
Abstract
Background:
Nonsmall cell lung carcinoma (NSCLC) is a major cause of cancer-related death worldwide. The resistance of NSCLC to chemical drugs, such as cisplatin (CDDP), poses a heavy burden for NSCLC therapy. Herein, the effects of circular_0008928 (circ_0008928) on the CDDP sensitivity and biological behavior of CDDP-resistant NSCLC cells and underlying mechanism are revealed.
Materials and Methods:
The expression of circ_0008928 and microRNA-488 (miR-488) was detected by quantitative real-time polymerase chain reaction. The expression of hexokinase 2 (HK2) protein and exosome-specific proteins was determined by Western blot. The half-maximal inhibitory concentration (IC50) value of CDDP was detected by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Cell proliferation and migratory and invasive abilities were illustrated by cell counting kit-8 and transwell assays. Cell glycolysis metabolism was illustrated by extracellular acidification rate assay, glucose kit and lactate kit assays and Western blot analysis. The binding sites between miR-488 and circ_0008928 or HK2 were predicted by starbase or microT-CDS online database, and identified by dual-luciferase reporter and RNA immunoprecipitation assays.
Results:
Circ_0008928 expression and HK2 protein expression were significantly upregulated, while miR-488 expression was obviously downregulated in NSCLC cells and CDDP-resistant NSCLC cells. Circ_0008928 expression was increased in serum exosomes of CDDP-resistant NSCLC patients compared with CDDP-sensitive NSCLC patients. In addition, circ_0008928 silencing improved CDDP sensitivity and attenuated CDDP-induced cell proliferation, migration, invasion, and glycolysis metabolism. Circ_0008928 was a sponge of miR-488, and miR-488 bound to HK2 in CDDP-resistant NSCLC cells. Furthermore, both miR-488 inhibitor and HK2 overexpression attenuated circ_0008928 absence-mediated impacts on CDDP sensitivity and tumor process in CDDP-resistant NSCLC.
Conclusions:
Circ_0008928 knockdown improved CDDP sensitivity and hindered cell proliferation, migration, invasion, and glycolysis metabolism by miR-488/HK2 axis in CDDP-resistant NSCLC. This finding provides a new mechanism for studying CDDP-resistant therapy in NSCLC.
Introduction
Lung cancer is a major cause of cancer-linked mortality worldwide. 1 Lung cancer patients present a poor prognosis with 5-year survival rate <20%. 2 Nonsmall cell lung carcinoma (NSCLC) accounts for more than half of lung cancers, and its morbidity ranks the second and fatality rate displays the first worldwide. 3,4 At present, chemotherapy is the primary treatment of NSCLC, and cisplatin (CDDP), a platinum drug, is routinely used to treat NSCLC. 5 However, >60% NSCLC patients have been reported to resist CDDP. 6 Thus, defining the mechanism of NSCLC resistance to CDDP is urgent.
Circular RNA (circRNA) is a novel noncoding RNA and is more stable than its linear form based on its closed loop. 7,8 CircRNA expression is often dysregulated in cancers. Multiple data showed that circRNA was involved in the occurrence and development of cancers, including NSCLC. 9,10 Previous study also reported that circRNA could mediate drug resistance in NSCLC progression. 11 For example, Li et al. indicated that enforced circ_0002483 expression suppressed cell proliferation and invasion, and inhibited taxol resistance in NSCLC. 12 Huang et al. also explained that circ_0001946 silencing contributed to cell proliferation, metastasis, and resistance to CDDP, and promoted cell apoptosis in NSCLC. 13 However, the effects and mechanism of circ_0008928 in NSCLC sensitivity to CDDP have not been addressed.
MicroRNA (miRNA) is a small noncoding RNA and regulates many biological behavior, such as cell progression, 14 glycolysis metabolism, 15 and drug sensitivity 16 in cancer progression. Present research studies revealed that miR-488 could inhibit NSCLC progression. For instance, Wang et al. revealed that miR-488 upregulation restrained cell proliferation and metastasis, and contributed to cell apoptosis in NSCLC 17 ; Fang et al. also demonstrated that miR-488 repressed cell proliferative ability and cell sensitivity to CDDP in NSCLC. 18 The above data implicate that miR-488 may play a vital part in NSCLC process.
The expression of hexokinase 2 (HK2), a rate-limiting enzyme of glycolysis, has been found constantly dysregulated in various cancers. 19 Research studies indicated that HK2 played a crucial part in the occurrence and maintenance of cancers, and HK2 or HK2-based glycolysis could be employed as a target in treating cancers. 20,21 HK2 was disclosed to suppress cell apoptosis through interacting with voltage-dependent anion channel. 22 In addition, Wang et al. explained that HK2 silencing hindered NSCLC growth. 23 Therefore, HK2 is an important object in revealing the pathogenesis of NSCLC.
Herein, the expression of circ_0008928, miR-488, and HK2 was determined in CDDP-resistant NSCLC cells. The effects of circ_0008928 on CDDP sensitivity, NSCLC process, and glycolysis metabolism were disclosed. Rescue experiments were employed to identify that circ_0008928 regulated CDDP sensitivity, tumor process, and glycolysis metabolism by miR-488/HK2 axis in CDDP-resistant NSCLC. Furthermore, circ_0008928 expression was detected in serum exosomes of CDDP-resistant NSCLC patients and CDDP-sensitive NSCLC suffers.
Materials and Methods
Serum collection and storage
Twenty-eight CDDP-sensitive NSCLC patients, 19 CDDP-resistant NSCLC patients, and 47 volunteers without NSCLC provided serum with signing the written informed consents. The clinicopathologic features of NSCLC suffers are shown in Supplementary Table S1. NSCLC patients aged >50 accounted for 57.1% in CDDP-sensitive group, which was higher than those found in CDDP-resistant group. In addition, the tumor size >3 cm accounted for 52.6% in CDDP-resistant group, but accounted for 39.3% in CDDP-sensitive group. NSCLC patients were from Affiliated Hospital of Xiangnan University. Serum was instantly kept at 4°C in a refrigerator. The Ethics Committee of Affiliated Hospital of Xiangnan University agreed with this study.
Cell culture and establishment of CDDP-resistant NSCLC cells
Human NSCLC cell lines (A549, H1299, H661, and SK-MES-1), human normal bronchial epithelial cell line 16HBE, and human embryonic kidney cell line 293T were obtained from Sciencell (Carlsbad, CA). Cells were cultured in Roswell Park Memorial Institute-1640 (RPMI-1640; Biosun, Shanghai, China) or Dulbecco's modified Eagle's medium (Biosun) with 10% fetal bovine serum (FBS; Biosun) with 1% streptomycin/penicillin under 37°C in moist air with 5% CO2. CDDP-resistant A549 cells (A549/CDDP) were purchased from Sciencell (Carlsbad).
CDDP-resistant H1299 cells (H1299/CDDP) were established as below. H1299 cells were seeded in 96-well plate for 16 h and incubated with the 1/50 half-maximal inhibitory concentration (IC50) of CDDP (Beyotime, Jiangsu, China). After their stable growth, the cells were incubated with an increasing dose of CDDP in multiples for 15 d, which lasted >6 months.
