
Abstract
Background:
Circular RNAs (circRNAs) have important roles in human malignancies, including breast cancer (BC). In this study, we explored the function of circRNA ribonuclease P RNA component H1 (circ_RPPH1) in BC development and clarify the mechanistic pathway.
Materials and Methods:
Expression of circ_RPPH1, microRNA-542-3p (miR-542-3p), and Rho GTPase-activating protein 1 (ARHGAP1) in BC tissues and cells was determined by quantitative real-time polymerase chain reaction or Western blot assay. The stability of circ_RPPH1 was confirmed by RNase R and actinomycin D treatment. Cell viability and colony formation ability were measured by methyl thiazolyl tetrazolium (MTT) assay and colony formation assay, respectively. Western blot analysis was also used to detect proliferation biomarker (Ki67) and epithelial–mesenchymal transition (EMT) biomarkers (E-cadherin, N-cadherin, and vimentin). Flow cytometry and Transwell assays were performed to monitor cell apoptosis, migration, and invasion. The binding potency between miR-542-3p and circ_RPPH1 or ARHGAP1 was validated by dual-luciferase reporter assay. Functional role of circ_RPPH1 in vivo was investigated by xenograft tumor reporter assay.
Results:
Upregulation of circ_RPPH1 and ARHGAP1, and downregulation of miR-542-3p were detected in BC tissues and cells. circ_RPPH1 knockdown or miR-542-3p introduction inhibited BC cell proliferation and metastasis, while promoted apoptosis in vitro. circ_RPPH1 sponged miR-542-3p to upregulate ARHGAP1 expression, thereby affecting BC progression. Moreover, depletion of circ_RPPH1 suppressed tumor growth in vivo.
Conclusions:
circ_RPPH1 contributed to BC tumorigenesis by sponging miR-542-3p and upregulating ARHGAP1, affording a novel mechanistic pathway in BC development.
Introduction
Breast cancer (BC) possesses the second highest incidence all over the world, and is an intractable and challenging solid tumor. 1 The latest cancer statistics showed that BC ranked as the most prevalent and the second highest deadly cancer among female patients. 2 Clarifying the development of BC at molecular level might be beneficial for its therapy.
With special covalently closed loop structure, circular RNAs (circRNAs) emerged as significant biomarkers in cancer diagnosis and treatment. 3 Moreover, circRNAs were reported to have close association with multiple cellular behaviors of BC cells, and could serve as biomarkers in BC progression. 4 For example, highly expressed circRNA hsa_circ_001783 in BC made it to be a new prognosis marker; and silenced hsa_circ_001783 inhibited BC cell proliferation and invasion. 5 Gao et al. highlighted the prognostic predictor role of circ_0006528 in BC, and this circRNA could facilitate the aggressive malignant behaviors of BC cells. 6 circIRAK3 was manifested to contribute to BC metastasis by modulating miR-3607/FOXC1 signaling axis. 7 circRNA ribonuclease P RNA component H1 (circ_RPPH1), namely hsa_circ_0000515, was found to be dysregulated in primary BC tumors. 8 However, the explicit role of circ_RPPH1 in BC remains to be elucidated.
MicroRNAs (miRNAs) are also noncoding RNA molecules, ∼22 nucleotides long, which could post-transcriptionally silence target genes so as to exert regulatory function. 9 miRNA could function as oncogenic drive (such as miR-374a, miR-135b, and miR-331) or tumor suppressor (like miR-497, miR-26a, miR-30b, and miR-708) in BC, and serve as possible biomarkers for diagnosis and prognosis. 10,11 Besides, miRNA-21 and miRNA let-7 were recognized as prognostic markers of BC patients. 12 MicroRNA-542-3p (miR-542-3p), a potential target of circ_RPPH1 forecasted by circBank and Starbase, was previously reported to be a tumor-suppressive miRNA in BC. 13 In this study, we evaluated the role of miR-542-3p in circ_RPPH1-mediated effects on BC progression.
Rho GTPase-activating protein 1 (ARHGAP1), a member of the Rho GAP family, was involved in gene expression, cell growth, migration, and invasion. 14,15 Dong et al. highlighted the anti-tumor effects of ARHGAP1 knockdown on BC cells. 16 Given that ARHGAP1 was a target gene of miR-542-3p, which was predicted by Starbase, miRDB, and PITA, its interaction with circ_RPPH1 or miR-542-3p in BC remains unclear.
