
Abstract
Background:
Apoptosis plays an essential role in the development and treatment of tumors, and caspase-8 (CASP8) plays an important role in the enzyme cascade reaction that leads to apoptosis. Human epidermal growth factor receptor 2 (HER-2) overexpressing breast cancer is highly aggressive and has a high recurrence rate and poor prognosis. This study investigated whether lentivirus-mediated Gag-CASP8 can effectively deliver activated CASP8 into primary human breast cancer cells overexpressing HER-2 to induce apoptosis and explore the underlying mechanism.
Materials and Methods:
HER-2 overexpressing primary human breast cancer cells were infected with lentivirus-like particles carrying Gag-CASP8.
Results:
After a 48h infection of primary human breast cancer cells with HER-2 by lentivirus-mediated Gag-CASP8, significant differences were observed in the survival rate, migration ability, S-phase number of cells, apoptosis rate, and intracellular activated CASP8 and caspase-3 levels in tumor cells compared with those in the control group (p < 0.05).
Conclusions:
Lentivirus-mediated Gag-CASP8 can deliver activated CASP8 into HER-2 overexpressing primary human breast cancer cells and induce apoptosis by activating caspase-3, a downstream apoptotic executive molecule. By blocking the S-phase to inhibit cell proliferation and migration, lentivirus-mediated Gag-CASP8 provides a reference for tumor gene therapy.
Introduction
Breast cancer is one of the most common cancers worldwide and the most common cause of cancer-related deaths in women. 1 Over the past 20 years, in addition to cytotoxic chemotherapy, drug therapy for breast cancer has evolved into a more targeted approach that has considerably improved patient outcomes; however, there remains no cure for metastatic breast cancer, and the 5-year overall survival rate of these patients is only 23%. 2 A significant and important finding in breast cancer treatment is the emergence of drug-resistant cancer cells. 3 –5 Therefore, to further improve patients' clinical outcomes and survival, new strategies are required for drug-resistant breast cancer.
Apoptosis is a naturally occurring process of programmed cell death, which plays a very important role in organismal homeostasis. Failure to regulate apoptosis is a key aspect of cancer development 6 because it produces a net increase in proliferation. 7 Although certain oncogenic mutations divert cells from apoptotic pathways, leading to tumor progression or metastasis, silencing some tumor suppressor genes may also increase the risk of tumor formation and progression; for example, the expression of caspase-8 (CASP8) and tumor suppressor genes in breast cancer cells is reduced by promoter methylation. 8 Therefore, apoptosis is a biological link between cancer genetics and therapy, with genes and proteins controlling apoptosis, presenting potential drug targets.
CASP8 plays an important role in the enzyme cascade leading to apoptosis. Adenovirus (Adv)-mediated CASP8 transfer has been demonstrated to induce apoptosis of U251 human glioma cells and inhibit their growth as xenograft tumors in vivo. 9 Moreover, introduction of CASP8 using the Adv-CASP8 vector enhanced the apoptosis of human gastric cancer cells and inhibit their peritoneal dissemination, 9 suggesting potential of CASP8 in breast cancer treatment.
Virus-like particles (VLPs) are self-assembled, nonreplicating, nonpathogenic, and genome-free particles with size and structure similar to those of intact infectious viruses. VLPs are attractive vaccine candidates because of their safety and efficacy in stimulating cellular and immune responses. 10 –13 Moreover, VLPs provide efficient transport vehicles for foreign genes and small molecules. 14 –16 A cytochrome P450 enzyme-expressing phage, P22 VLP, has been developed for the combined treatment of breast cancer by binding to photosensitizers and targeting their surface domains. 17 –19 The HIV group-specific antigen Gag effectively forms VLPs, even in the absence of other viral proteins. 20 A Gag-VLP pseudoprotein with vesicular stomatitis virus G glycoprotein (VSV-G) efficiently enters a broad range of cells, thereby enhancing the cytoplasmic delivery of VLP components and their immunogenicity. 21
Previously, the authors reported that CASP8-VLPs with the VSV-G pseudotype entered breast cancer cells and induced apoptosis. In a 4T1 mouse model of breast cancer, tumor growth was significantly inhibited when Gag-CASP8-VLPs were injected into tumors. 22 Herein, a Gag-CASP8 fusion protein capable of forming mature VLPs in the presence of HIV protease was generated, thus allowing the expression of active CASP8. This Gag-CASP8 was evaluated in human epidermal growth factor receptor 2 (HER-2) overexpressing primary breast cancer cells to assess its clinical potential for treatment of this drug-resistant breast cancer.
