
Abstract
The Editor-in-Chief of Cancer Biotherapy and Radiopharmaceuticals officially retracts the article entitled, “C Long Noncoding RNA CTD-2589M5.4 Inhibits Ovarian Cancer Cell Proliferation, Migration, and Invasion Via Downregulation of the Extracellular Matrix-Receptor Interaction Pathway,” by Yuanyuan Gu, Ke Huang, Min Zhang, Fang Teng, Lili Ge, Juan Zhou, Juan Xu, and Xuemei Jia (Cancer Biother Radiopharm, 37(7):2022;580–588; doi: 10.1089/cbr.2020.4429).
The corresponding author of the paper, Dr. Xuemei Jia, contacted the journal's editorial office requesting to replace Figure 3K in the published article, stating:
”An error was identified in Fig. 3K, which was made by careless figure assembling. These experiments were repeated by two students for at least three times….These corrections do not change the conclusion of this article. All authors agree to the correction and apologize for any inconvenience caused.” [sic]
An independent analysis of the figures was performed which confirmed image manipulation, raising doubts about the originality of data and the reliability of the published results.
Dr. Xuemei Jia, as well as all coauthors, were notified via email of the decision to retract, utilizing the email addresses provided to the journal at the time of submission in 2020, though no responses were received.
The Editor and Publisher of Cancer Biotherapy and Radiopharmaceuticals are committed to preserving the veracity of scientific literature and the community it serves.
Introduction
Malignant ovarian neoplasms are highly malignant tumors that rank as the fifth most prevalent reason for tumor mortality in women in the United States, with ∼14,000 deaths per year, 1 and only <40% of women with such neoplasms achieve remission. 2 Ovarian neoplasms are composed of several histopathologic entities, and therapy is dependent on the particular tumor type. 3 Epithelial ovarian cancer accounts for about 90% of malignant ovarian neoplasms. 4 –6 Despite the dramatic progress that has been achieved via surgery, chemotherapy, and radiotherapy treatments, the 5-year survival rate has remained ∼46.5%. 7 As such, a greater understanding of the pathological mechanisms and development of novel therapeutic approaches for ovarian cancer are urgently needed.
Complete genomic regulation involves a large number of noncoding RNAs (ncRNAs), a discovery that has revolutionized the understanding of the human genome. 8 Approximately 51,382 long noncoding RNAs (lncRNAs) are encoded in the human genome, which represent a significant category of ncRNAs 9 and transcripts longer than 200 nucleotides that do not have the prominent signature motifs characteristic of mRNAs. 10 It is believed that lncRNAs participate in several physiological and pathological processes, including maintaining epithelial cell homeostasis, 11 vascularization, 12 Alzheimer's disease, 13 and tumorigenesis. 14 An increasing number of studies have shown that lncRNAs act by affecting the transcriptional machinery, translation, and chromatin organization. 15 In ovarian cancer, previous studies have reported that the lncRNAs UCA1, 16 CTD-2020K17.1, 17 and HOTAIR 18 are associated with the development of ovarian cancer and poor survival of these patients.
In our previous study, the authors unexpectedly identified the multidrug-resistant lncRNA CTD-2589M5.4 via high-throughput sequencing and provided results clarifying its role in the chemoresistant process. 19 While biased expression of lncRNAs has been shown to be involved in multiple diseases, limited information regarding the detailed molecular mechanisms by which they act in various disease processes is still limited. 20 In addition, the role that lncRNA CTD-2589M5.4 plays in the development and/or progression of ovarian cancer remains unclear.
The current study sought to detect the expression and function of CTD-2589M5.4 in ovarian tumors. It was shown that CTD-2589M5.4 expression levels were downregulated in primary ovarian cancer tissues (POCTs) and ovarian cancer cell lines. Moreover, knockdown of CTD-2589M5.4 enhanced the proliferation, migration, and invasion of ovarian cancer cells. To further explore the mechanisms of CTD-2589M5.4 function, the differentially expressed mRNAs in CTD-2589M5.4-overexpressed and control cells were screened via high-throughput RNA sequencing. In addition, Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis was used to analyze the possible function of these mRNAs. The results indicated that CTD-2589M5.4 likely has a significant role in the progression of ovarian cancer.