In this study, the dysregulation of circ_RPPH1 in BC tissues and cells was first uncovered. Then loss-of-function assays were performed to evaluate the function of circ_RPPH1 in BC cell proliferation, metastasis, and tumorigenesis. In addition, its downstream miR-542-3p was identified, their effects on cellular behaviors of BC were evaluated. Afterward, the target gene of miR-542-3p, and its role were also demonstrated.
Materials and Methods
Tissue samples and cells
Forty pairs of BC tissues and matched normal tissues were procured from 40 BC patients hospitalized at Cancer Institute and Cancer Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College. All patients did not undergo any therapeutic means before tissue collection. All experiments were executed after acquisition of written informed consents from all individuals as well as authorization from the Ethics committee of Cancer Institute and Cancer Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College.
Human normal mammary gland epithelial cell line MCF-10A (CRL-10317) and BC cell lines MCF-7 (HTB-22), MDA-MB-231 (HTB-26), MDA-MB-468 (HTB-132), SK-BR-3 (HTB-30) were all commercially obtained from American Tissue Culture Collection (Manassas, VA). Cell culture was implemented in Dulbecco's modified Eagle's medium (HyClone, Logan, UT) mixed with 10% fetal bovine serum (FBS; HyClone) and 1% penicillin/streptomycin (Transgen, Beijing, China) in a controllable incubator containing 5% CO2 and 95% air at 37°C.
Transient transfection
Small interfering RNA (siRNA) against circ_RPPH1 (si-circ_RPPH1), miR-542-3p mimic (miR-542-3p), miR-542-3p inhibitor (anti-miR-542-3p), and overexpression vector of ARHGAP1 (ARHGAP1), as well as their respective negative control (si-NC, miR-NC, anti-miR-NC, and pcDNA) were synthesized by GenePharma Co. Ltd (Shanghai, China). Then, oligonucleotides (40 mM) or plasmids (2 μg) were introduced into MCF-7 and MDA-MB-231 cells exploiting Lipofectamine 3000 (Thermo Fisher Scientific, Inc., Waltham, MA).
Quantitative real-time polymerase chain reaction
Extraction of RNA from clinical tissues and cells was conducted using TRIzol reagent (Life Technologies, Grand Island, NY) or mirVana miRNA Isolation kit (Life Technologies). Complementary DNA was generated using M-MLV Reverse Transcriptase (Invitrogen, Carlsbad, CA) or TaqMan MicroRNA Reverse Transcription Kit (Thermo Fisher Scientific, Inc.). Subsequently, qPCR was performed on Applied Biosystems 7500 thermocycler (Thermo Fisher Scientific, Inc.) utilizing SYBR Green mix (TaKaRa, Otsu, Japan) or TaqMan MicroRNA assays (Thermo Fisher Scientific, Inc.). Relative expression of circ_RPPH1, linear RPPH1, miR-542-3p, and ARHGAP1 was analyzed by 2−ΔΔCt cycle threshold method, and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH, for circ_RPPH1, RPPH1, and ARHGAP1) or U6 (for miR-542-3p). All qPCR primers were synthesized by Sangon (Shanghai, China): circ_RPPH1 sense, 5′-GGTCAGACTGGGCAGGAGAT-3′ and circ_RPPH1 antisense, 5′-GAGTGACAGGACGCACTCAG-3′; RPPH1 sense, 5′-TGGGCAGGAGATGCCGTGGA-3′ and RPPH1 antisense, 5′-CAAAGGAGGCATCCGCCGGG-3′; ARHGAP1 sense, 5′-CAGCGTGTGATGGAATCAG-3′ and ARHGAP1 antisense, 5′-TGGAGGAAGGAGCCATAAG-3′; GAPDH sense, 5′-GGTCGGAGTCAACGGATTTGGTTTGG-3′ and GAPDH antisense, 5′-AAACCATGTAGTTGAGGTCAAT-3′ miR-542-3p sense, 5′-GCGCGTGTGACAGATTGATAA-3′ and miR-542-3p antisense, 5′-GTGCAGGGTCCGAGGT-3′; U6 sense, 5′-CTCGCTTCGGCAGCACA-3′ and U6 antisense, 5′-AACGCTTCACGAATTTGCGT-3′.