Materials and Methods
Cells and antibodies
To isolate HER-2 overexpressing primary human breast cancer cells, informed consent was obtained from patients before surgery (Consent Form No. K2020-029/March 29, 2020). Surgeries were performed in cooperation with the Department of Thyroid and Mammary Surgery of the First Affiliated Hospital of Guizhou University of Traditional Chinese Medicine, where the patients were hospitalized from January 2019 to April 2020. After preoperative fine-needle aspiration biopsy, the patients, aged 28–48 years, were diagnosed with HER-2 overexpressing breast cancer by pathologists.
Stable primary lines of HER-2 overexpressing breast cancer cells were isolated from the tumors of 3 patients. These cells were routinely cultured in Dulbecco's modified Eagle's medium (DMEM; HyClone) with 20% fetal bovine serum (FBS; HyClone) in an incubator at 37°C and 5% CO2.
The following antibodies were used: CA15-3 rabbit polyclonal, rabbit antihuman CASP8 (Proteintech Group, Rosemont, IL), mouse antihuman P24 (Abcam, UK), anticaspase-3 (Abcam), and horseradish peroxidase-conjugated goat antimouse IgG H&L (Abcam).
Plasmids
Three lentivirus-packaged plasmids (Δ8.2, VSV-G, and Lent-Gag-CASP8) were provided by Dr. Zhujun Ao of the University of Manitoba, Canada. As described previously, 23 in the SVCMV-Gag-Pol plasmid, HIV Pol was replaced with full-length human CASP8 cDNA, amplified by polymerase chain reaction, to construct a pSVCMVGag-CASP8 fusion protein-expressing plasmid. The HIV-1 protease cleavage site (SQNY/PIV) was inserted between Gag and CASP8. Auxiliary packaging of pCMV-Δ8.2 and VSV-G expression has been described previously. 24 Three HIV-1 RT/IN/Env defective proviral plasmids (ΔRI/E−/Gluc+) for the generation of stable cell lines have been described previously. 23
Preparation and transfection of VLPs
Lentivirus-like particles were collected from a culture of 293T human embryonic kidney cells (the sources were human embryonic kidney cells, purchased from the Chinese Academy of Sciences Cell Bank, cultured in an incubator at 37°C and 5% CO2 and tested for mycoplasma) and cotransfected, using polyethylenimine, with Lent-Gag-CASP8, Δ8.2, and VSV-G to obtain Gag-CASP8-VLPs. The same lentiviral particles were also cotransfected with Δ8.2 and VSV-G to obtain Gag-VLPs.
Culture and identification of HER-2 overexpressing primary human breast cancer cells
HER-2 overexpressing primary human breast cancer cells were successfully isolated from breast cancer tissues diagnosed as the HER-2 overexpressing type. Primary cells were cultured using an improved tissue block culture method, purified by an enzyme digestion time difference method. HER-2 overexpression was confirmed, and identified by immunohistochemistry and flow cytometry. 25
Infection of HER-2 overexpressing human breast cancer primary cells with Gag-CASP8-VLPs
Purified HER-2 overexpressing primary human breast cancer cells were infected with Gag-CASP8-VLPs at volume gradients of (1:25), (1:50), and (1:100). Their survival was measured using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), and half-inhibitory volumes were calculated to select the optimal action volume.
Lentivirus-like particle infection HER-2 overexpressing primary human breast cancer cells
Before infection, HER-2 overexpressing primary human breast cancer cells were seeded into 6- or 96-well plates at a density of 2 × 104 cells/mL and cultured for 24 h. Equal amounts of Gag-CASP8-VLPs or the empty control vector Gag-VLPs were added to DMEM at a volume ratio of 1:10, followed by incubation with cells for 24 and 48 h. Survival, migration, apoptosis, cell cycle phase, and apoptosis-related protein levels were then determined.
