
Abstract
Background:
Circular (Circ)RNA circFAT1 play tumor-suppressive or oncogenic roles in different cancers. Microarray analysis observed altered expression of circFAT1 in endometrial cancer (EC) and its inverse correlation with miR-21.
Materials and Methods:
Expression of circFAT1 and miR-21 in EC and paired nontumor tissues collected from 62 EC patients was analyzed by quantitative reverse transcription PCR (RT-qPCR). An experiment was conducted to evaluate the expression and interaction between circFAT1 and miR-21, followed by RT-qPCR and methylation-specific PCR. The role of circFAT1 and miR-21 in regulating the stemness was assessed by cell stemness assay. Heml 1.0 software was used to show differential gene expression. ANOVA Tukey's test and Pearson's correlation coefficient was used.
Results:
CircFAT1 was upregulated in EC and positively correlated with miR-21 across EC tissues. In RL95-2 and HEC-1-A cells, overexpression of circFAT1 increased the expression levels of miR-21 and decreased the methylation of miR-21 gene, whereas overexpression of miR-21 did not alter the expression of circFAT1. Cell stemness analysis showed that overexpression of circFAT1 and miR-21 increased cell stemness, and miR-21 inhibition decreased cell stemness. Moreover, inhibitor of miR-21 suppressed the role of circFAT1.
Conclusions:
In conclusion, circFAT1 is upregulated in EC and it may increase cancer cell stemness by upregulating miR-21.
Introduction
As a malignancy originates from the endometrium (lining) of the uterus, endometrial cancer (EC) has been recognized as the most common type of gynecological cancer in developed countries. 1 In developing countries, such as China, EC is also responsible for a considerable portion of new cancer cases and cancer deaths. 2 It is estimated that China accounts for about 11.7% of all new cancer cases. 3 With the development of cancer therapies, such as chemotherapy, surgery, and radiation therapy, the overall 5-year survival rate of EC patients with localized tumors has reached 95%. 4 However, there is still no cure for metastatic EC. Once distant tumor metastasis has occurred, only <20% patients can survive 5 years. 5,6 Therefore, novel approaches are needed to treat EC.
Molecular targeted therapies are emerging novel anticancer approaches to treat cancers by regulating the expression of cancer-related genes. 7 With the increased understanding of the molecular pathogenesis of EC, some molecular players, such as the PI3K/Akt/mTOR pathway, have been recognized as potential targets for EC targeted therapy. 8 –10 Circular RNAs (circRNAs) are recognized as long noncoding RNAs (lncRNAs) and they regulate cancer development and progression by regulating gene expression at transcriptional and translational levels, 11,12 suggesting the potential of circRNAs for targeted therapies of cancer. 13 However, the functions of most lncRNAs in cancer biology are unknown. It has been reported that circFAT1 may play tumor-suppressive or oncogenic roles in different cancers, 14,15 whereas its role in EC is unknown. Our preliminary sequencing analysis revealed altered expression of circFAT1 in EC and the inverse correlation between circFAT1 and miR-21 (data not shown). MiR-21 is a critical player in cancer biology. 16 In this study, we explored the crosstalk between circFAT1 and miR-21 in EC.
Materials and Methods
EC patients
A total of 62 female patients (60.3 ± 4.9 years old) diagnosed as EC through a histopathological analysis at Regional Specialized Disease Center of Northern Zhejiang, Jiaxing Maternal and Child Health Hospital Affiliated to Jiaxing University between May 2017 and August 2020 were enrolled in this study. The Ethics Committee of this hospital approved this study. The 62 patients were excluded from other clinical disorders, initiated therapy, previous history of malignancies, and mental disorders. Only newly diagnosed EC cases were enrolled. The 62 patients were classified into AJCC stage I or II (n = 22) and III or IV (n = 40). All patients provided a written form of informed consent. The clinical data of patients are listed in Table 1.
Clinical Data of Patients
BMI, body mass index.
EC tissues and cells
EC and adjacent (within 3 cm around tumors) tissue samples were collected through fine needle aspiration from each patient. All tissue samples were confirmed by histopathological analysis. Tissue samples were kept in liquid nitrogen before the subsequent assays. Two human EC cell liens RL95-2 and HEC-1-A (ATCC) were used as the cell model of EC. Cells were cultivated in a medium composed of 10% fetal bovine serum and 90% Dulbecco's modified Eagle's medium. Before the subsequent experiments, cells were cultivated at 37°C in a 5% CO2 incubator with 95% humidity to reach about 85% confluence.