RNase R and actinomycin D treatment
RNase R could degrade linear RNAs, rather than circRNAs; and circRNAs have longer half-life than linear RNAs. 17,18 Therefore, the stability of circ_RPPH1 was demonstrated by RNase R digestion and actinomycin D disposition.
For RNase R treatment, 5 μg RNA isolated from MCF-7 or MDA-MB-231 cells was digested by RNase R (3 U/μg; Epicentre Biotechnologies, Madison, WI) or not (Mock) at 37°C for 20 min. Then, treated RNA was subjected to quantitative real-time polymerase chain reaction (qRT-PCR) assay for circ_RPPH1 and RPPH1.
For actinomycin D disposition, culture medium containing MCF-7 or MDA-MB-231 cells was added with actinomycin D (2 μg/mL; Sigma-Aldrich, St. Louis, MO) or dimethyl sulfoxide (DMSO; Sigma-Aldrich) for 0, 8, 16, and 24 h. Then, cells were collected for RNA isolation and qRT-PCR assay.
Methyl thiazolyl tetrazolium assay
To monitor cell viability, transfected MCF-7 and MDA-MB-231 cells were subjected to methyl thiazolyl tetrazolium (MTT) assay. In short, 4 × 103 cells were plated on 96-well plates and cultured for 0, 1, 2, or 3 d. Then 10 μL MTT solution (5 mg/mL; Sigma-Aldrich) was dropped into each well. Four hours later, DMSO was added to replace original medium to dissolve crystals. The optical density value of each well at 490 nm was recorded by feat of a microplate reader (Bio-Rad, Hercules, CA).
Colony formation assay
This assay allowed colony formation ability of BC cells to be evaluated. After transfection, 4 × 102 cells were tiled into 24-well plates. After regular culture for 10 d, generated colonies (exceeding 50 cells) were treated by ethanol for fixation and then dyed with crystal violet solution (Sigma-Aldrich). Finally, colonies were counted and photographed.
Western blot
Radioimmunoprecipitation assay (RIPA) buffer (CWBIO, Beijing, China) was applied to isolate protein from clinical tissues and cells following the manufacturer's direction. After determination of quality and concentration, 40 μg protein samples were loaded on sodium dodecyl sulfate–polyacrylamide gel for electrophoresis, and then transferred to polyvinylidene difluoride membranes. All membranes were subjected for blockage in 5% fat-free milk, incubated with primary antibody and second antibody. Visualization of protein blot was realized utilizing a chemiluminescence kit (Thermo Fisher Scientific, Inc.), and gray values were analyzed by ImageJ software. All antibodies were as follows: anti-Ki67 (ab92742, 1:1000 dilution; Abcam, Cambridge, MA), anti-E-cadherin (ab40772, 1:1500 dilution; Abcam), anti-N-cadherin (ab18203, 1:2000 dilution; Abcam), anti-vimentin (ab193555, 1:1000 dilution; Abcam), anti-ARHGAP1 (ab224231, 1:1000 dilution; Abcam), anti-GAPDH (ab181602, 1:3000 dilution; Abcam), and goat anti-rabbit second antibody conjugated with horseradish peroxidase (ab205718, 1:5000 dilution; Abcam).
Flow cytometry
To examine cell apoptosis, Annexin V-fluorescein isothiocyanate (FITC) apoptosis detection kit (BD Biosciences, Franklin Lakes, NJ) was exploited. Based on the supplier's instructions, transfected BC cells were collected and stained with Annexin V-FITC and propidium iodide (PI) staining solution for 15 min away from light. Flow cytometer (BD Biosciences) was used to observe apoptotic BC cells. Apoptotic rate represented the percentage of cells with Annexin V-FITC positive and PI negative or positive.
Transwell assay
For invasion analysis, 5 × 104 transfected MCF-7 and MDA-MB-231 cells in 200 μL medium without serum were plated into the insert compartment of Transwell chamber (BD Biosciences) coated with Matrigel, whereas medium containing 10% serum was added into the lower compartment. After 48 h, cells invaded through the Transwell membrane were stained with crystal violet and counted under an optical microscope (magnification, × 100). The operation procedures for migration analysis began with 1 × 104 transfected cells, and the chamber was Matrigel free. Other steps were the same as those used for the invasion analysis.