Cell viability assays
Equal quantities of Gag-CASP8-VLPs or Gag-VLPs were added to the primary human breast cancer cells with HER-2 overexpression cultured in 96-well plates, and viability was measured 24 and 48 h after treatment. Medium containing 0.5 mg/mL MTT was added to each well and cells were cultured for 3–5 h. After crystallization, the original medium was aspirated and dissolved by adding 150 μL DMSO. Cells were gently shaken at room temperature for 5 min to completely dissolve the crystals. Absorbance was measured at 570 nm as described previously. 26
Cell migration assays
The HER-2 overexpressing primary human breast cancer cells cultured in six-well plates were treated with the same amount of Gag-CASP8-VLPs or Gag-VLPs. After 48 h, the confluent cell sheet was scratched perpendicularly to the cell surface with the tip of a scalpel. Cells were washed three times with sterile phosphate buffered saline (PBS) to remove the scratched cells. Culture medium containing 1% FBS was used as negative control, with 1% FBS containing Gag-CASP8-VLPs used for drug stimulation. There were two duplicate wells for each group. Scratch widths of different treatment groups were observed after 0 and 48 h. Cell migration (%) was calculated using the formula (0 h scratch width–48 h scratch width)/(0 h scratch degree) × 100% as described previously. 26
Flow cytometric cell cycle assays
Cells were collected 48 h after infection, washed with PBS at 4°C, and pelleted at 800g for 5 min. Cells were resuspended in 1 mL of PBS and incubated with 2 mL of 70% alcohol at 4°C for 30 min. After washing with 1 mL PBS, cells were incubated with 500 μL RNase A (20 μg/mL in PBS) at 37°C for 30 min. After a final wash with 1 mL PBS, cells were incubated with 500 μL propidium iodide (PI) solution (50 μg/mL in PBS) at room temperature for 30 min in the dark, filtered into a flow tube with a 300-mesh sieve, stored at 4°C, and assayed by flow cytometry as previously described. 27
Apoptosis assays detection by acridine orange/ethidium bromide staining
After 48 h of incubation of HER-2 overexpressing primary human breast cancer cells with equal amounts of Gag-CASP8-VLPs or Gag-VLPs in a 96-well plate, acridine orange (AO; 0.5 mg/mL) and ethidium bromide (EB; 0.5 mg/mL) were mixed to make AO/EB. This AO/EB mixture (10 μL) was added to each well and the cells were observed by inverted fluorescence microscopy ( × 200). Apoptosis was quantified as previously described. 28
Flow cytometric apoptosis assays
Human HER-2 primary breast cancer cells were collected 48 h after infection, washed with PBS at 4°C, suspended in 1 mL 1 × binding buffer, and pelleted at 800 g for 5 min. The supernatant was discarded and 100 μL 1 × binding buffer was added to each tube, followed by the addition of 5 μL annexin V–fluorescein isothiocyanate. Tubes were incubated at room temperature away from light for 10 min. Then, 5 μL PI was added and the tube was incubated at room temperature in the dark for 5 min, followed by the addition of PBS to 500 μL and gentle mixing. Flow cytometry was performed within 1 h as described previously. 28
Western blotting
Primary breast cancer cells were collected 24 h after infection and lysed in RIPA lysis buffer. Proteins were separated by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Gag-VLPs and Gag-CASP8-VLPs were also lysed in 1% Triton X-100 and separated by 12% SDS-PAGE. CASP8 and the viral protein Gag-p24 were detected by Western blotting as described previously. 26
Statistical analysis
SPSS 22.0 software was used for all statistical tests. Data are expressed as (
Results
Lentivirus-like particles carrying Gag-CASP8 are packaged
Western blotting showed that P24, Gag-CASP8, and activated CASP8 protein fragments were expressed in the Gag-CASP8-VLP group. The Gag-VLP group expressed only the P24 protein, indicating that the lentivirus-like particles carrying CASP8 and the empty carrier had been successfully packaged (Fig. 1A).