Transient transfections
RL95-2 and HEC-1-A cells (108) were transfected with circFAT1 expression vector (1 μg) or miR-21 mimic (40 nM) to overexpress circFAT1 and miR-21. Cells were also transfected with miR-21 inhibitor (40 nM) to silence miR-21. Backbone expression vector or circFAT1 was constructed with pcDNA3.1 vector (Invitrogen). MiR-21 mimic and negative control (NC) miRNA as well as miR-21 inhibitor and NC inhibitor were purchased from Sangon Biotech (Shanghai, China). Control (C) cells were untransfected cells. NC cells were cells transfected with empty vector, NC miRNA, or NC inhibitor. Before the subsequent assays, transfected cells were cultivated in fresh medium for an additional 48 h.
Preparation of RNA samples
Total RNAs were extracted from tissues and cells by using Ribozol (Invitrogen). RNA samples were incubated with DNase I (Invitrogen) at 37°C to digest genomic DNA until an OD value close to 2.0 (pure RNA samples) was reached. RNA integrity was analyzed by urea-PAGE (polyacrylamide gel electrophoresis) gel (5%) electrophoresis.
Quantitative reverse transcription PCRs
SS-IV-RT system (Invitrogen) was used to reverse transcribe RNA samples into cDNA samples. To determine the expression of circFAT1, quantitative reverse transcription PCR (RT-qPCR) were carried out by using Bio-Rad cDNA Supermix Kit (Bio-Rad) with 18S rRNA as an internal control.
The expression of mature miR-21 was analyzed by using All-in-One™ miRNA qRT-PCR Detection Kit (Genecopoeia) by adding poly (A), followed by miRNA reverse transcriptions and qPCRs. Ct values of target genes were normalized to endogenous controls by using the 2−ΔΔCT method. The sample with the biggest ΔCT was set to value “1.” All other samples were normalized to this sample to calculate the relative expression levels of genes.
Methylation-specific PCR
Genomic DNA was extracted from both RL95-2 and HEC-1-A cells at 48 h post-transfection. DNA samples were converted by using EZ DNA Methylation Kit (D5001; ZYMO Research). RT-PCRs and methylation-specific PCRs (MSPs) were performed by using Taq 2X Master Mix (NEB) to amplify methylated- and unmethylated-miR-21 gene using different primer pairs. PCR products were subjected to 1% agarose gel electrophoresis, followed by ethidium bromide staining.
Cell stemness analysis
RL95-2 and HEC-1-A cells with transfections were collected and washed with precold phosphate-buffered saline. Cells from each transfection group (105 cells) were then subjected to staining with IgG1-PE or CD133-PE antibody (Meltenyi Biotec) at 4°C for 20 min. Finally, ACS Aria system (BD Immunocytometry Systems) was used to detect the staining signals. Cell stemness was reflected by the percentage of CD133+ cells.
Statistical analyses
Differential expression of circFAT1 and miR-21 in EC and paired nontumor tissues were expressed as heatmaps plotted by using Heml 1.0 software. Comparisons among multiple transfection groups were analyzed by ANOVA Tukey's test. Correlations were explored by Pearson's correlation coefficient. p < 0.05 was considered statistically significant.
Results
Altered expression of circFAT1 and miR-21 in EC tissues
EC and paired nontumor tissues collected from the 62 EC patients were subjected to RT-qPCRs to analyze the differential expression of circFAT1 and miR-21 in EC. Heatmaps were plotted by using Heml 1.0 software. In the heatmaps, cold and warm colors represented low and high expression levels of the gene, respectively. The results illustrated that, compared with nontumor tissues, EC tissues exhibited increased expression levels of circFAT1 (Fig. 1A) and miR-21 (Fig. 1B).