Plasmid construction and cell transfection
The small interfering RNAs against circ_0008928 (si-circ_0008928#1, si-circ_0008928#2, and si-circ_0008928#3), miR-488 mimic (miR-488), the overexpression plasmids of circ_0008928 (circ_0008928) and HK2 (HK2), miR-488 inhibitor (in-miR-488) and control groups (si-con, miR-con, pCD5-ciR, pcDNA, and in-miR-con) were constructed by Invitrogen (Carlsbad, CA). Plasmids were transfected into A549/CDDP and H1299/CDDP cells using Lipofectamine 2000 (Invitrogen). The synthesized sequences were si-circ_0008928#1 5′-ACCGCACACAGGTTTGATGAT-3′, si-circ_0008928#2 5′-AACCGCACACAGGTTTGATGA-3′, si-circ_0008928#3 5′-CCGCACACAGGTTTGATGATT-3′, miR-488 mimic 5′-UUGAAAGGCUAUUUCUUGGUC-3′, miR-488 inhibitor 5′-GACCAAGAAAUAGCCUUUCAA-3′, si-con 5′-GGAATAATCAAGCTGGGAA-3′, miR-con 5′-UUUGUACUACACAAAAGUACUG-3′ and in-miR-con 5′-CAGUACUUUUGUGUAGUACAAA-3′.
Quantitative real-time polymerase chain reaction
NSCLC cells, 16HBE, CDDP-resistant NSCLC cells, and serum exosomes were lysed using TransZol (TransGen, Beijing, China) or GenElute™ RNA isolation kit (Sigma, St Louis, MO), and total RNA was extracted with RNAsimple kit (Tiangen, Beijing, China). cDNA was synthesized using FastKing RT Kit (Tiangen) or MicroRNA RT Kit (Thermo Fisher, Waltham, MA). Then, SuperReal PreMix Color (Tiangen) was used to detect the amount of circRNA/miRNA/mRNA. Data were assessed with the 2−ΔΔCt method. U6 and β-actin were employed as references. The sense and antisense primers were circ_0008928 5′-AGCCAACCAACTCTTCCG-3′ and 5′-TCCAACTGCTCCACGACA-3′; miR-488 5′-CTCACGAAGAAAGGCAAA-3′ and 5′-ACCTCAAGAACAGTATTTCCAGG-3′; SATB homeobox 2 (SATB2) 5′-CTGCGTCTTCTCGGCTCTTG-3′ and 5′-CGTTCTGGAGAGAAAGGGCT-3′; HK2 5′-TGTGAATCGGAGAGGTCCCA-3′ and 5′-GGAGACGTGATTTTGGCGTC-3′; U6 5′-TGCGGGTGCTCGCTTCGGCAGC-3′ and 5′-GTGCAGGGTCCGAGGT-3′; β-actin 5′-CTCCATCCTGGCCTCGCTGT-3′ and 5′-GCTGTCACCTTCACCGTTCC-3′.
Cytoplasmic and nuclear RNA analysis of quantitative real-time polymerase chain reaction
The location of circ_0008928 in cell was analyzed by PARIS™ Kit (Thermo Fisher). In short, fresh A549/CDDP and H1299/CDDP cells were collected and washed using phosphate buffer solution (PBS; Solarbio, Beijing, China). Cells were suspended in 400 μL cold buffer (Thermo Fisher), and then placed on ice for 6 min. The cells were centrifuged at 450 rpm for 5 min. The supernatant was carefully separated from nuclear pellet. Cell disruption buffer (Thermo Fisher) was added, and RNA was extracted. Results were assessed using quantitative real-time polymerase chain reaction (qRT-PCR). β-actin and U6 were selected as references.
Identification of circRNAs circular characteristic
Random primers and oligo (dT)18 primers were synthesized from GENEWIZ (Suzhou, China), and were employed to identify the circular characteristic of circ_0008928 based on oligo (dT)18 primers only amplifying RNA containing poly (A) tails and random primers amplifying all RNA. In short, RNA from A549/CDDP and H1299/CDDP cells was reversely transcribed using random primers or oligo (dT)18 primers. Synthesized cDNA was quantified by qRT-PCR with SATB2 as a control.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
The IC50 value of CDDP was detected by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. In short, A549 and H1299 cells and CDDP-resistant A549 and H1299 cells were seeded into 96-well plate, and cells were cultured for 14 h. si-circ_0008928, in-miR-488, or HK2 was transfected into cells with their controls, and cells were continued to be cultured for 24 h. Cells were incubated with MTT solution (Beyotime) for 4 h. After that, medium was discarded, and dimethyl sulfoxide (Sigma) was used to dissolve formazan crystals. The IC50 value of CDDP was determined by measuring absorbance at 490 nm using a microplate reader (Thermo Labsystems, Waltham, MA).
Cell counting kit-8 assay
The proliferation of A549 and H1299 cells and CDDP-resistant A549 and H1299 cells was revealed by cell counting kit-8 (CCK-8) assay. In short, cells were grown in 96-well plate for 14 h. After performing cell transfection with various treatments, cells were continued to be cultivated for 0, 24, 48, and 72 h. The supernatant was removed, and cells were treated with CCK-8 reagent (Beyotime). Four hours later, cell proliferation was determined by detecting the absorbance of wavelength at 450 nm with microplate reader (Thermo Labsystems).
Transwell assay
Transwell chamber without or with Matrigel (Corning, Madison, NY) was employed to determine cell migration and invasion, respectively. In brief, cells were cultured in 24-well plate. Then, RPMI-1640 without serum (Biosun) was mixed with cells, and was then placed into the upper chamber. RPMI-1640 containing 15% FBS (Biosun) was added into the lower chamber. After 24 h, medium was discarded and cells were washed using PBS (Solarbio). After that, methanol (Beyotime) and crystal violet (Beyotime) were severally employed to mix and dye cells. Results were assessed under microscope (Olympus, Tokyo, Japan) with 100 × magnification.
Extracellular acidification rate assay
Extracellular acidification rate (ECAR) was determined by Seahorse XF Glycolysis Stress Test Kit (Agilent, Beijing, China). In short, cells were seeded into Seahorse XF microwell plate (Agilent) for 18 h. Medium was discarded, and cells were washed using PBS (Solarbio). Then, glucose (Agilent), oligomycin (Agilent), and 2-deoxyglucose (Agilent) were diluted using glutamine (Millipore, Billerica, MA), and were used to incubate with cells at 37°C in an incubator with CO2. Results were analyzed with Seahorse XF Report Generator (Agilent).
Determination of glucose consumption and lactate production
For determining glucose uptake and lactate production, glucose assay kit (Abcam, Cambridge, United Kingdom) and lactate assay kit (Abcam) were employed. In brief, cells were collected after various treatments. Then, cells were washed using cold PBS (Solarbio). The cells were suspended in glucose/lactate assay buffer (Abcam). Insoluble material was removed by centrifuging at 4°C for 2 min. After that, supernatant was collected and disposed using Deproteinizing kit (Abcam). Glucose consumption and lactate production were determined by detecting the absorbance at 570 and 450 nm, respectively, with microplate reader (Thermo Labsystems).
Western blot analysis
NSCLC cells, 16HBE, CDDP-resistant NSCLC cells, and serum exosomes were lysed using lysis buffer (Beyotime). Lysate was mixed with loading buffer (Thermo Fisher) and was boiled in boiling water for 10 min. Then, protein sample was loaded onto 12% bis-tris-acrylamide gel (Thermo Fisher) for ∼2 h. Protein bands were electrotransferred onto polyvinylidene fluoride (Millipore), which was then immersed in 5% nonfat milk (Solarbio). And membranes were incubated with anti-HK2 (1:1000; CST, Boston, MA), anticlusters of differentiation 81 (anti-CD81) (1:1000; CST), antitumor susceptibility gene 101 (anti-TSG101) (1:1000; Abcam), and anti-β-actin (1:1000; CST) overnight at 4°C, and were incubated with secondary antibody marked with horseradish peroxidase (1:5000; CST) at 37°C for 2 h. Protein bands were visualized with RapidStep ECL Reagent (Millipore). β-actin was employed as a control.