Dual-luciferase reporter assay
Here, circBank and Starbase v3.0 were searched to seek the potential miRNAs interacting with circ_RPPH1. Starbase v3.0, miRDB and PITA were utilized to forecast the target genes of miR-542-3p. miR-542-3p and ARHGAP1 were identified as corresponding candidates. The following dual-luciferase reporter assay was carried out to validate above prediction. At first, wild-type luciferase reporter vectors of circ_RPPH1 (circ_RPPH1-wt) and ARHGAP1 (ARHGAP1-wt) were synthesized by inserting their respective fragment containing binding position with miR-542-3p into pGL3 luciferase promoter vector (Promega, Madison, WI). Subsequently, mutant-type vectors were constructed by cloning fragment of circ_RPPH1 and 3′ untranslated region (UTR) of ARHGAP1 harboring mutant binding sites, called circ_RPPH1-mut and ARHGAP1-mut. Generated luciferase reporter vector and miR-NC or miR-542-3p were cotransfected into MCF-7 and MDA-MB-231 cells. Forty-eight hours later, a dual-luciferase assay system kit was utilized for luciferase activity detection.
In vivo tumorigenicity assay
Experiments performed on animals were ratified by the Animal Care and Use Committee of Cancer Institute and Cancer Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College. Short hairpin RNA for circ_RPPH1 (sh-circ_RPPH1) and its negative control (sh-NC) were constructed by Genechem (Shanghai, China). Afterward, MCF-7 cells (2 × 106) stably transfected with sh-NC or sh-circ_RPPH1 were subcutaneously inoculated into the right flank of BALB/c nude mice (female, 4–6 weeks old; Shanghai SLAC Laboratory Animal Co., Ltd., Shanghai, China; n = 4). Ten days later, the size of formed tumors was measured every 5 d. Thirty days later, all mice were killed, and formed tumors were dissected for weighing and qRT-PCR or Western blot analysis.
Statistical analysis
All data derived from ≥3 repeat experiments were represented as mean ± standard deviation. Statistical analysis was implemented on SPSS 19.0 software with Student's t-test or one-way analysis of variance. Pearson correlation analysis was applied to determine the correlation among the levels of circ_RPPH1, miR-542-3p, and ARHGAP1 in 40 BC tissues. Value of p < 0.05 was recognized as statistically significant.
Results
circ_RPPH1 was upregulated in BC tissues and cells
To explore the relevance of circ_RPPH1 to BC progression, we first examined its expression level in BC tissues. qRT-PCR assay revealed that circ_RPPH1 was significantly upregulated in BC tissues relative to normal tissues (N = 40) (Fig. 1A). In addition, higher enrichment of circ_RPPH1 was also detected in BC cell lines (MCF-7, MDA-MB-231, MDA-MB-468, and SK-BR-3) compared with MCF-10A cells (Fig. 1B). After RNase R treatment, rather than RPPH1, circ_RPPH1 seemed to be resistant to RNase R digestion (Fig. 1C, D). The greater half-life of circ_RPPH1 in MCF-7 and MDA-MB-231 cells than RPPH1 further proved the stability of circ_RPPH1 (Fig. 1E, F). In sum, circ_RPPH1 was upregulated in BC.

circ_RPPH1 was upregulated in BC tissues and cells.
circ_RPPH1 acted an oncogenic role in BC cells in vitro
Given the dysregulation of circ_RPPH1 in BC, we then performed loss-of-function assays to investigate the effect of circ_RPPH1 on BC development in vitro. siRNA targeting circ_RPPH1 was designed to silence circ_RPPH1 expression in MCF-7 and MDA-MB-231 cells, the knockdown impact of si-circ_RPPH1 was confirmed by qRT-PCR assay, with si-NC as negative control (Fig. 2A). MTT assay showed that circ_RPPH1 knockdown efficiently reduced the cell viability of MCF-7 and MDA-MB-231 cells (Fig. 2B, C). Depletion of circ_RPPH1 also decreased the colony formation ability of BC cells (Fig. 2D) and protein level of Ki67 (a proliferation biomarker) in BC cells (Fig. 2E), which further suggested that circ_RPPH1 knockdown repressed BC cell proliferation. Flow cytometry manifested that circ_RPPH1 knockdown elevated the apoptotic rate of MCF-7 and MDA-MB-231 cells (Fig. 2F). As given in Figure 2G and H, less migrated and invaded cells appeared in si-circ_RPPH1 group compared with si-NC group. Besides, absence of circ_RPPH1 triggered the increase of E-cadherin protein and the reduction of N-cadherin and vimentin proteins in BC cells (Fig. 2I, J). These data collectively demonstrated that circ_RPPH1 knockdown retarded proliferation, mobility, and epithelial–mesenchymal transition (EMT) while induced apoptosis of BC cells in vitro.