EC tissues exhibited altered expression of circFAT1 and miR-21. Paired EC and nontumor tissues collected from the 62 EC tissues were subjected to RNA isolation and RT-qPCRs to analyze the differential expression of circFAT1
Positive correlation between circFAT1 and miR-21 across EC tissues
The correlations between circFAT1 and miR-21 across both EC and nontumor tissues were explored by Pearson's correlation coefficient. The data used in the correlation analysis were mean values of gene expression levels in three technical qPCR replications. It was observed that the expression of circFAT1 and miR-21 was significantly and positively correlated across EC tissues (Fig. 2A; R 2 = 0.8009; p < 0.0001). No significant correlation between them was observed across nontumor tissues (Fig. 2B; R 2 = 7.375e-007; p = 0.9947).
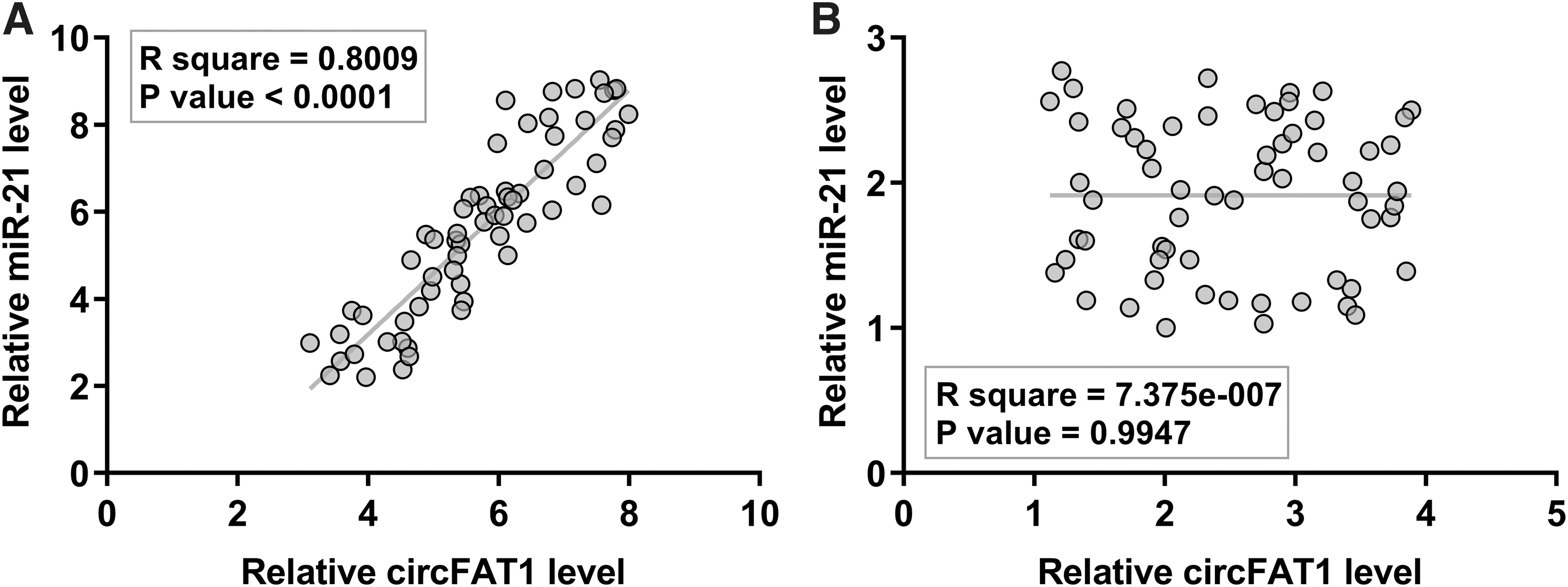
A positive correlation between circFAT1 and miR-21 was observed across EC tissues. The correlations between circFAT1 and miR-21 across both EC
Overexpression of circFAT1 increased the expression levels of miR-21 in both RL95-2 and HEC-1-A cells
To explore the interaction between circFAT1 and miR-21 in EC, RL95-2 and HEC-1-A cells were transfected with either circFAT1 expression vector or miR-21 mimic, and the overexpression of circFAT1 and miR-21 was analyzed at 24 and 48 h post-transfection. It was observed that circFAT1 and miR-21 were significantly overexpressed in both cell lines at the two time points. In both cell lines, a two-fold increase was observed at 24 h and a four-fold increase was observed at 48 h (p < 0.05) (Fig. 3A). Moreover, overexpression of circFAT1 significantly increased the expression levels of miR-21. A 2-fold increase was observed at 24 h, and a >3-fold increase was observed at 48 h (p < 0.05) (Fig. 3B). In contrast, overexpression of miR-21 did not alter the expression of circFAT1 (Fig. 3C).