Dual-luciferase reporter assay
The binding sites between miR-488 and circ_0008928 or HK2 were predicted by starbase or microT-CDS online database. The wild-type (WT) sequences of circ_0008928 and the 3′-untranslated region (3′UTR) of HK2 were subcloned into pmirGLO vector (Promega, Madison, WI), and named circ_0008928-WT and HK2-WT. The mutant (MUT) plasmids of circ_0008928 (circ_0008928-MUT) and HK2 (HK2-MUT) were built in the same manner as shown above. Plasmids were transfected into cells with controls using DharmaFECT 4 (Thermo Fisher). Luciferase activities were determined with Dual-Lucy Assay Kit (Solarbio) with Ranilla Luciferase activity as a reference.
RNA immunoprecipitation assay
RNA immunoprecipitation (RIP) assay was performed with Magna RIP Kit (Millipore). In short, cells were collected after various treatments, and lysed using RIP lysis buffer (Millipore) containing RNase inhibitor (Millipore). Then, cell lysates were incubated with magnetic beads coated with anti-immunoglobulin G (anti-IgG; Abcam) or antiargonaute2 (anti-Ago2; Abcam) for 24 h. Lysates were incubated with proteinase K (Millipore) and RNA was purified. The contents of circ_0008928, miR-488, and HK2 enriched by anti-Ago2 or anti-IgG were analyzed by qRT-PCR.
Exosome isolation
Serum exosomes were isolated using exosome isolation kit (Umibio, Shanghai, China). In brief, serum was diluted using cold PBS buffer (Solarbio), which was then mixed with Blood PureExo Solution (Umibio). Mixture was centrifuged at 10,000 g for 60 min, and supernatant was removed. Precipitation was suspended in cold PBS (Solarbio) and centrifuged at 12,000 g for 2 min. Then, the extracted exosomes were purified using exosome purification filter (Umibio).
Transmission electron microscopy
The structure of isolated exosomes was further identified by transmission electron microscopy (TEM). In brief, exosomes were first suspended in methanol (Beyotime) and adsorbed onto a glow-discharged copper grid. After that, exosome-adsorbed copper grid was dyed using >4% uranyl acetate droplet. Finally, exosome structure was observed by TEM (FEI, Hillsboro, VA).
Data analysis
Data were obtained based on three replicates and analyzed using SPSS 21.0 software (IBM, Somers, NY), which were shown as means ± standard deviations. Significant differences were assessed by two-tailed Student's t-tests between the two groups, and by analysis of variance (ANOVA) or the Kruskal–Wallis test among three or more groups. p-Value <0.05 was considered statistically significant.
Results
Circ_0008928 expression was upregulated in A549/CDDP and H1299/CDDP cells
The expression profile of circ_0008928 was first disclosed in CDDP-resistant NSCLC cells. Results primarily showed that circ_0008928 expression was significantly upregulated in A549, H1299, H661, and SK-MES-1 cells, especially in A549 and H1299 cells, compared with control groups (Fig. 1A), and A549 and H1299 cells were chosen for further study. Subsequently, it was found that the expression of circ_0008928 was obviously increased in A549/CDDP and H1299/CDDP cells as compared with A549 and H1299 cells, respectively (Fig. 1B). It was also found that the expression level of circ_0008928 was significantly higher in cytoplasm than in nucleus (Fig. 1C), implicating that it was located in cytoplasm. Furthermore, results showed that circ_0008928 amount was lower in oligo (dT)18 primers group relative to random primers group; whereas the RNA level of linear SATB2 was not significantly changed in oligo (dT)18 primers group compared with that in random primers group (Fig. 1D), suggesting that circ_0008928 was a circRNA. These data mean that circ_0008928 might play an important role in tumor process in CDDP-resistant NSCLC.
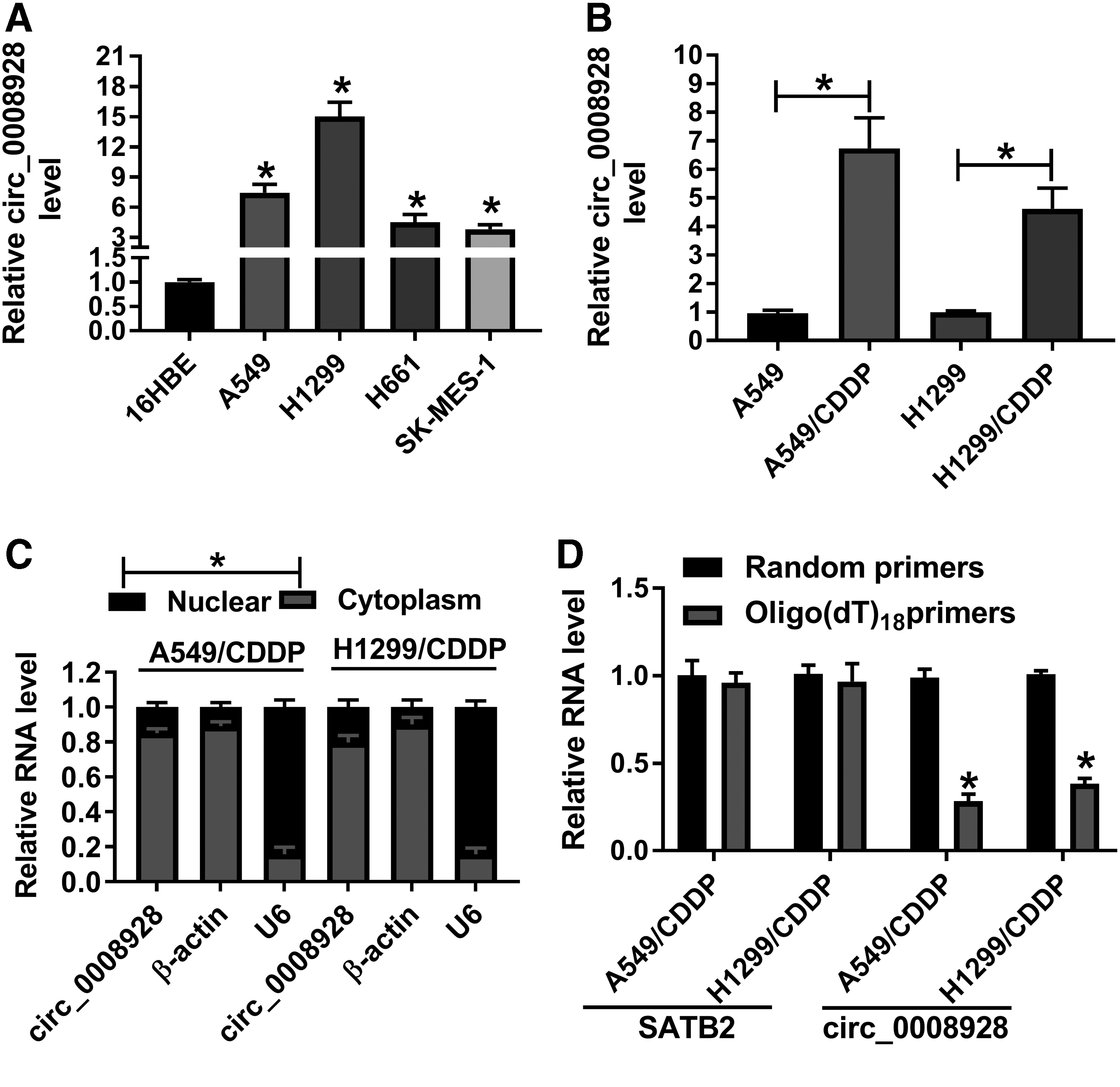
Circ_0008928 was highly expressed in A549/CDDP and H1299/CDDP cells.