Materials and Methods
Clinical specimens
Thirty-three samples of ovarian tissue specimens, including 16 normal ovarian tissues (NOTs) and 17 POCTs, were acquired from patients who had previously undergone surgery between January 2016 and November 2017 at the Women's Hospital of Nanjing Medical University (Nanjing, China). Detailed information regarding these patients is summarized in Table 1. NOTs were acquired from patients who had undergone an ovariotomy due to the presence of cervical cancer. Each sample was diagnosed by at least two professional pathologists. All fresh tissue specimens were flash-frozen in liquid nitrogen and then frozen at −80°C before RNA extraction. No patient had preoperative treatment before the surgery. This study was approved by the Human Ethics Committee of Women's Hospital of Nanjing Medical University and all participants signed informed consent forms (Nanjing, China).
Clinical Characteristic of the Ovarian Cancer Patients Used in This Study
Cell culture
SKOV3, A2780, HO8910, and IOSE80 cells were purchased from Jiangsu KeyGEN BioTECH Corp., Ltd. (China). OVCAR3 cells were purchased from Shanghai Mingjin Biotech Corp., Ltd. (China). A2780, OVCAR3, HO8910, and IOSE80 were all maintained in Dulbecco's modified Eagle's medium (KeyGEN BioTECH, Nanjing, China) replenished with 10% fetal bovine serum (FBS; Invitrogen, Carlsbad, CA). SKOV3 was maintained in McCoy's 5A medium (KeyGEN BioTECH) supplemented with 10% FBS. All cells were cultivated in a humidified incubator at 37°C with 5% CO2. For the rescue experiments, fibronectin, purchased from Biological Industries (Israel), was diluted in phosphate-buffered saline (PBS) to a concentration of 20 ng/mL, and six-well plates were coated with this solution and incubated overnight at 4°C before use.
RNA extraction and real-time quantitative PCR
For RNA isolation, total RNA from the tissue specimens and cells was extracted using TRIzol reagent (Invitrogen). The high-quality RNA was reverse-transcribed into cDNA using a PrimeScript TMRT reagent kit (Takara Bio, China) following the manufacturer's instructions. CTD-2589M5.4 levels were evaluated by real-time quantitative PCR (RT-qPCR) using the SYBR Green method. The primer sequences were as follows: CTD-2589M5.4 Forward: 5′-TAAAGGCATCCCAACGCAGT-3′ and CTD-2589M5.4 Reverse: 5′-CCACACTCCGGAAAAGCTCA-3′; fibronectin 1 (FN1) Forward: 5′-TGGGCAACTCTGTCAACGAA-3′, FN1 Reverse: 5′-GAGCAAATGGCACCGAGATA-3′; SV2A Forward: 5′-TCTGCTCATCTCGCTCTCAG-3′, SV2A Reverse: 5′-GTACACGCCACCAATCATCC-3′; ITGB8 Forward: 5′-ACCAGGAGAAGTGTCTATCCAG-3′, ITGB8 Reverse: 5′-CCAAGACGAAAGTCACGGGA-3′. The primers specific to β-actin were as follows: 5′-AGCGAGCATCCCCCAAAGTT-3′ (forward) and 5′-GGGCACGAAGGCTCATCATT-3′ (reverse). The lncRNA or mRNA expression was calculated using the 2−△ct method, with ΔCT = CT (target) − CT (β-actin).
Western blot
For Western blot analysis, both the control and CTD-2589M5.4-overexpressed SKOV3 cells were lysed in RIPA buffer, including 0.1 mM phenylmethylsulfonyl fluoride (PMSF), and then centrifuged. The protein concentration was determined using bicinchoninic acid analysis (Pierce, Rockford). The protein in the cell lysates (30 μg/lane) was separated via sodiumdodecyl sulfate polyacrylamide gel electrophoresis and then transferred to polyvinylidence difluoride membranes (Millipore, Billerica). The membranes were then incubated with the primary antibodies overnight at 4°C after blocking in 5% milk. Protein expression was detected after incubating with horseradish peroxidase-conjugated secondary antibodies (Biosharp, Nanjing, China) and visualization using enhanced chemiluminescent reagents. The antibodies used in this study were as follows: anti-FN1 (15613-1-AP; Proteintech, Wuhan, China), anti-ITGB8 (DF2545; Affinity Biosciences), and anti-SV2A (DF13377; Affinity Biosciences).