circ_RPPH1 acted an oncogenic role in BC cells in vitro. MCF-7 and MDA-MB-231 cells were introduced with si-NC or si-circ_RPPH1.
circ_RPPH1 could sponge miR-542-3p in BC cells
In an attempt to explore how circ_RPPH1 affects BC progression, we searched the target miRNAs of circ_RPPH1 with the aid of circBank and Starbase, and identified miR-542-3p and miR-326 as candidates (Supplementary Fig. S1A). Moreover, miR-326 expression was changeless in BC with circ_RPPH1 knockdown (Supplementary Fig. S1B, C). Therefore, miR-542-3p was selected for following assays. The binding sites between circ_RPPH1 and miR-542-3p are given in Figure 3A. We then performed dual-luciferase reporter assay to verify the potential interaction. Data revealed that the luciferase intensity of circ_RPPH1-wt in MCF-7 and MDA-MB-231 cells reduced >50% in response to miR-542-3p, whereas the luciferase activity of circ_RPPH1-mut changed little (Fig. 3B, C). In addition, we found that depletion of circ_RPPH1 remarkably elevated miR-542-3p expression in BC cells (Fig. 3D). The relative expression of miR-542-3p in BC tissues and cells was assessed. The results showed that miR-542-3p was downregulated in BC cells (Fig. 3E) and tissues (Fig. 3F) in contrast to corresponding controls. Of note, circ_RPPH1 expression in BC tissues was negatively correlated with miR-542-3p level (r = −0.5424, p = 0.0003) (Fig. 3G). Taken together, circ_RPPH1 functioned as a sponge of miR-542-3p in BC cells.

circ_RPPH1 could sponge miR-542-3p in BC cells.
Silencing of miR-542-3p almost reversed circ_RPPH1 knockdown-mediated inhibitory effect on BC progression in vitro
In view of the fact that circ_RPPH1 targeted miR-542-3p in BC cells, rescue assays were implemented to study the impact of miR-542-3p on circ_RPPH1 knockdown-mediated BC cells. qRT-PCR assay suggested that anti-miR-542-3p undermined the upregulation of miR-542-3p caused by circ_RPPH1 knockdown (Fig. 4A). In addition, circ_RPPH1 knockdown mediated the decreased cell viability (Fig. 4B, C), colony formation ability (Fig. 4D) and Ki67 (Fig. 4E) expression were all weakened by miR-542-3p inhibitor. Flow cytometry uncovered that absence of circ_RPPH1-promoted cell apoptosis of BC cells, which was relieved by additional miR-542-3p inhibitor (Fig. 4F). As for cell metastasis, miR-542-3p inhibitor also attenuated circ_RPPH1 knockdown-induced reduced migration (Fig. 4G) and invasion (Fig. 4H) of BC cells, and the upregulation of E-cadherin and the downregulation of N-cadherin and vimentin (Fig. 4I, J). Therefore, circ_RPPH1 knockdown might inhibit BC development by activating miR-542-3p expression.

Silencing of miR-542-3p almost reversed circ_RPPH1 knockdown-mediated inhibitory effect on BC progression in vitro. MCF-7 and MDA-MB-231 cells were introduced with si-NC, si-circ_RPPH1, si-circ_RPPH1+anti-miR-NC, or si-circ_RPPH1+anti-miR-542-3p.