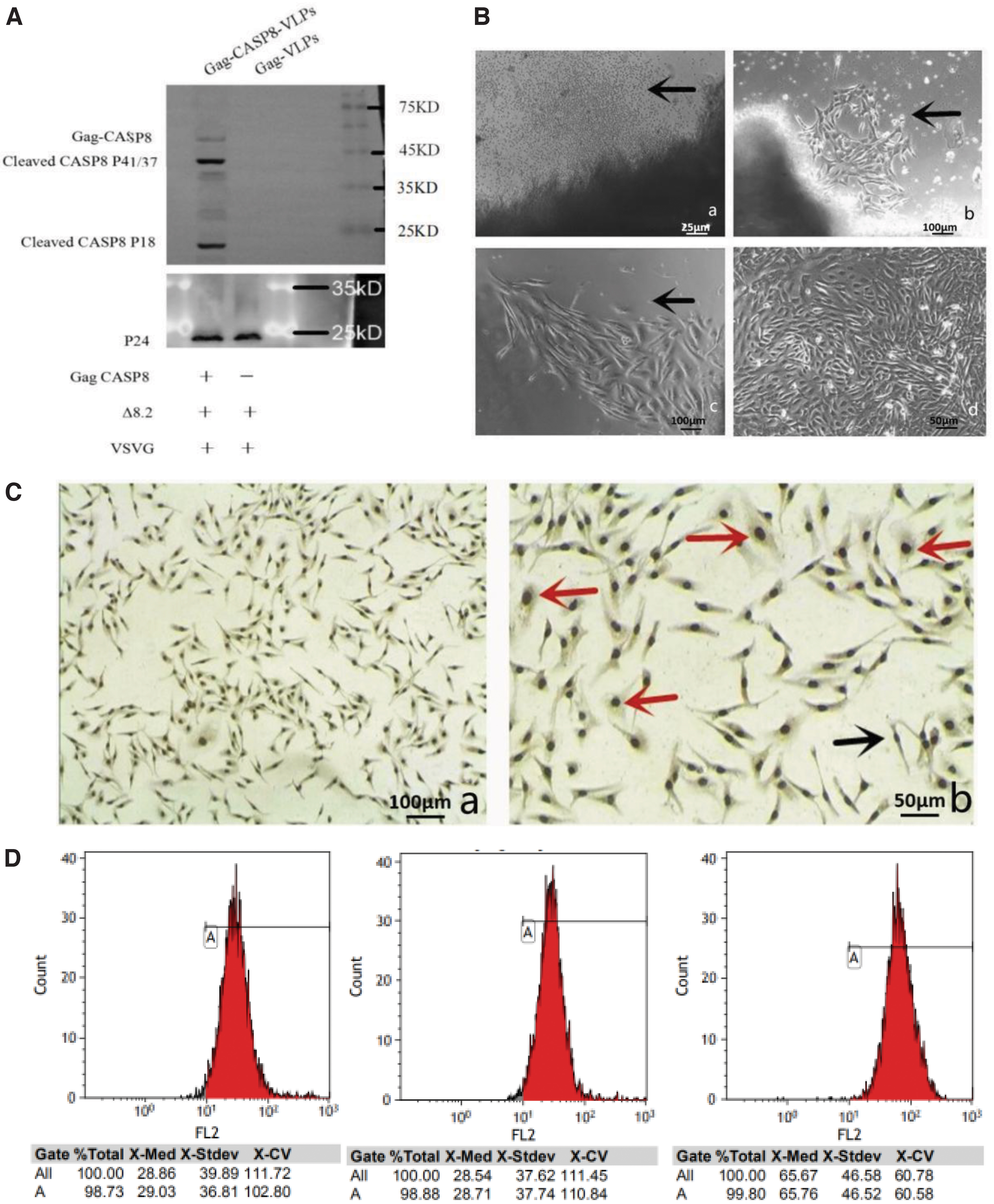
Culture and identification of HER-2 overexpressing primary human breast cancer cells
Human breast cancer tissue blocks were tightly adhered to culture surfaces after 1 d, showing poor organization and appearing as a dark shaded area, surrounded by cells whose morphology deviated from the typical round shape, with small volumes and refractile (Fig. 1B-a). After 3 d, cells were scattered and adhered to the culture surface (Fig. 1B-b). At 5 d, after most of the adherent cells had proliferated, nuclei were more prominent, with evidence of cell fusion, covering ∼50% of the culture bottle (Fig. 1B-c). After 7 d, cells formed intercellular contacts, exhibiting a typical “aura” covering the culture bottle (Fig. 1B-b).