Vector construction and cell transfection
The CTD-2589M5.4 overexpression plasmid and empty plasmid, pcDNA3.1, were purchased from Nanjing Genebay Biotech Co., Ltd (Nanjing, China). The CTD-2589M5.4 plasmid and empty plasmid were transfected into cells using Lipofectamine 2000 reagent (Invitrogen) following the manufacturer's instructions. The cells were then subdivided into the CTD-2589M5.4-overexpression group and the negative control (NC) group. Three shRNAs targeting CTD-2589M5.4 and a scrambled shRNA, which served as a NC, were synthesized by Genomeditech (Shanghai, China). The sequences of shRNAs were: sh-CTD-2589M5.4-1: 5′-GGATATGAACAGCCGTCTTGG-3′, sh-CTD-2589M5.4-2: 5′-GCTGGATGTCACTTACTAGAG-3′, sh-CTD-2589M5.4-3: 5′-GGAGGGCATCGGTTAGAAGGA-3′, sh-NC: 5′-TTCTCCGAACGTGTCACGT-3′. The shRNA plasmids and control plasmids (pGMLV-SC5) were transfected into the SKOV3 and A2780 cells using Lipofectamine 2000 reagent. Forty-eight hours after transfection, the cells were isolated for use in subsequent assays.
Transwell migration and invasion assays
To explore the migration and invasion capacity of SKOV3 and A2780 cells, transwell assays were conducted. A total of 3 × 104 transfected cells per well were plated in the upper chambers of the transwells with serum-free media for analysis of migration. The bottom chambers were filled with media supplemented with 20% FBS. An invasion assay was carried out using the transwell apparatus with Matrigel-coated (BD, Franklin Lakes, NJ) filters, following the manufacturer's instructions. After the cells were transfected, they were seeded at 6 × 104 cells per well in the upper chambers with serum-free media. The lower chambers included media supplemented with 20% FBS. After the transwell apparatus was cultivated in a humidified incubator at 37°C with 5% CO2 for 48 h, the upper chamber filters were fixed and dyed with 1% crystal violet. The cells on the bottom of the membrane were observed and photographed under an inverted microscope. Finally, the cells were lysed in RIPA buffer, including 0.1 mM PMSF. The cell lysate from each well was divided into three parts and quantified by measuring the absorbance at 562 nm to evaluate the relative amount of cells.
Cell proliferation and cell cycle analyses
For analyzing the proliferation capacity of the SKOV3 and A2780 cells after transfection, CCK8 assays (Beyotime, Shanghai, China) were performed. In brief, 1.5 × 103 cells were seeded into each well in 96-well plates and then cultivated for 24, 48, and 72 h. At each time point, 10 μL of CCK8 was added to each well. After 1 h of cultivation, the absorbance at 450 nm was measured to assess the cell proliferative capacity using a microplate reader.
After transfection for 48 h, the cells were treated with ethylenediaminetetraaceticacid-free trypsin for cell cycle detection. The cells were washed with PBS and fixed in 70% ice-cold ethanol overnight at 4°C. Then the cell cycle was analyzed using a cell cycle and apoptosis analysis kit (Beyotime) according to the manufacture's protocol. In brief, after being washed twice in PBS, the cells were maintained in 500 μL binding buffer containing 25 μL propidium iodide and 10 μL RNase A in the dark for 30 min at 37°C. Subsequently, the cell cycle was investigated using a flow cytometer (BD Bioscience). The data were analyzed using Modfit 5.0 software.
Wound-healing assay
To analyze the wound-healing capacity, 5 × 105 transfected cells per well were reseeded in a six-well plate and cultivated to confluence. Factitious homogeneous wounds were formed using a sterilized 200 μL micropipette tip to scrape through the cell layer. Subsequently, the cells were maintained with serum-free medium. Cell-free areas were photographed at 0, 24, and 48 h using a light microscope and measured using ImageJ software.
mRNA sequencing
Total RNA was extracted from the CTD-2589M5.4 and NC groups of A2780 cells using TRIzol following the instructions of the manufacturer. The RNA quality, 28s/18s rRNA ratio, and RNA concentration were evaluated using an Agilent 2100 Bioanalyzer (Agilent Technologies, CA). Three RNA specimens for each group were sent to CapitalBio Technology Co, Ltd. (Beijing, China) for mRNA deep sequencing on a Hiseq2500 X-10 platform. In brief, the mRNA library was constructed after rRNA depletion, removal of all trace DNA, fragmentation, first-strand cDNA synthesis, second-strand cDNA synthesis, end repair, 3′ end A-tailing, linker ligation with adapters, and RNA enrichment using a NEBNext Ultra RNA Library Prep Kit for Illumina (cat E7530s; New England Biolabs, China) according to the instruction manual. Subsequently, the qualified library was subjected to analysis on an Illumina Hiseq2500 X-10 platform.