Circ_0008928 knockdown improved CDDP sensitivity and attenuated CDDP-induced cell proliferation, migration, and invasion in NSCLC cells
To reveal the characteristic of circ_0008928 in CDDP-resistant NSCLC cells, the interfering plasmids of circ_0008928 were first constructed. Results showed that si-circ_0008928#1, si-circ_0008928#2, or si-circ_0008928#3 significantly decreased circ_0008928 expression, especially si-circ_0008928#1 (Fig. 2A, B). And si-circ_0008928#1 (si-circ_0008928) was employed for further study. Subsequently, results showed that the IC50 value of CDDP was significantly increased in CDDP-resistant A549 and H1299 cells, whereas this effect was hindered by circ_0008928 silencing (Fig. 2C, D). Cell proliferation was also promoted in CDDP-resistant A549 and H1299 cells; however, circ_0008928 silencing restored this effect (Fig. 2E, F). Furthermore, transwell assay illustrated that cell migration and invasion were accelerated in CDDP-resistant A549 and H1299 cells, which were impaired after circ_0008928 silencing (Fig. 2G–J). These results presented that circ_0008928 knockdown could improve CDDP sensitivity and suppress cell proliferation, migration, and invasion in CDDP-resistant A549 and H1299 cells.

Circ_0008928 silencing improved CDDP sensitivity and hindered CDDP-induced cell proliferation, migration, and invasion in A549 and H1299 cells.
Circ_0008928 silencing abolished CDDP-induced glycolysis metabolism in H1299 and A549 cells
The study continued to explore the impact of circ_0008928 on glycolysis metabolism in CDDP-resistant H1299 and A549 cells. Results showed that ECAR was significantly upregulated in A549/CDDP and H1299/CDDP, whereas circ_0008928 knockdown restored this impact (Fig. 3A, B). Meanwhile, glucose uptake and lactate production were also promoted in A549/CDDP and H1299/CDDP, which were relieved by circ_0008928 silencing (Fig. 3C–F). Furthermore, it was analyzed using Western blot that the protein expression of HK2 was obviously upregulated in A549/CDDP and H1299/CDDP; however, circ_0008928 absence blocked this effect (Fig. 3G, H). The above evidence explained that circ_0008928 could regulate CDDP-induced glycolysis metabolism in NSCLC cells.
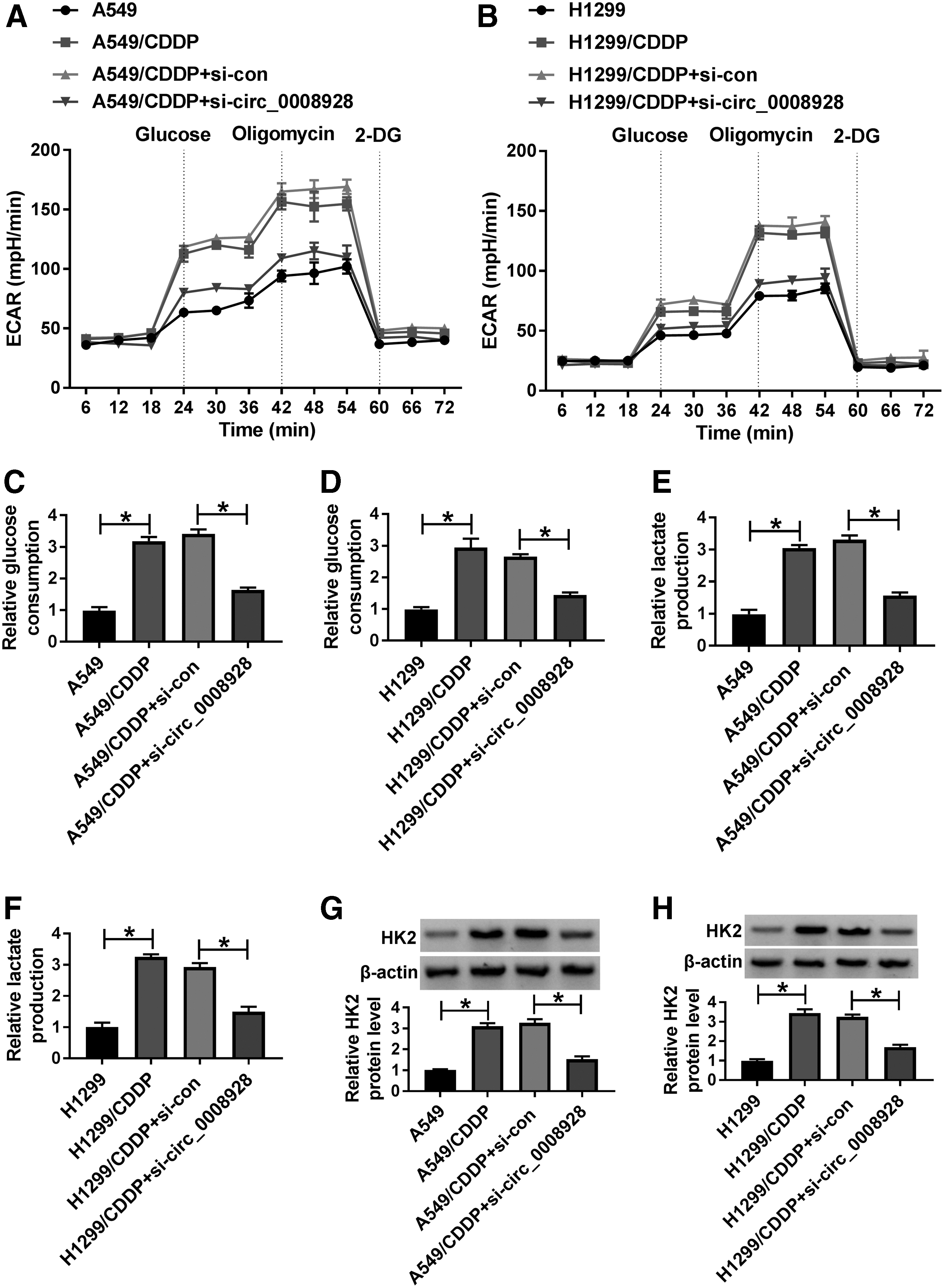
Circ_0008928 knockdown suppressed glycolysis metabolism in CDDP-resistant NSCLC cells.
Circ_0008928 was associated with miR-488 in CDDP-resistant NSCLC cells
The underneath mechanism of circ_0008928 in regulating CDDP sensitivity and biological behavior in NSCLC was continued to disclose. Results first presented that circ_0008928 contained the binding sites of nine miRNAs after taking intersection of the results predicted by circbank, starbase, and circRNA interactome online database (Fig. 4A). To further seek miRNAs associated with circ_0008928, the transfection efficiency of miR-488 mimic was detected. Results showed that miR-488 mimic significantly upregulated miR-488 expression in A549/CDDP and H1299/CDDP (Fig. 4B). Dual-luciferase reporter assay further illustrated that only miR-488 and circ_0008928 cotransfection significantly repressed luciferase activity (Fig. 4C), suggesting that miR-488 might be associated with circ_0008928. The relative luciferase activity was also detected after cotransfection between circ_0008928-MUT and miR-488 or miR-con, and data exhibited that the relative luciferase activity had no obvious change (Fig. 4D). And the binding sites between miR-488 and circ_0008928 are shown in Figure 4E. Meanwhile, RIP assay illustrated that the amount of circ_0008928 and miR-488 was apparently enriched in anti-Ago2 groups compared with that in anti-IgG groups (Fig. 4F, G). In addition, qRT-PCR results investigated that miR-488 expression was obviously decreased in A549, H1299, H661, and SK-MES-1 cells as compared with 16HBE cells (Fig. 4H) and was also strikingly downregulated in CDDP-resistant A549 and H1299 relative to A549 and H1299, respectively (Fig. 4I). Furthermore, the effects of circ_0008928 overexpression or knockdown on miR-488 expression were disclosed. Results initially demonstrated that circ_0008928 expression was notably upregulated in both A549/CDDP and H1299/CDDP cells transfected with circ_0008928 (Fig. 4J). It was then found that the expression of miR-488 was effectively downregulated by circ_0008928 overexpression, and was obviously upregulated after circ_0008928 knockdown in A549/CDDP and H1299/CDDP cells (Fig. 4K, L). The above data implicated that miR-488 bound to circ_0008928 in CDDP-resistant NSCLC cells.