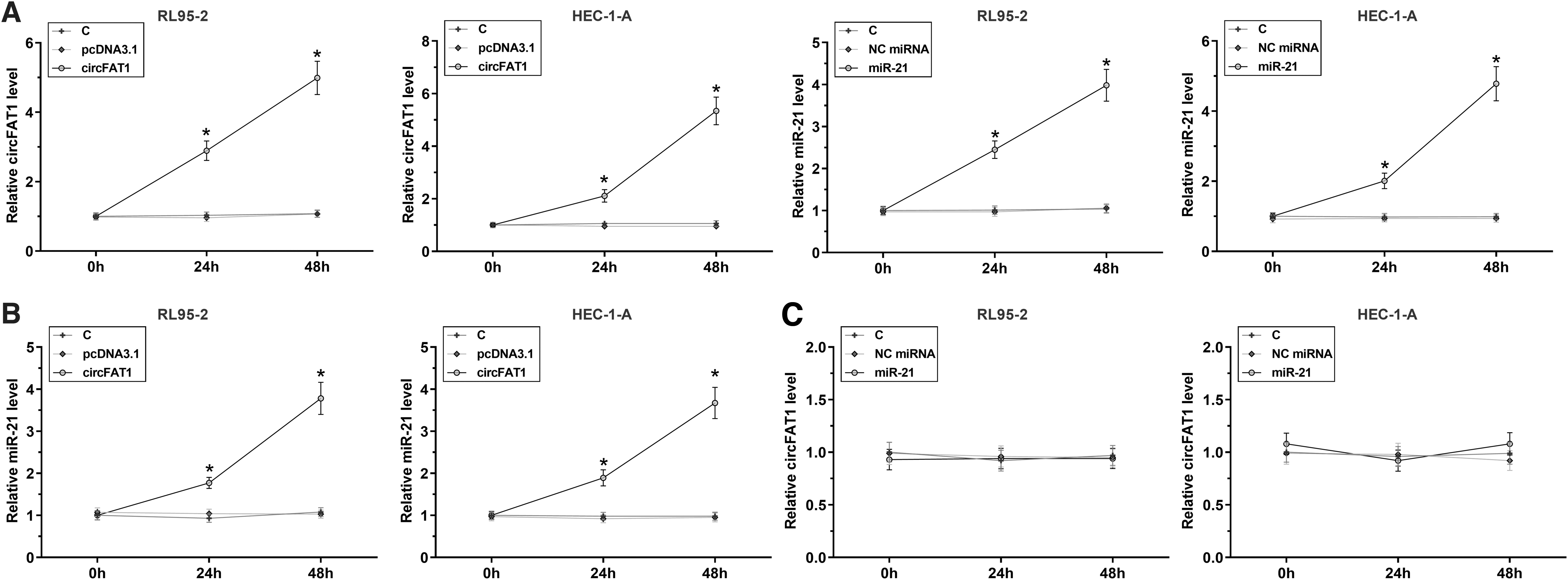
Overexpression of circFAT1 increased the expression levels of miR-21 in both RL95-2 and HEC-1-A cells. To explore the interaction between circFAT1 and miR-21 in EC, RL95-2 and HEC-1-A cells were transfected with either circFAT1 expression vector or miR-21 mimic, and the overexpression of circFAT1 and miR-21 was analyzed at 24 and 48 h post-transfection
Overexpression of circFAT1 decreased the methylation of miR-21 gene in both RL95-2 and HEC-1-A cells
The MSP was performed to analyze the effects of overexpression of circFAT1 on the methylation of miR-21 gene. RL95-2 and HEC-1-A cells were transfected with either circFAT1 expression vector or empty vector. The MSPs and PCRs were performed to amplify methylated- and unmethylated-miR-21 gene. PCR products were subjected to 1% agarose gel electrophoresis, followed by ethidium bromide staining. Compared with cells transfected with empty vector, cells transfected with circFAT1 expression vector exhibited decreased methylation of miR-21 gene (Fig. 4).