KEGG pathway analysis
To identify the underlying mechanism of CTD-2589M5.4 function in ovarian cancer progression, KEGG pathway analysis was used for analyzing the primary functions of the differentially expressed genes between CTD-2589M5.4 overexpression and control A2780 cells based on the KEGG database. The p-values and adjusted p-values revealed the significance of the pathway.
Statistical analysis
The quantitative data are expressed as the mean ± standard error of the means. Statistical analysis was performed using the Student's t-test or one-way analysis of variance via the GraphPad Prism 5.0 software (GraphPad Software, La Jolla, CA). p-values <0.05 were considered statistically significant.
Results
CTD-2589M5.4 is significantly downregulated in ovarian cancer tissues and ovarian cancer cells
To investigate the role of CTD-2589M5.4 in ovarian cancer progression, 17 POCTs and 16 NOTs were collected. The clinical characteristics of the patients are listed in Table 1. The expression level of the lncRNA CTD-2589M5.4 in ovarian cancer tissues and cell lines were determined via RT-qPCR. The results indicated that CTD-2589M5.4 was dramatically downregulated in POCTs (n = 17) compared to NOTs (n = 16) (p < 0.05; Fig. 1A). In addition, CTD-2589M5.4 expression was also downregulated in ovarian cancer cells (HO8910, OVCAR3, A2780, and SKOV3) compared with normal ovarian epithelium cells (IOSE80) (Fig. 1B).

lncRNA CTD-2589M5.4 was downregulated in ovarian cancer tissues and cell lines.
Knockdown of CTD-2589M5.4 enhances the proliferation, invasion, and migration of ovarian cancer cells
To further investigate the potential function of CTD-2589M5.4 in regulating ovarian cancer progression, the sh-CTD-2589M5.4-1, sh-CTD-2589M5.4-2, and sh-CTD-2589M5.4-3 plasmids were constructed and individually transfected into SKOV3 and A2780 cells. RT-qPCR indicated that all three targeted shRNAs were effective at silencing CTD-2589M5.4 in both cell lines (Fig. 2A, B). The knockdown efficiency of sh-CTD-2589M5.4-2 was the highest, and this was thereby selected for the subsequent studies. CCK8 assays revealed that cell proliferation was enhanced after CTD-2589M5.4 knockdown in both SKOV3 and A2780 cells (Fig. 2C, D). In addition, the cell invasion and migration abilities were also significantly elevated in SKOV3 and A2780 cells following CTD-2589M5.4 knockdown, as based on transwell and wound-healing assays (Fig. 2E–P).