MiR-542-3p targeted ARHGAP1
Starbase, miRDB, and PITA were utilized to predict the target mRNAs of miR-542-3p, and UBR4, NAV1 and ARHGAP1 were discovered to have binding region with miR-542-3p (Supplementary Fig. S2A). In BC cells transfected with miR-542-3p, ARHGAP1 level significantly decreased among these three predicted genes (Supplementary Fig. 2B, C). Therefore, we chose ARHGAP1 as a target of miR-542-3p for further investigation. The binding sites between miR-542-3p and ARHGAP1 are given in Figure 5A. Dual-luciferase reporter assay manifested the target relationship between miR-542-3p and ARHGAP1 (Fig. 5B, C). Subsequently, the transfection efficiency of miR-542-3p and miR-542-3p inhibitor was confirmed by qRT-PCR assay (Fig. 5D). Remarkably, gain of miR-542-3p apparently repressed ARHGAP1 expression, whereas silencing of miR-542-3p induced reverse outcome, as demonstrated by qRT-PCR assay and Western blot analysis (Fig. 5E, F). Furthermore, ARHGAP1 expression in BC was examined. As given in Figure 5G and I–K, compared with relative control, the mRNA and protein levels of ARHGAP1 were significantly upregulated in BC tissues and cells. Of interest, an obvious negative correlation existed between the expression levels of miR-542-3p and ARHGAP1 mRNA in BC tissues (r = −0.5993, p < 0.0001) (Fig. 5H). Afterward, the effect of circ_RPPH1 on ARHGAP1 expression was determined. The downregulated mRNA and protein levels of ARHGAP1 caused by circ_RPPH1 knockdown were restored by miR-542-3p inhibitor (Fig. 5L, M). Taken together, miR-542-3p could target ARHGAP1 in BC cells.

MiR-542-3p targeted ARHGAP1.
MiR-542-3p suppressed BC progression by inhibiting ARHGAP1
To further clarify the role of miR-542-3p/ARHGAP1 axis in BC progression, a series of rescue experiments in MCF-7 and MDA-MB-231 cells were executed. qRT-PCR assay and Western blot analysis disclosed the restoration effect of overexpressed ARHGAP1 on the miR-542-3p-induced ARHGAP1 downregulation (Fig. 6A, B). Meanwhile, miR-542-3p-induced the reduction of cell viability (Fig. 6C, D), colony formation capacity (Fig. 6E), Ki67 expression (Fig. 6F), migration (Fig. 6H), and invasion (Fig. 6I), and the promotion of apoptosis (Fig. 6G) were relieved by ARHGAP1 overexpression. In addition, miR-542-3p also inhibited EMT process in BC cells, which was relieved by overexpression of ARHGAP1 (Fig. 6J, K). In conclusion, miR-542-3p played tumor-suppressor role in BC by downregulating ARHGAP1.

MiR-542-3p suppressed BC progression by inhibiting ARHGAP1. MCF-7 and MDA-MB-231 cells were transfected with miR-NC, miR-542-3p, miR-542-3p+pcDNA or miR-542-3p+ARHGAP1.
circ_RPPH1 facilitated BC tumor growth in vivo
The functional role of circ_RPPH1 in vivo was investigated by xenograft tumor assay. MCF-7 cells stably transfected with sh-NC or sh-circ_RPPH1 were injected into nude mice. From our data, the flank tumors in the sh-circ_RPPH1 group exhibited smaller size (Fig. 7A) and lighter weight (Fig. 7B) in contrast to sh-NC group. Furthermore, circ_RPPH1 (Fig. 7C) and ARHGAP1 (Fig. 7E, F) were downregulated, and miR-542-3p was upregulated (Fig. 7D) in tumors generated in sh-circ_RPPH1 group. These results demonstrated that circ_RPPH1 had significant protumoral activity in BC.