Overexpression of circFAT1 decreased the methylation of miR-21 gene in both RL95-2 and HEC-1-A cells. The MSP was performed to analyze the effects of circFAT1 overexpression on the methylation of miR-21 gene. RL95-2 and HEC-1-A cells were transfected with either circFAT1 expression vector or empty vector. The MSP and routine PCRs were performed to analyze the methylation of miR-21 gene. M, methylation; MSP, methylation-specific PCR; PCR, polymerase chain reaction; U, un-methylation.
Overexpression of circFAT1 increased the stemness of RL95-2 and HEC-1-A cells through miR-21
Cell stemness assay was performed to investigate the role of circFAT1 and miR-21 in regulating the stemness of both RL95-2 and HEC-1-A cells. The percentage of CD133+ cells represents cell stemness. The results showed that overexpression of circFAT1 and miR-21 increased cell stemness, and inhibitor of miR-21 decreased cell stemness. Moreover, inhibitor of miR-21 reduced the effects of overexpression of circFAT1 on cell stemness (p < 0.05) (Fig. 5).
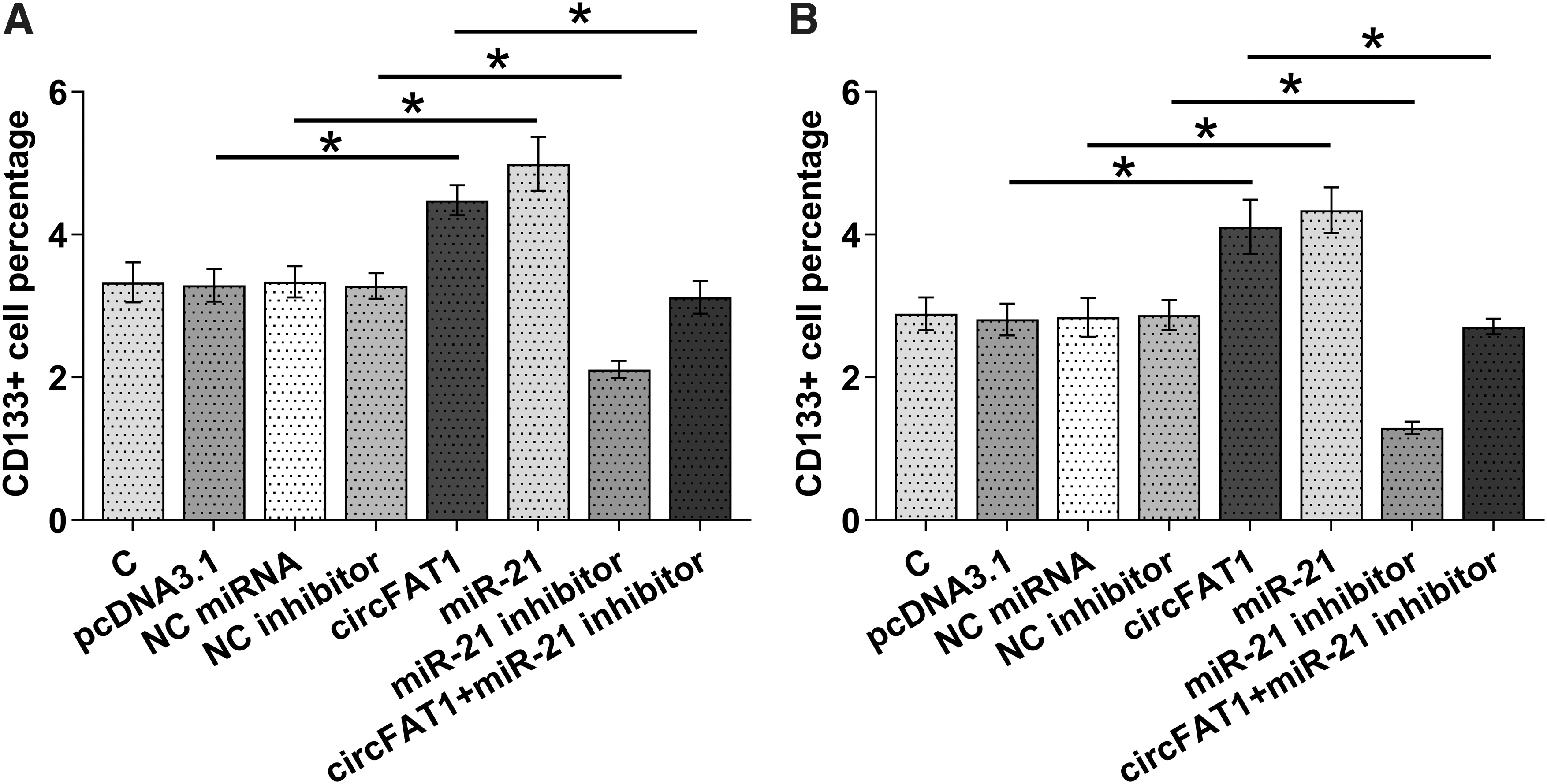
Overexpression of circFAT1 increased the stemness of RL95-2 and HEC-1-A cells through miR-21. Cell stemness assay was performed to analyze the role of circFAT1 and miR-21 in regulating the stemness of both RL95-2
Discussion
This study explored the crosstalk between circFAT1 and miR-21 in EC. We found that circFAT1 was significantly upregulated in EC and it may upregulate the expression of miR-21 through methylation, thereby increasing cancer cell stemness.
It has been reported that circFAT1 may play different roles in different types of cancer. 14,15 For instance, circFAT1 is downregulated in gastric cancer and it may targets miR-548a in cytoplasm to upregulate YBX1, thereby suppressing cancer progression. 14 In contrast, circFAT1 is upregulated in osteosarcoma and sponge miR-181b to upregulate HK2, thereby promoting cancer progression. 15 In this study, we found that circFAT1 was upregulated in EC. In addition, overexpression of circFAT1 significantly increased EC cell stemness, which is closely correlated with the poor survival of most cancers. 17 Therefore, our study proved that circFAT1 in an oncogenic circRNA in EC.