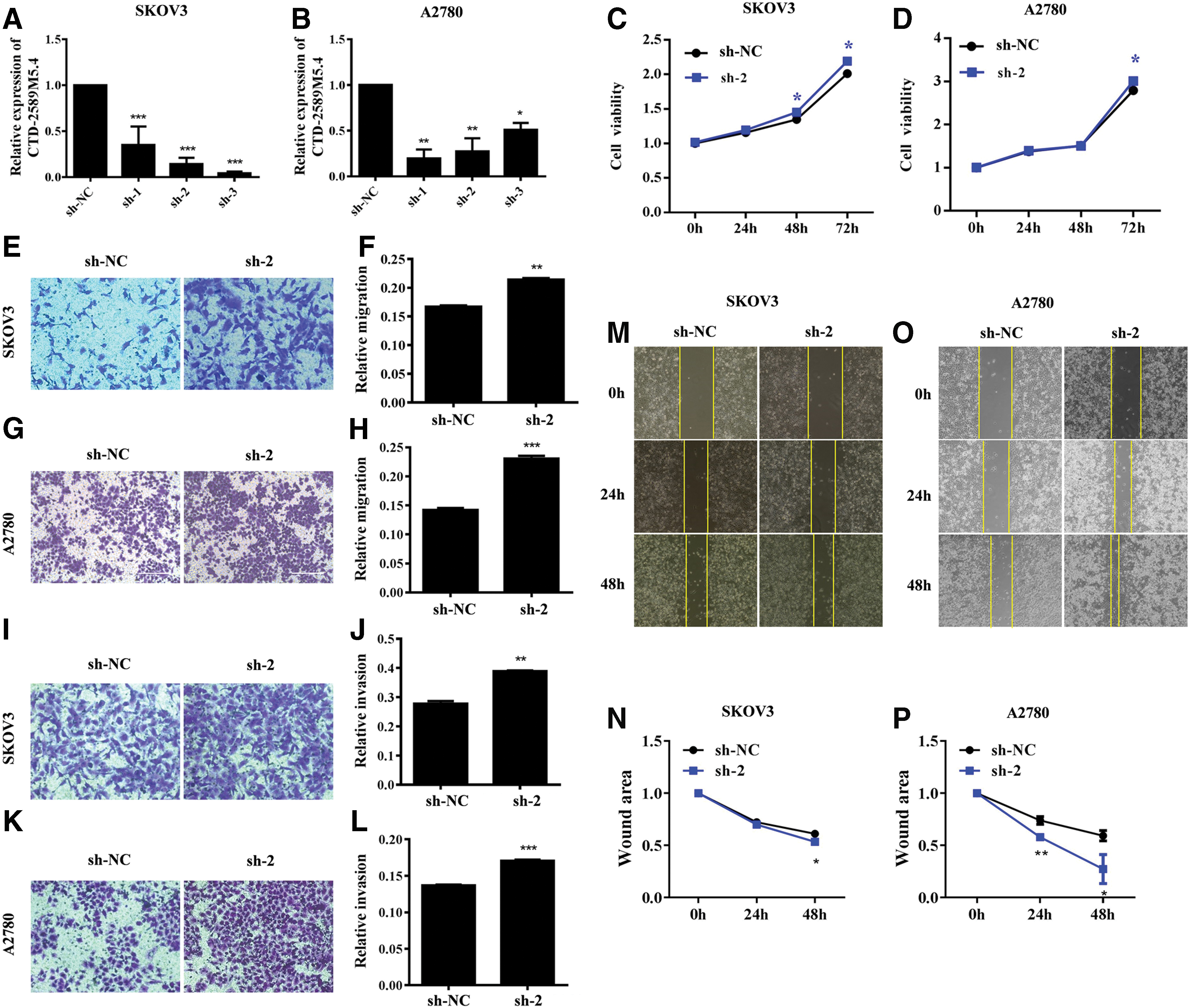
Knockdown of CTD-2589M5.4 enhances cell migration, invasion, and proliferation of SKOV3 and A2780 cells.
Overexpression of CTD-2589M5.4 represses ovarian cancer cell proliferation, invasion, and migration
To further demonstrate the tumor-suppressive function of CTD-2589M5.4, a CTD-2589M5.4 overexpression vector was also constructed. Analysis via RT-qPCR showed that CTD-2589M5.4 expression was markedly increased in the CTD-2589M5.4 overexpression group at 48 h after transfection with the CTD-2589M5.4 overexpression vector (p < 0.01, Fig. 3A, B). The results of CCK8 assays indicated that the cell proliferation ability was repressed in CTD-2589M5.4-overexpressed SKOV3 and A2780 cells (Fig. 3C, D). A notable difference was viewed at 72 h between the CTD-2589M5.4-overexpressed group and NC group. In addition, flow cytometry analysis revealed that the number of cells in S-phase was notably reduced in CTD-2589M5.4-overexpressed SKOV3 and A2780 cells, while the number of cells in the G2-phase was markedly enhanced (Fig. 3E, F).

Overexpression of CTD-2589M5.4 represses cell migration, invasion, and proliferation of A2780 and SKOV3 cells.
The results of transwell and wound-healing assays showed that the cell invasion and migration ability was significantly repressed in CTD-2589M5.4-overexpressed SKOV3 and A2780 cells compared to the control cells (p < 0.05, Fig. 3G–R).
Differentially expressed mRNAs in CTD-2589M5.4-overexpressed A2780 cells and control A2780 cells
To investigate the possible mechanism of CTD-2589M5.4 function in ovarian cancer cells, high-throughput RNA-sequencing of CTD-2589M5.4-overexpressed A2780 and control A2780 cells was performed. A total of 750 mRNAs and 233 mRNAs were shown to be upregulated and downregulated in the CTD-2589M5.4-overexpressed A2780 cells, respectively (fold change ≥2, q < 0.05) (Fig. 4A, B and Supplementary Table S1).