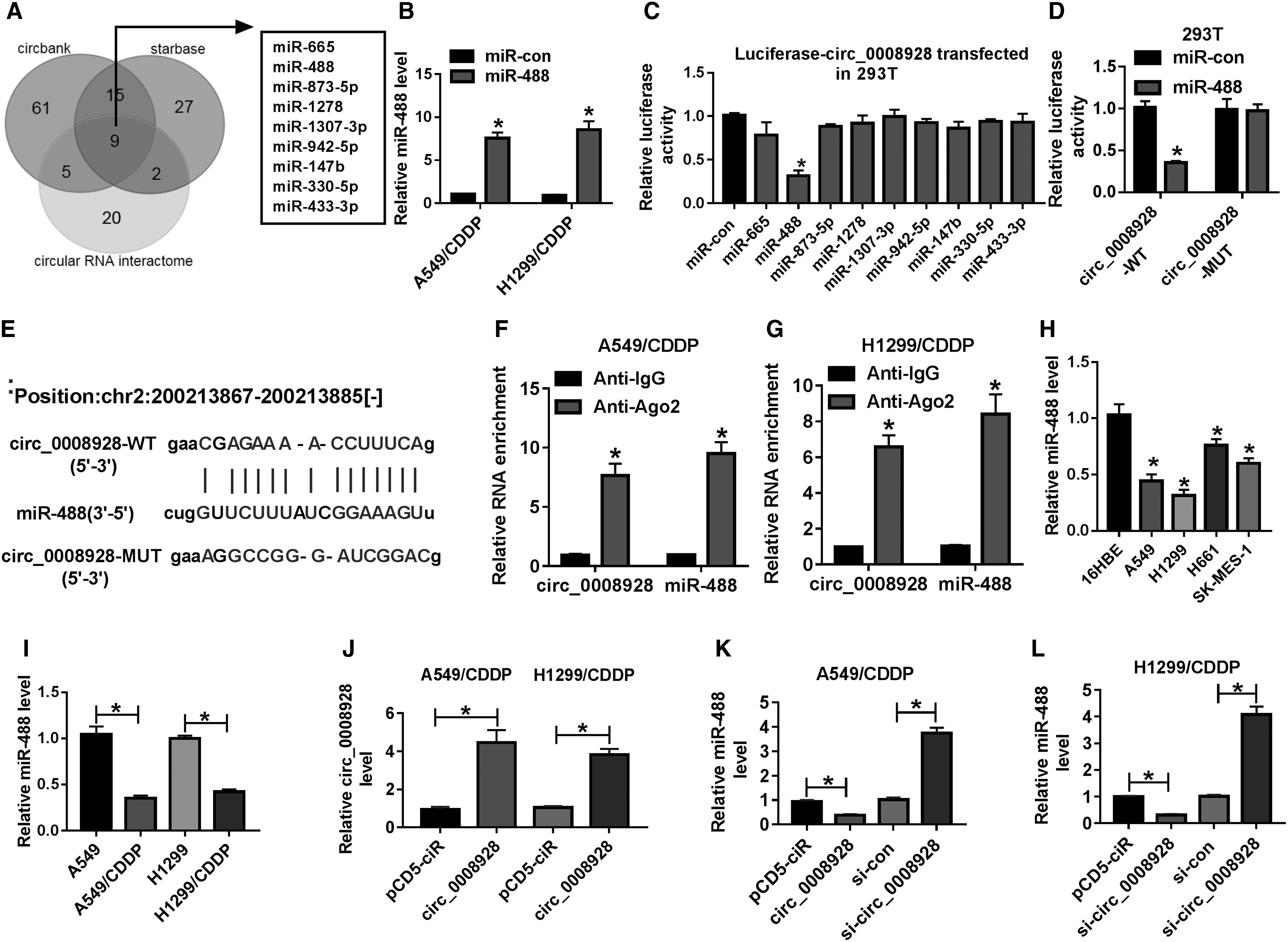
Circ_0008928 acted as a sponge of miR-488 in CDDP-resistant NSCLC cells.
MiR-488 interacted with HK2 in CDDP-resistant NSCLC cells
The associated mRNA with miR-488 was continued to be explored. Results showed that HK2 3′UTR (chr2:75119262–75119280) possessed the binding sequence of miR-488 (Fig. 5A). Dual-luciferase reporter assay investigated that luciferase activity in miR-488 and HK2-WT group was significantly inhibited, while there was no dramatic change after miR-488 and HK2-MUT cotransfection (Fig. 5B). RIP assay also demonstrated that both HK2 and miR-488 were significantly enriched in anti-Ago2 groups compared with that in anti-IgG group (Fig. 5C, D). In addition, it was found that the expression level of HK2 was apparently upregulated in A549, H1299, H661, and SK-MES-1 cells and CDDP-resistant A549 and H1299 cells as compared with control groups, respectively (Fig. 5E, F). To reveal the effects of miR-488 mimic or inhibitor on HK2 expression, the efficiency of miR-488 inhibitor in knocking down miR-488 expression was detected. Results showed that miR-488 expression was obviously decreased after miR-488 inhibitor transfection in CDDP-resistant A549 and H1299 cells (Fig. 5G). And results illustrated that HK2 protein expression was apparently downregulated by miR-488 mimic, and was notably upregulated by miR-488 inhibitor in CDDP-resistant A549 and H1299 cells (Fig. 5H, I). Therefore, this evidence demonstrated that miR-488 was associated with HK2 in CDDP-resistant NSCLC cells.