After 5 d, human breast cancer cells were cultured; after a one-time trypsin digestion to remove fibroblasts, CA153 immunohistochemical staining and flow cytometry were performed. Microscopic observation revealed a large number of cell nuclei and CA153-positive human breast cancer cells, forming thin fibers (Fig. 1C). Flow cytometry revealed CA153 expression in ∼99% of cells in all three human breast cancer cell lines (Fig. 1D).
Gag-CASP8-VLPs infect HER-2 overexpressing human breast cancer primary cells using half-inhibitory volume
MTT assays showed that survival after Gag-CASP8-VLP infection at the three volume ratios of Gag-CASP8-VLPs for 24 and 48 h was significantly lower (p < 0.05) than those of the Gag-VLP and PBS groups. The half-inhibitory volume ratio of Gag-CASP8-VLPs was 0.088 (1:11) at 24 h and 0.141 (1:7) at 48 h (Fig. 2A).
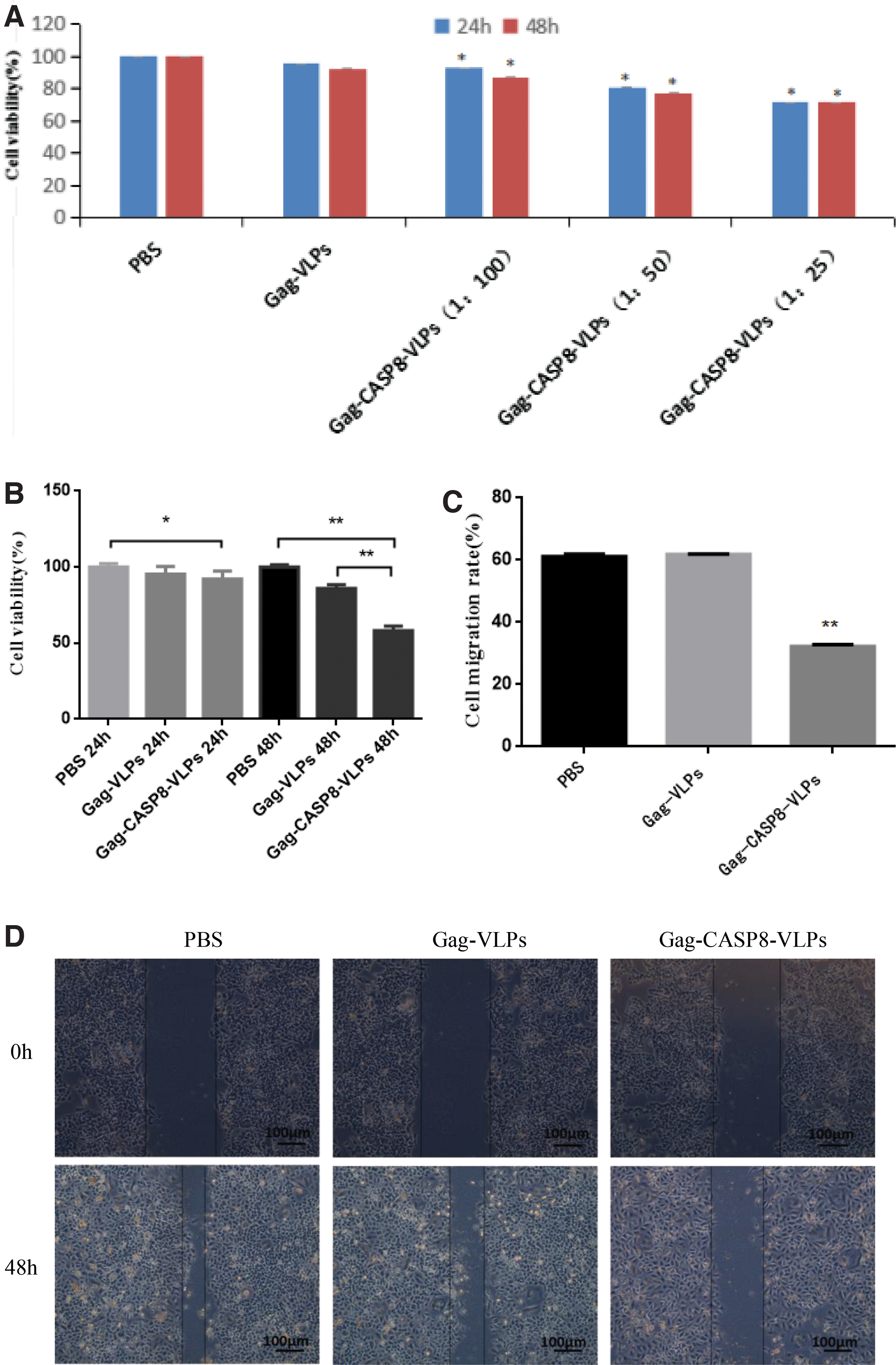
Gag-CASP8-VLP decreases cell viability
The survival of HER-2 overexpressing primary human breast cancer cells was 100.00% ± 2.22% in the PBS control group, 95.30% ± 5.01% in the Gag-VLP group, and 92.22% ± 5.09% in the Gag-CASP8-VLP group 24 h after infection with lentivirus-like granulation. Survival of the Gag-CASP8-VLP group was significantly lower than that of the PBS group (p < 0.05; Fig. 2B).