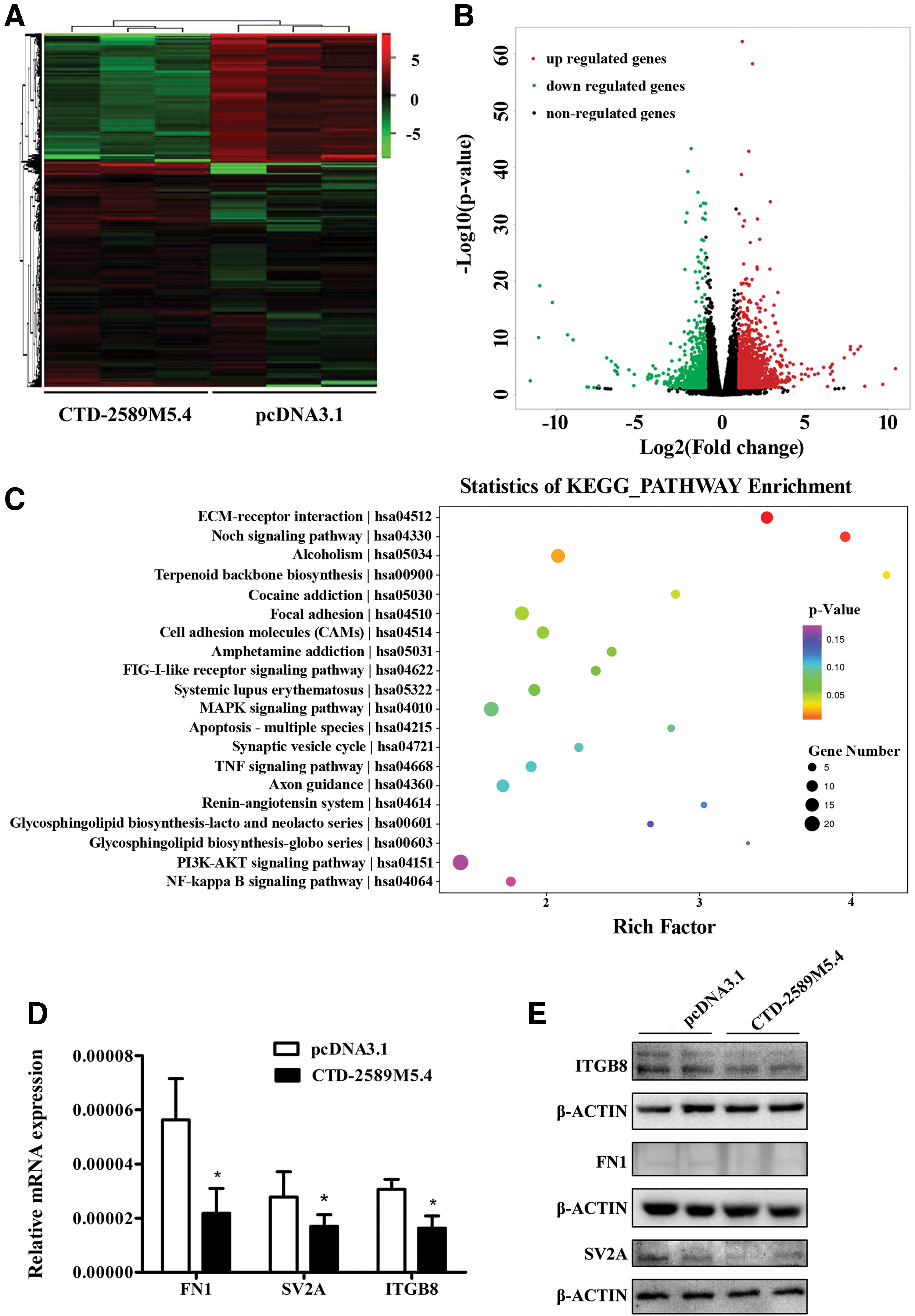
The extracellular matrix–receptor signaling pathway was significantly downregulated in CTD-2589M5.4-overexpressed ovarian cancer cells.
CTD-2589M5.4 inhibits ovarian cancer progression, possibly via downregulation of the extracellular matrix–receptor signaling pathway
KEGG pathway analysis indicated that the differentially expressed mRNAs were mainly enriched for the following signaling pathways: extracellular matrix (ECM)–receptor interaction, Notch signaling pathway, alcoholism, terpenoid backbone biosynthesis, focal adhesion, and cell adhesion molecules (Fig. 4C).