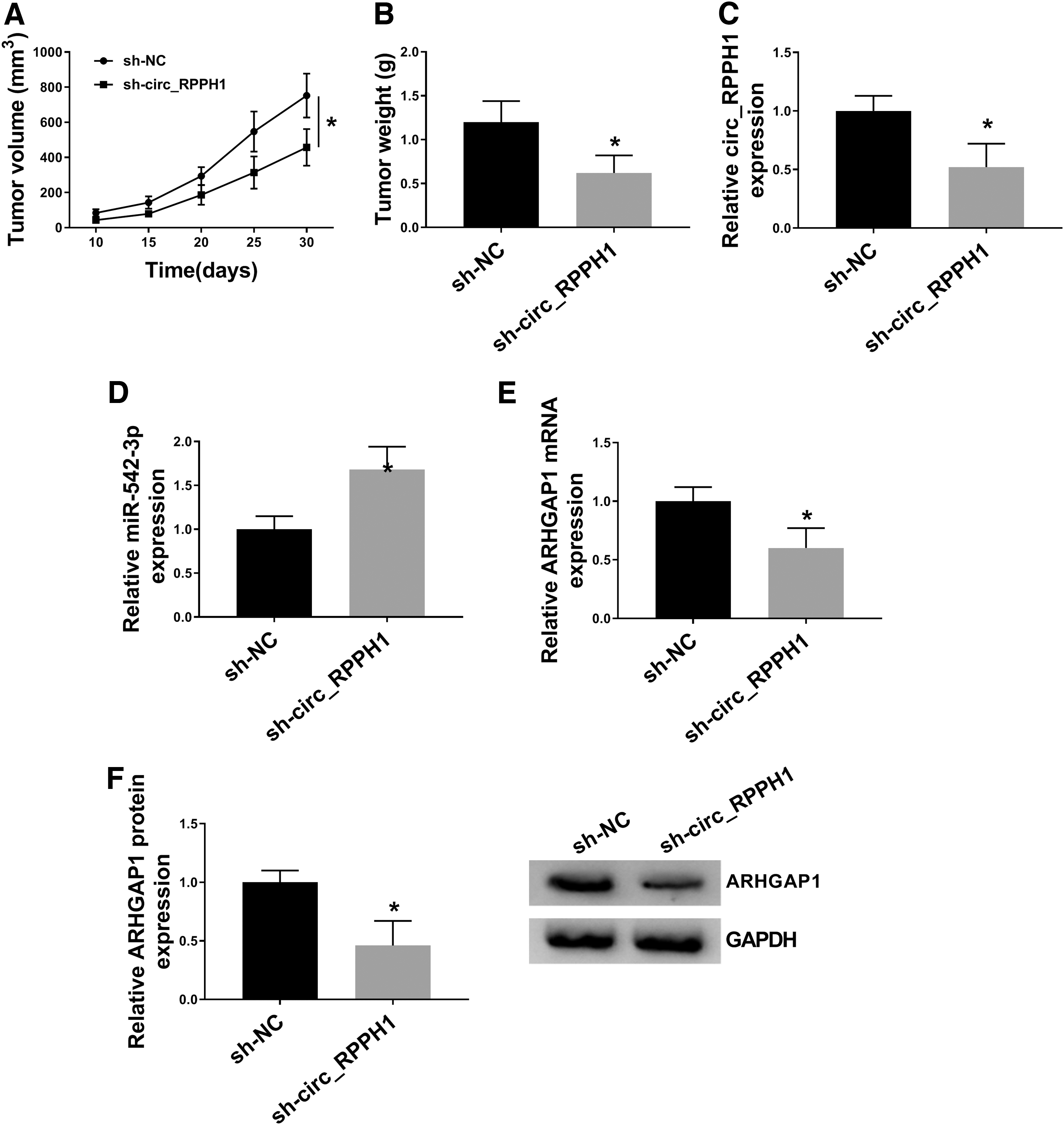
circ_RPPH1 facilitated BC tumor growth in vivo. MCF-7 cells stably expressing sh-NC or sh-circ_RPPH1 were implanted into BALB/c nude mice.
Discussion
In human diseases, circRNAs could serve as a group of biomarkers owing to their stability and function as pathophysiology controllers. 19 In this study, we detected the abnormal upregulation of circ_RPPH1 in BC tissues and cells. Its protumor roles in BC were proved to be related to miR-542-3p/ARHGAP1 axis.
According to microarray data (GSE101124) from the Gene Expression Omnibus (GEO) database, circ_RPPH1 (also called hsa_circ_0000515) was upregulated in BC tissues. Our data also confirmed its upregulation in BC tissues and cells, in concordance with former study. 8 To our knowledge, research about circ_RPPH1 is rare. Accumulating works suggested that numerous dysregulated circRNAs were implicated with BC progression, 20 which prompted us to figure out the impact of circ_RPPH1 on BC development. Function assays in vitro and in vivo were carried out, and the results demonstrated that circ_RPPH1 knockdown inhibited proliferation, migration, and invasion, and EMT, while apoptosis of BC cells in vitro was facilitated, and blocked tumor growth in vivo, acting as an oncogene.
circRNAs could exert their functions by regulating the activity and expression of miRNAs at the transcriptional level, acting as miRNA sponges. 21,22 We then searched the target miRNAs of circ_RPPH1 for better understanding the molecular mechanism of circ_RPPH1. Through online bioinformatics analysis and dual-luciferase reporter assay validation, miR-542-3p was confirmed to be a downstream miRNA of this circRNA.