After 48 h of infection with lentivirus-like granulations, the survival of HER-2 overexpressing primary human breast cancer cells was 100.00% ± 1.53% in the PBS group, 85.97% ± 2.33% in the Gag-VLP group, and 58.20% ± 2.96% in the Gag-CASP8-VLP group. The cell survival rate of the Gag-CASP8-VLP group was significantly lower than that of the Gag-VLP and PBS groups (p < 0.01; Fig. 2B).
Gag-CASP8-VLP decreases cell motility
A scratch assay revealed that the migration ability of HER-2 overexpressing primary human breast cancer cells was significantly less than that of the Gag-VLP and PBS controls 48 h after treatment with lentivirus-like particles (Fig. 2D); the cell migration rate of the Gag-CASP8-VLP group (32.20% ± 0.15%) was lower than that of the Gag-VLP group (61.85% ± 0.11%) and PBS group (61.16% ± 0.08%), **p < 0.01 (Fig. 2C).
Gag-CASP8-VLP alters the cell cycle
Forty-eight hours after infection of HER-2 overexpressing primary human breast cancer cells, the proportion of cells in S-phase with Gag-CASP8-VLP infection was 30.90% ± 0.56%, significantly higher than that in the Gag-VLP (24.70% ± 0.34%) and PBS (19.93% ± 0.25%) controls (p < 0.01; Fig. 3A, 3D).
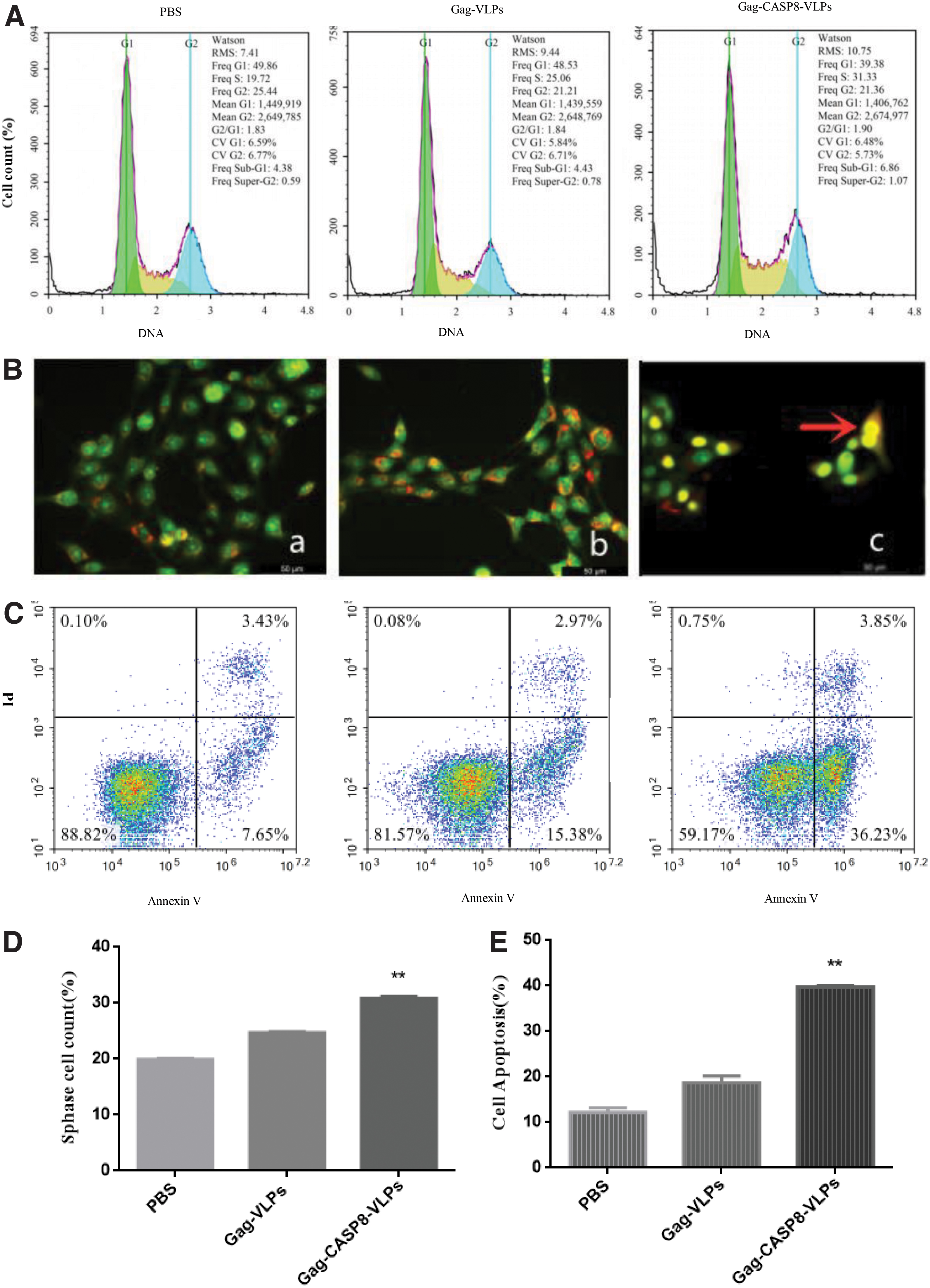
Gag-CASP8-VLP induces apoptosis
Apoptosis detection by AO/EB staining