The authors noticed that the ECM–receptor interaction pathway was the most dysregulated pathway in the CTD-2589M5.4-overexpressed A2780 cells. The genes in the ECM–receptor interaction pathway are typically dysregulated in ovarian cancers, and their high expression indicates a poor prognosis for patients with different cancers, including gastric cancer, lung cancer, colorectal cancer, and breast cancer. 21 –25 In addition, subsequent RT-qPCR analysis indicated that FN1, SV2A, and ITGB8 were all significantly decreased in CTD-2589M5.4-overexpressed A2780 cells, which was consistent with the RNA-sequencing data (Fig. 4D). Western blot analysis of these three proteins confirmed these results (Fig. 4E). Since FN1, which encodes fibronectin, is one of the most important components of the ECM, we performed a rescue experiment by adding fibronectin to CTD-2589M5.4-overexpressed A2780 cells. Importantly, the application of ectopic fibronectin eliminated the tumor suppressive function of CTD-2589M5.4 in ovarian cancer cells (Fig. 5A–D). In conclusion, these results indicated that CTD-2589M5.4 might serve as a tumor suppressor in ovarian cancer cells by downregulating the ECM–receptor interaction pathway.
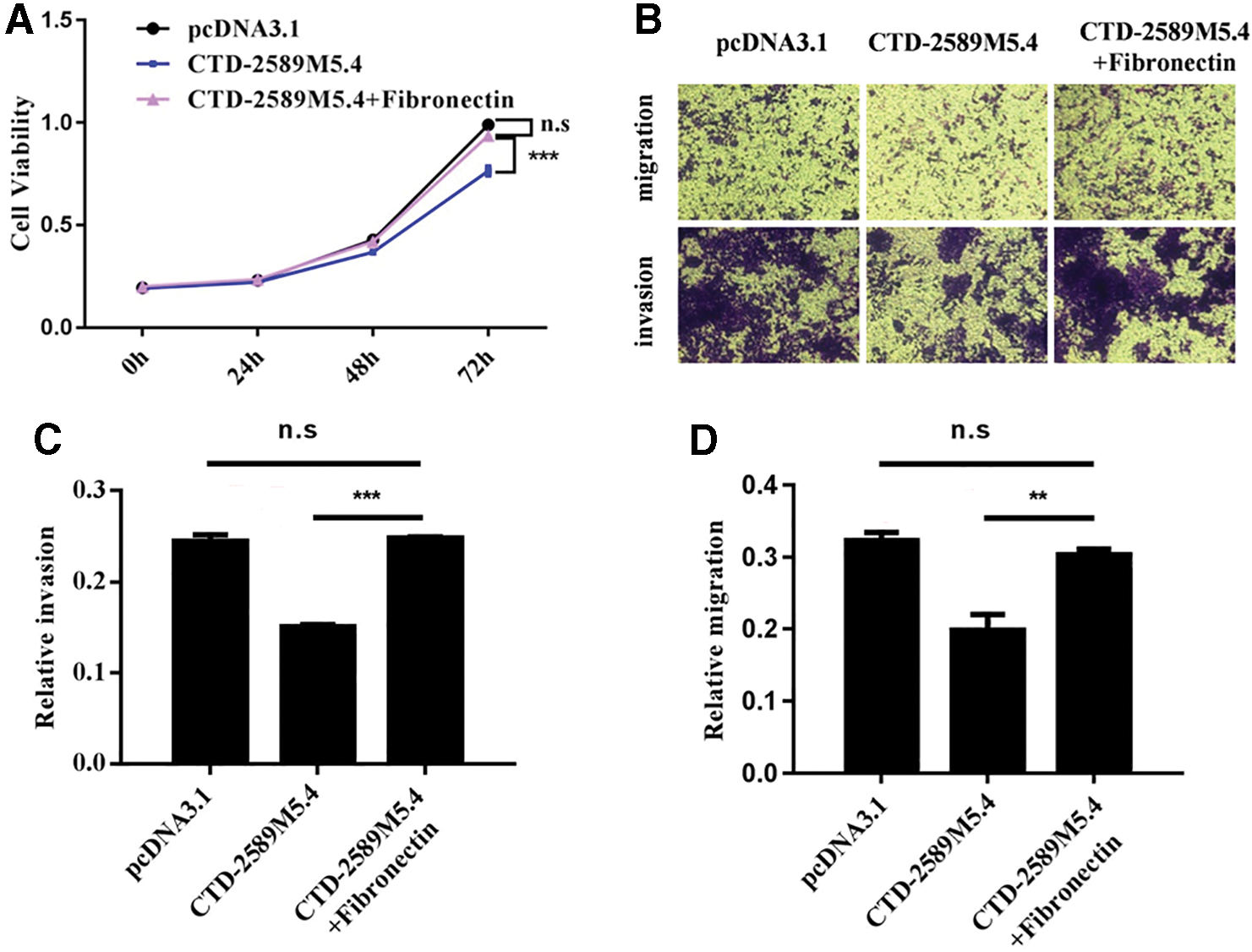
The tumor suppressive function of CTD-2589M5.4 can be rescued by fibronectin.