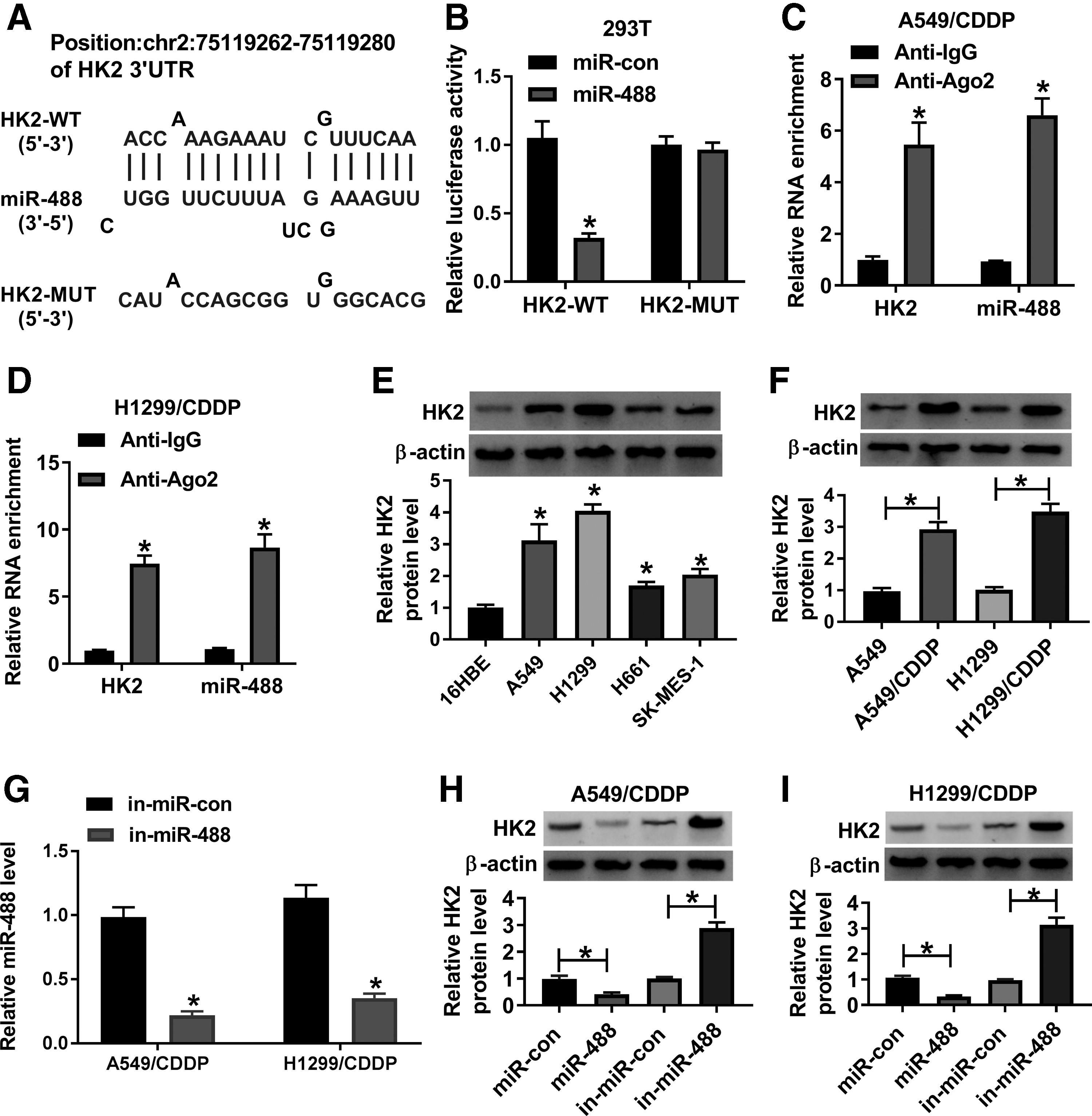
MiR-488 bound to HK2 in CDDP-resistant NSCLC cells.
Circ_0008928 knockdown improved CDDP sensitivity and inhibited tumor progression and glycolysis metabolism by downregulating HK2 expression through sponging miR-488 in CDDP-resistant NSCLC cells
Given that circ_0008928 was a sponge of miR-488 and miR-488 bound to HK2, whether circ_0008928 regulated CDDP sensitivity and biological behavior of CDDP-resistant NSCLC cells by mediating HK2 expression through sponging miR-488 was further unveiled. Our results first showed that HK2 protein expression was significantly downregulated by circ_0008928 silencing, whereas this effect was abolished by miR-488 inhibitor or HK2 overexpression (Fig. 6A, B). Subsequently, the IC50 value of CDDP was reduced by circ_0008928 knockdown, but this impact was restored after miR-488 inhibitor or HK2 transfection (Fig. 6C, D). MiR-488 inhibitor or ectopic HK2 expression also attenuated the inhibitory effect of circ_0008928 absence on cell proliferation in CDDP-resistant A549 and H1299 cells (Fig. 6E, F). Transwell assay demonstrated that the migration and invasion of CDDP-resistant A549 and H1299 cells were inhibited by circ_0008928 knockdown; however, miR-488 inhibitor or HK2 overexpression impaired these impacts (Fig. 6G–J). In addition, miR-488 inhibitor or HK2 overexpression restrained the inhibitory effect of circ_0008928 repression on ECAR (Fig. 7A, B). The glucose consumption and lactate production of CDDP-resistant A549 and H1299 cells were repressed by circ_0008928 knockdown, which were blocked by miR-488 inhibitor or HK2 overexpression (Fig. 7C–F). Thus, these data investigated that circ_0008928 could regulate CDDP resistance, cell progression, and glycolysis metabolism by downregulating HK2 expression through sponging miR-488 in CDDP-resistant NSCLC cells.
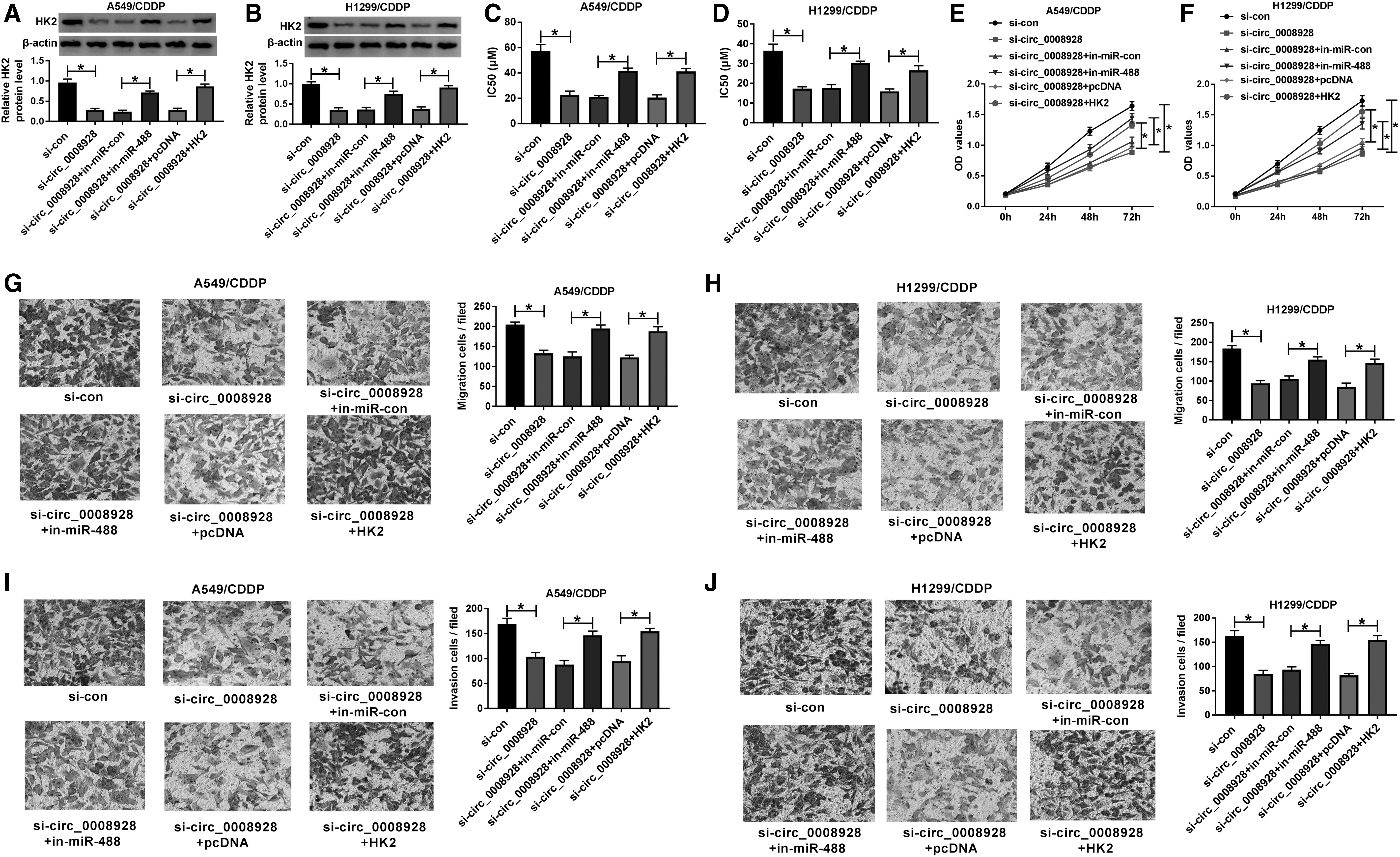
Circ_0008928 regulated CDDP resistance, NSCLC progression, and glycolysis metabolism by sponging miR-488 or modulating HK2 expression in CDDP-resistant NSCLC.