Fluorescence microscopy showed HER-2 overexpressing primary human breast cancer cells to be living, with green chromatin and a uniform and regular structure. Large number of cells in the Gag-VLP and the PBS groups were observed. Some cells in the Gag-CASP8-VLP group with yellow-green or yellow chromatin stain were early apoptotic cells. The late apoptotic cells showed orange fluorescence, and there was a significant increase in number of apoptotic cells in the Gag-CASP8-VLP group compared with that in the Gag-VLP and the PBS control groups (Fig. 3B).
Cell apoptosis was detected by flow cytometry
By flow cytometry, the apoptosis rate of the Gag-CASP8-VLP group (39.46% ± 1.41%) was significantly higher than that of the Gag-VLP (20.31% ± 1.42%) and PBS (12.78% ± 1.30%) groups, 48 h after infection by chronic disease VLPs of HER-2 overexpressing primary human breast cancer cells (p < 0.01; Fig. 3C, 3E).
Gag-CASP8-VLP alters protein expression
Western blotting demonstrated P24, Gag-CASP8, pro-CASP8, cleaved-CASP8, and Caspase-3 protein expression in Gag-CASP8, pro-CASP8, and cleaved-CASP8, respectively, in the Gag-CASP8 group after infection with lentivirus-like particles for 24 h. P24 and a low level of pro-CASP8, cleaved-CASP8, and caspase-3 protein expression were observed in the Gag-VLP control, whereas only a low level of pro- and cleaved-CASP8 protein expression was observed in the PBS control (Fig. 4A).