Discussion
In the previous study, we assessed for differentially expressed lncRNAs in paclitaxel (PTX)-resistant A2780 cells and PTX-sensitive A2780 cells via RNA-sequencing analysis, and this analysis identified the novel lncRNA CTD-2589M5.4. The authors' subsequent study demonstrated that CTD-2589M5.4 was downregulated in ovarian cancer tissues and cell lines. In this current study, we demonstrated that CTD-2589M5.4 could inhibit the proliferation, migration, and invasion of ovarian cancer cells. Moreover, the differentially expressed mRNAs in CTD-2589M5.4-overexpressed ovarian cancer cells and control ovarian cancer cells were identified, and the results indicated that CTD-2589M5.4 might inhibit ovarian cancer progression via downregulation of the ECM–receptor interaction pathway. However, how CTD-2589M5.4 functions to downregulate the ECM-receptor pathway still requires further study.
Currently, many differentially expressed lncRNAs have been implicated in oncogenesis and tumor development, 26 including the upregulated lncRNAs HOTAIR and H19 27 and downregulated lncRNAs HOXA11-AS 28 and BC200 29 in ovarian cancer. LncRNAs play a pivotal role in the process of many tumors, indicating that they might serve as potential diagnostic biomarkers for use in clinical therepy 30 and/or therapeutic targets for inhibiting cancer progression. 31 Based on the outcomes of this current study, loss- and gain-of-function experiments indicated that CTD-2589M5.4 negatively regulates ovarian cancer cell viability via downregulation of the ECM–receptor interaction pathway, thereby suggesting that CTD-2589M5.4 could be a potential therapeutic target for the prevention and/or treatment of ovarian cancer.
However, the upregulation of CTD-2589M5.4 in chemoresistant ovarian cancer cells was in opposition to the proposed tumor-suppressive function of CTD-2589M5.4 during ovarian cancer progression. Interestingly, it was noticed that there was a proportion of ovarian cancer patients who progressed rapidly but were also sensitive to drug treatments, which correlated with a better prognosis. As such, CTD-2589M5.4 downregulation in ovarian cancer cells might contribute to a more rapid progression but a better prognosis.
The ECM is a macromolecular network that modulates the cellular response under both normal and pathological conditions. The ECM is composed of fibronectin, collagen, laminin, vitronectin, and several other glycoproteins. The matrix components bind with cell surface receptors, such as fibronectin binding to ITGB1 and laminin binding to SV2A, thus influencing the progression of cancers. In addition, the ECM is essential to providing mechanical support to tissues and regulating the Notch signaling pathway, 32 which was also shown to be significantly altered in the RNA sequencing data. Moreover, the genes comprising the ECM–receptor interaction pathway are frequently altered in cancer patients. For example, FN1 and ITGB1 are significantly upregulated in ovarian cancer, and their high expression often indicate a poor prognosis. 22,33 However, the upregulation of FN1 and ITGB1 also promotes the chemoresistance of cancer cells, which could indicate that different mechanisms might underlie the function of CTD-2589M5.4 in ovarian cancer progression and chemoresistance.
Since surgery and platinum-based chemotherapies are still the main clinical treatments used to treat ovarian cancer, the study of ovarian cancer should focus on the function of genes and lncRNAs in relationship to the progression of ovarian cancer as well as their function with regard to chemoresistance.
Financial Disclosure
There are no financial conflicts of interest to disclose.
Footnotes
Disclosure Statement
There are no existing financial conflicts.
Funding Information
The study was financially supported by the National Natural Science Foundation of China (Grant number: 81572556), Nanjing Medical Science and Technique Development Foundation (Grant numbers: ZKX15046, ZDX16015, QRX17159).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.