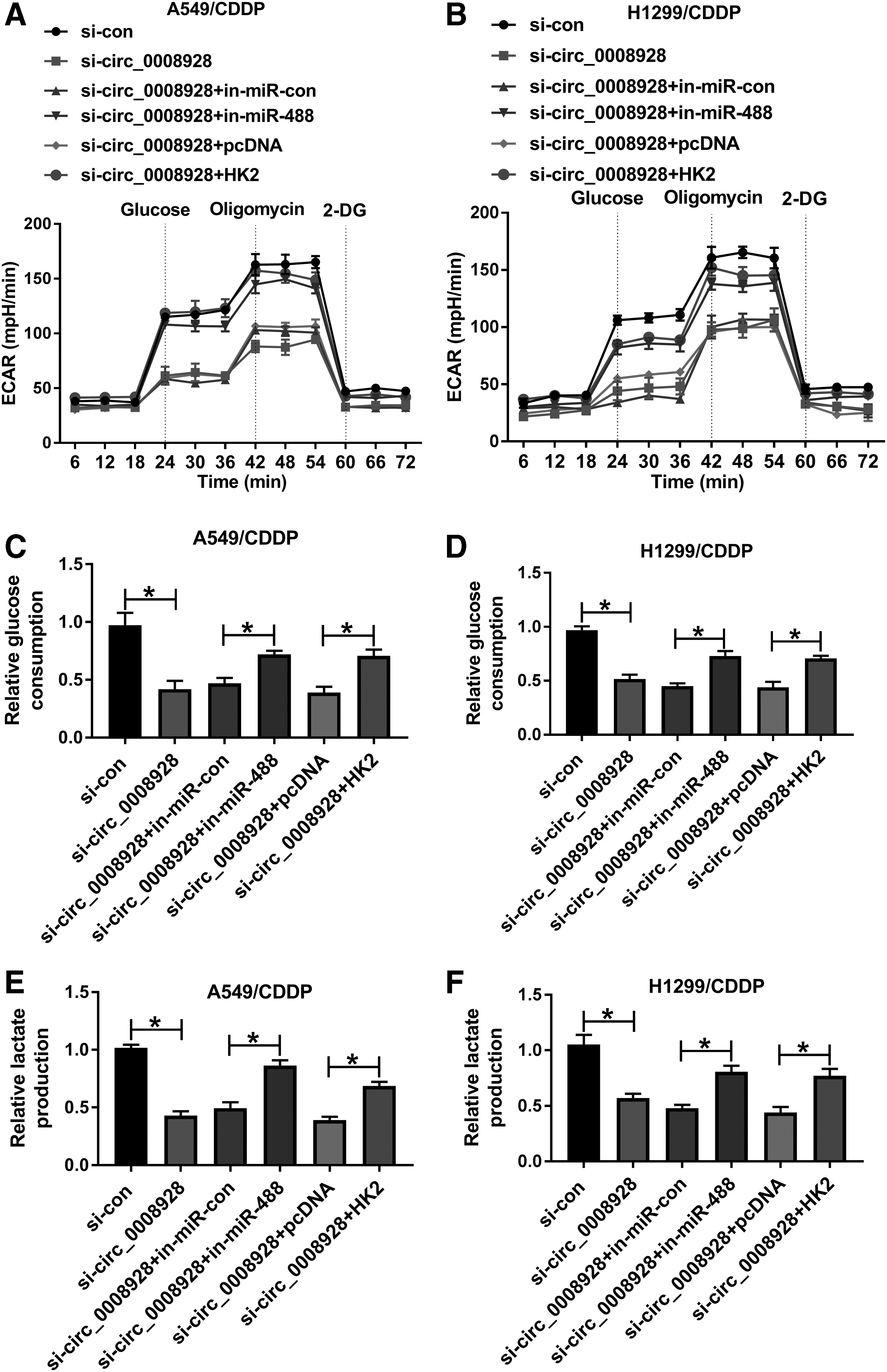
Circ_0008928 regulated glycolysis metabolism by sponging miR-488 or modulating HK2 expression in CDDP-resistant NSCLC cells.
Circ_0008928 expression was significantly upregulated in serum exosomes of CDDP-resistant NSCLC patients
Exosomes, a kind of extracellular vesicles, can be produced by various types of cells, including cancer cells. 24 Exosomes play vital parts in transferring or exchanging RNAs, proteins as well as lipids, and are of great concern in developing new therapeutic targets for cancers. 25 In this article, to determine whether circ_0008928 expression was affected in serum exosomes of CDDP-resistant NSCLC, exosomes were collected from 28 CDDP-sensitive NSCLC patients, 19 CDDP-resistant NSCLC patients, and 47 volunteers without NSCLC. Results first showed that exosomes were successfully extracted as extracted particles had a similar structure to exosomes (Fig. 8A) and exosome-related proteins (CD81 and TSG101) were detected (Fig. 8B). Subsequently, qRT-PCR results showed that circ_0008928 expression was significantly upregulated in nonresponse group compared with control group (Fig. 8C). These data showed that CDDP sensitivity affected circ_0008928 expression in serum exosomes.
