role of PD in cancer development.
ICD assists in blocking the immunosuppressive TME, maintaining the natural components, and priming T cell-regulated adaptive immunity, and therefore ICD is the critical target for the CRC combinational therapy. 25 It has been reported that the epithelial growth factor receptor-related antibody cetuximab coupled with chemotherapy, promotes ICD in CRC. 39 Photothermal/chemotherapy-combined nanoparticles with TME response improves the induction of ICD for efficient treatment of CRC. 40 Trifluridine/Tipiracil, combined with oxaliplatin, increases PD-1 blockade by provoking ICD in CRC. 41 Nanosecond integrated electric fields stimulate endoplasmic reticulum stress and ICD in CRC murine models. 42 Their data demonstrated that PD enhanced 5-FU-induced ICD of CRC in vitro and in vivo. Their findings presented valuable information of the combinational effect of 5-FU and PD on ICD, providing a new evidence of the role of combination therapy in targeted ICD in cancer.
As essential cellular signaling in cancer progression, STAT3 signaling is involved in the modulation of ICD. Inhibition of STAT3 promotes Newcastle disease virus-induced ICD in prostate cancer cells. 30 Static increases ICD induced by chemotherapy by targeting STAT3 in cancer cells. 43 Cytostatic hydroxycoumarin OT52 stimulates ER/Golgi stress and targets STAT3-promoted synergy and noncanonical cell death with BH3 mimetics of lung cancer. 44 Inhibition of STAT3 improves immunogenic chemotherapy. 45 The inhibition of STAT3 is involved in Oxaliplatin-induced disturbance of glioma cell apoptosis. 46 The BET bromodomain inhibitor i-BET151 reduces ovarian cancer metastasis and increases antitumor immunity by modulating STAT3 signaling. 47 In the present study, the authors found that STAT3 was involved in the combinational effect of 5-FU and PD on the ICD. It provides new evidence of the crucial role of STAT3 in the modulation of ICD.
Conclusions
In conclusions, they discovered that PD enhanced 5-FU-induced antitumor effect and ICD in CRC by inactivating STAT3. Their findings provide new insights into the mechanism by which the cotreatment of PD and 5-FU promotes ICD and inhibits the progression of CRC. The combined application of 5-FU with PD may improve the antitumor activity of 5-FU in CRC.
Availability of Data and Material
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Footnotes
Acknowledgment
All coauthors have reviewed and approved of the article before submission.
Authors' Contributions
X.Y. and W.C. performed the experiments. Y.J. provided technical support, analyzed the data, and edited the article.
Disclosure Statement
There are no existing financial conflicts.
Funding Information
This work was supported by the Special Scientific Research Project of Health Young Medical Science and Technology talents in Xinjiang Uygur Autonomous Region (WJWY-201939).
Supplementary Material
Supplementary Data
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.