Mapped in Xq26.3, miR-542-3p was identified to be a tumor suppressor in multiple human malignancies, such as osteosarcoma, 23 epithelial ovarian cancer, 24 hepatocellular carcinoma, 25 colon cancer, 26 nonsmall cell lung cancer, 27 and BC. 28 Apart from this, miR-542-3p was previously demonstrated to repress angiogenic activity of endothelial cells, tumor growth, and metastasis of mouse models by Angpt2. 29 Furthermore, co-delivery of doxorubicin and miR-542-3p from nanoparticle system PEI-PLGA was a promising therapy means for triple-negative BC. 30 Similar to anterior studies, 13,28 qRT-PCR assay in this project revealed that miR-542-3p was downregulated in BC. Moreover, gain-of-function showed that enforced expression of miR-542-3p obviously suppressed BC cell proliferation and metastasis. We also performed rescue assays, and found that miR-542-3p inhibition could weaken circ_RPPH1 knockdown-mediated suppressed impact on the malignant behaviors of BC cells. Thus, circ_RPPH1 exerted its procancer roles in BC by negatively regulating miR-542-3p expression.
It is well-documented that miRNAs could play vital regulatory roles by directly binding to mRNAs, causing post-transcriptional repression. 31,32 MiR-542-3p functioned as tumor suppressor by targeting various mRNAs, like ILK, 23 CDK14, 24 FTSJ2, 27 and S1PR1. 28 We discovered that miR-542-3p had complementary binding region with the 3′UTR of ARHGAP1. Dual-luciferase reporter assay testified the interaction between miR-542-3p and ARHGAP1.
ARHGAP1 had dual effects on ischemic cardiomyopathy (ICM), was a potential therapy target for ICM patients. 33 Targeted by miR-130b, ARHGAP1 was demonstrated to have clinical association with Ewing sarcoma patients. 34 Moreover, increased expression of ARHGAP1 had anti-tumor activity in cervical cancer, reflected by its inhibitory effects on cell proliferation, migration, and invasion. 35 ARHGAP1 was involved in the modulation of BC progression by serving as a target mRNA of miR-34b-5p, playing oncogenic role. 16 In sum, ARHGAP1 had multiple functions in human diseases. In this study, we also detected the apparent upregulation of ARHGAP1 in BC tissues and cells. Besides, accumulation of ARHGAP1 undermined the miR-542-3p-induced BC cell proliferation and metastasis inhibition. From our data, circ_RPPH1 positively regulated ARHGAP1 expression by absorbing miR-542-3p. Collectively, circ_RPPH1 contributed to BC development by regulating miR-542-3p/ARHGAP1 axis.
In this study, we first recognized a pro-BC circRNA, circ_RPPH1 in BC progression, whose depletion inhibited cell proliferation and metastasis of BC cells in vitro, and reduced tumor growth in vivo, probably by mediating miR-542-3p/ARHGAP1 pathway, at least. This study suggested that circ_RPPH1 was a novel target for BC treatment, and uncovered the potential mechanism.
Footnotes
Acknowledgments
The authors sincerely appreciate all members who participated in this study.
Authors' Contributions
Conception and design: L.Q. and B.S.; Development of methodology: L.Q., B.S. and B.Y.; Acquisition of data: L.Q. and B.S.; Analysis and interpretation of data: L.Q., B.Y. and S.L.; Writing, review, and revision of article: L.Q., B.S., B.Y., and S.L.; Administrative, technical, and material support: L.Q., B.S. and S.L.; Study supervision: L.Q. and B.Y. All coauthors have reviewed and approved of the article before submission.
Disclosure Statement
There are no existing financial conflicts.
Funding Information
This work was supported by The National Natural Science Foundation of China (No. 81001187).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.