MiR-21 is a well-established oncogenic miRNA in different types of cancer, including EC. 18,19 MiR-21 is upregulated in EC and targets multiple tumor suppressor genes, such as PTEN and SRY‑box 17, to increase cancer cell proliferation and invasion as well as epithelial-to-mesenchymal transition. 18,19 However, the role of miR-21 in EC cell stemness is unknown. In this study, we observed upregulation of miR-21 in EC. In addition, overexpression of miR-21 significantly increased the stemness of RL95-2 and HEC-1-A cells. Therefore, miR-21 may also promote EC progression by increasing cell stemness.
In this study, we showed that miR-21 did not affect the expression of circFAT1. Therefore, circFAT1 is unlikely a target of miR-21 in EC. Previous studies have shown that circFAT1 participates in cancer biology mainly by sponging miRNAs. 14,15 In this study, we showed that circFAT1 may also regulate the expression of miRNAs through methylation. Therefore, the present study increased the understanding of the role of circFAT1 in cancer biology. Interestingly, circFAT1 and miR-21 were only significantly correlated across EC tissues, but not nontumor tissues. Therefore, the interaction between them in RL95-2 and HEC-1-A cells may be mediated by certain EC-related methylation factors.
This study reported the role of circFAT1 in regulating the methylation of miR-21, which is a key player in cancer biology. 16 However, only cell lines were used and the interactions between them remain to be studied in vivo. Future studies may also focus on the clinical application of circFAT1 in the diagnosis and treatment of EC. In addition, the interactions between circFAT1 and miR-21 in other types of cancer are unknown. More studies are needed.
In conclusion, circFAT1 is upregulated in EC. In addition, circFAT1 may upregulate miR-21 through methylation to increase EC cell stemness.
Footnotes
Authors' Contributions
W.W., W.Z.: study concepts, literature research, clinical studies, data analysis, experimental studies, and article writing and review; J.Z.: study design, literature research, experimental studies, and article editing; Y.W.: definition of intellectual content, clinical studies, data acquisition, and statistical analysis; X.T.: data acquisition, article preparation, and data analysis.
Declarations
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Fund Project of Jiaxing Science and Technology Bureau (Grant No. 2017BY18029, 2018AD32105).