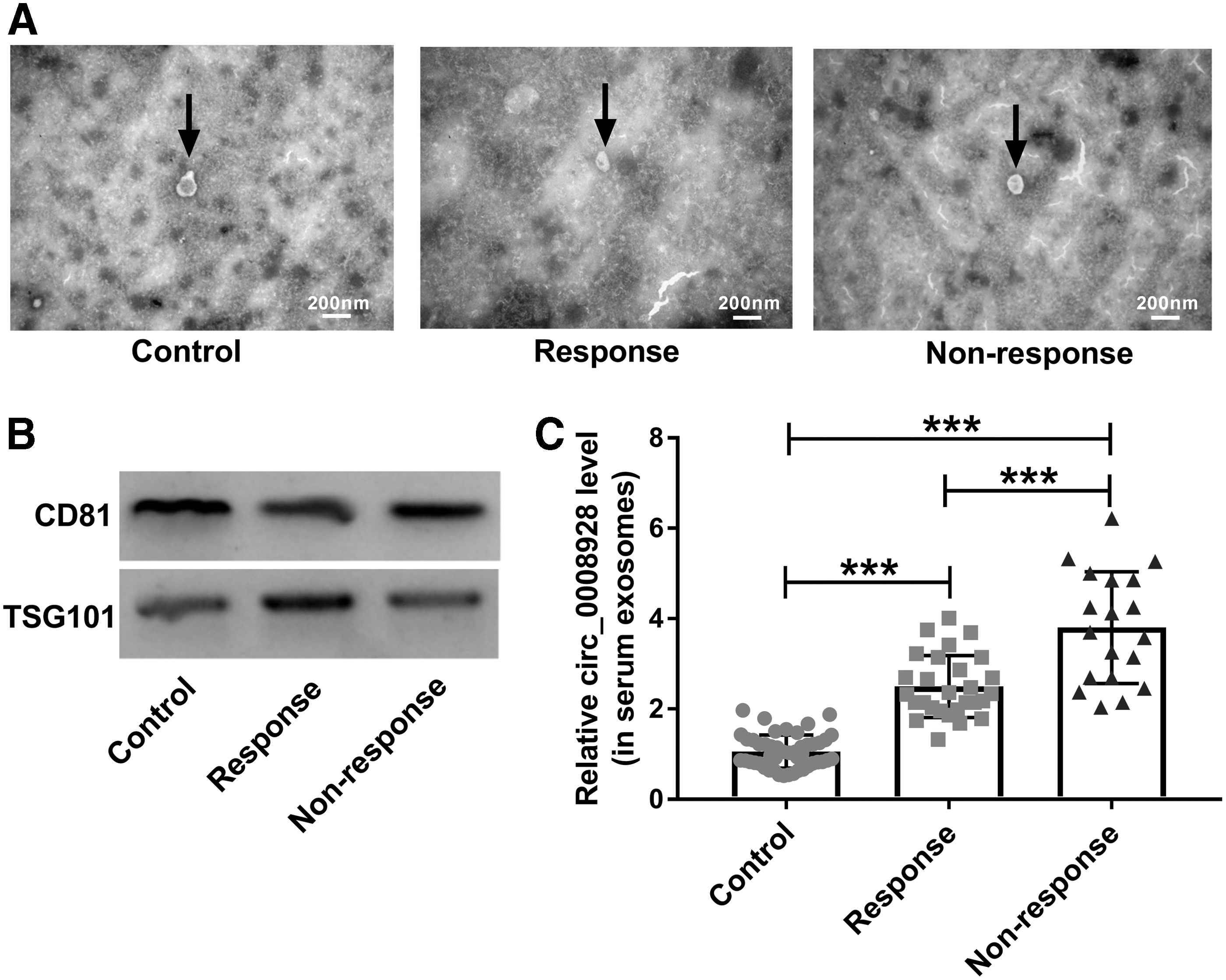
Circ_0008928 expression was upregulated in serum exosomes of CDDP-resistant NSCLC patients.
Discussion
The expression of circRNA has been revealed to be dysregulated in NSCLC cases. 26 And circRNA has been considered as a biomarker during NSCLC therapy. 27 Previous studies explained that circRNA facilitated cell proliferation and metastasis, and suppressed cell apoptosis in NSCLC patients. 28,29 CDDP is always used to treat advanced NSCLC patients; however, CDDP resistance brings many obstacles to the treatment of NSCLC patients. Nevertheless, the mechanism of NSCLC resistance to CDDP has not been completely unveiled. In this study, the regulatory mechanism of circ_0008928 in NSCLC sensitivity to CDDP was disclosed.
CircRNAs are a kind of vital regulators in NSCLC progression. It was indicated that circ_0106705 knockdown suppressed cell proliferative and migratory abilities and glycolysis production in NSCLC. 30 Downregulation of circ_0087862 expression was found to weaken cell proliferation and metastasis, and induce cell apoptosis in NSCLC. 31 Zhang et al. also explained that circ_0008928 (named circSATB2) facilitated cell proliferation, migration, and invasion in NSCLC. 32 CDDP resistance has been found to be regulated by circRNAs, such as circ_0072083, 11 circ_0007385, 33 and circ_0003418. 34 However, there were no relevant data on CDDP sensitivity regulated by circ_0008928 in NSCLC. And in this study, circ_0008928 was disclosed to participate in regulating CDDP resistance in NSCLC for the first time. Our results showed that circ_0008928 expression was significantly upregulated in CDDP-resistant NSCLC cells, and circ_0008928 silencing improved CDDP sensitivity and repressed cell proliferation, migration, and invasion in CDDP-resistant NSCLC cells. Furthermore, circ_0008928 knockdown inhibited cell glycolysis metabolism by detecting ECAR, glucose, lactate, and HK2 protein expression in CDDP-resistant NSCLC cells. An increasing number of studies presented that exosomes could transmit bioactive cargoes, including circRNAs. 35 And, circ_0008928 was indicated to be transmitted by exosomes. 32 Thus, to illustrate whether circ_0008928 expression was affected in exosomes of CDDP-resistant NSCLC patients, serum exosomes of CDDP-NSCLC patients and volunteers without NSCLC were collected. Results demonstrated that circ_0008928 expression was obviously upregulated in serum exosomes of CDDP-resistant NSCLC patients relative to serum exosomes of CDDP-sensitive NSCLC patients or volunteers. These data unveiled that exosomes-based circ_0008928 acted as an oncogene and inhibited CDDP sensitivity in NSCLC.
Data indicate that a defined function of circRNA is to mediate protein expression through sponging miRNA. 36 Based on that, the associated miRNAs with circ_0008928 were predicted and identified by online database and dual-luciferase reporter and RIP assays, respectively. Results displayed that circ_0008928 was associated with miR-488. According to present data, miR-488 served as a cancer suppressor in NSCLC progression. For instance, circ_0020123 silencing repressed cell growth, migration, and invasion, and promoted cell apoptosis by sponging miR-488, 37 implicating that miR-488 hindered NSCLC progression. In another example, Fang et al. reported that miR-488 suppressed cell proliferation and metastasis in NSCLC. 18 In this study, it was found that the inhibitor of miR-488 attenuated the inhibitory effects of circ_0008928 knockdown on cell proliferation, migration, and invasion in A549/CDDP and H1299/CDDP cells, which showed that miR-488 inhibited NSCLC development. Beyond that, miR-488 was found to improve CDDP sensitivity and repress glycolysis metabolism in CDDP-resistant NSCLC cells.
Glycolysis offers huge energy to sustain cell proliferation in cancers, and HK2 plays a dominant role in glycolysis. 38 HK2 has been unveiled to participate in the progression of gallbladder cancer and breast cancer. 39,40 He and Liu displayed that HK2 participated in regulating CDDP resistance in NSCLC. 41 Based on the main characteristic of HK2 in cancer progression, the target gene of miR-488 was predicted. As we expected, results showed that miR-488 was associated with HK2. Thus, we hypothesized that circ_0008928 might regulate CDDP sensitivity by upregulating HK2 expression. To test that, the effects between circ_0008928 knockdown and HK2 overexpression on CDDP sensitivity, tumor progression, and glycolysis metabolism were illustrated. Our results displayed that ectopic HK2 expression hindered the effects of circ_0008928 silencing on CDDP sensitivity, tumor progression, and glycolysis metabolism.
All in all, the expression of circ_0008928 and HK2 was upregulated, and miR-488 expression was downregulated in CDDP-resistant NSCLC cells. Circ_0008928 knockdown suppressed cell proliferation, migration, invasion, and glycolysis metabolism, and improved CDDP sensitivity in CDDP-resistant NSCLC. In addition, miR-488 inhibitor or HK2 overexpression attenuated the effects of circ_0008928 knockdown on CDDP sensitivity, tumor progression, and glycolysis metabolism in CDDP-resistant NSCLC. Furthermore, circ_0008928 expression was demonstrated to be increased in serum exosomes of CDDP-resistant NSCLC patients. Our findings illustrate that circ_0008928 can be employed as a potential biomarker for the diagnosis of NSCLC. In addition, our result provides a theoretical basis for studying circRNA-mediated therapy in NSCLC with CDDP.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by technology innovation ability cultivation project of Chenzhou Science and Technology Bureau Science (zdyf201950) and Hospital Project of Chenzhou First People's Hospital (N2020-42).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.