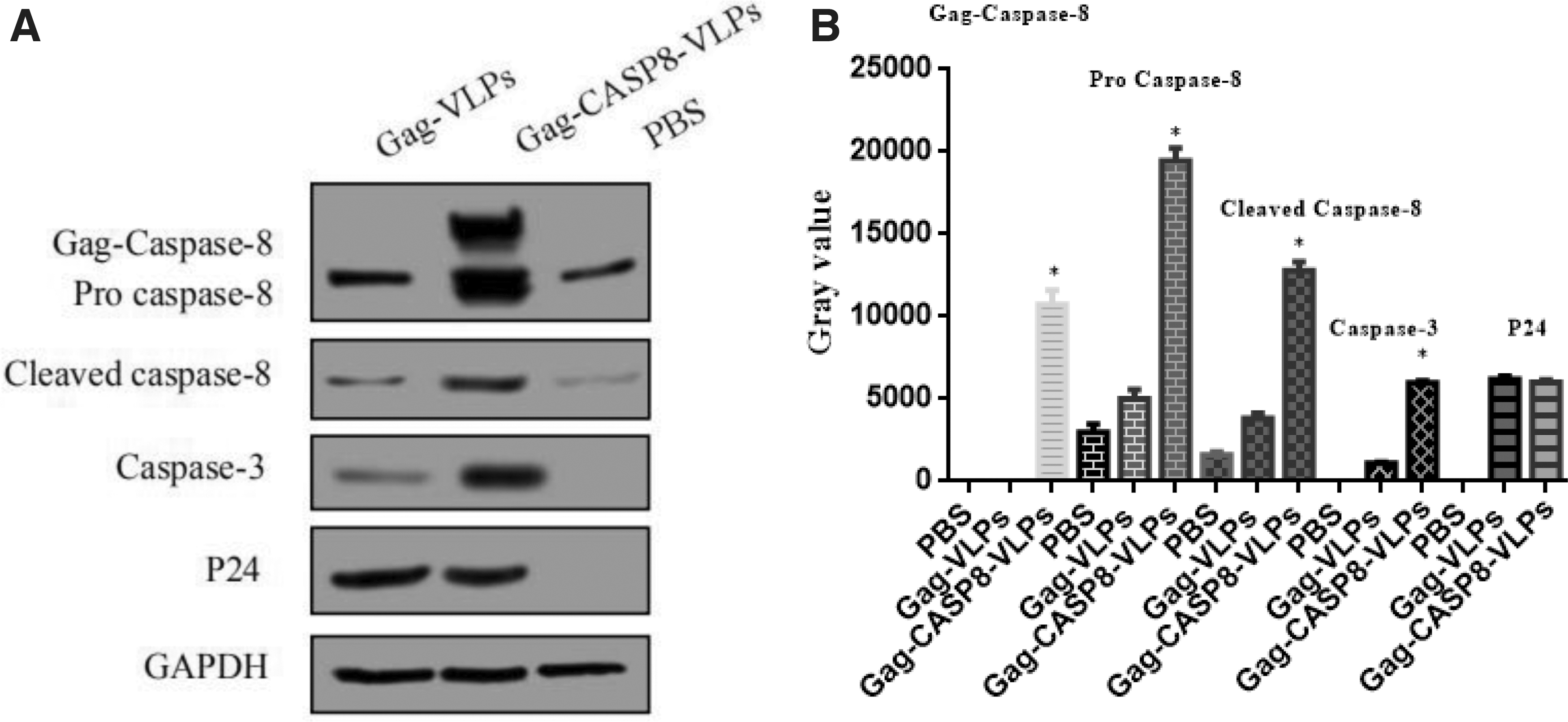
The grayscale intensities of Gag-CASP8, pro-CASP8, cleaved-CASP8, and caspase-3 protein were significantly higher in the Gag-CASP8-VLP group than in the Gag-VLP group and PBS groups (p < 0.05); there was no significant difference in the grayscale intensities of P24 protein between the Gag-CASP8-VLP and Gag-VLP groups (p > 0.05; Fig. 4B).
Discussion
Herein, lentivirus-like particles containing the apoptotic protein CASP8 were packaged and assembled using an HIV Gag-CASP8 fusion protein. Through the pseudotype of VSV-G, Gag-CASP8-VLPs were able to enter HER-2 overexpressing primary human breast cancer cells, inhibit cell proliferation and migration, and kill by inducing apoptosis; this mechanism was further studied.
A half-inhibitory volume is needed to determine gene therapy dosages for cell killing; the dose is too low and cannot achieve the goal of treatment 29 but allows quick evaluation of growth inhibition. After calculating the half-inhibitory volume for Gag-CASP8-VLPs, it was first verified whether Gag-CASP8-VLPs could inhibit the proliferation of human breast cancer cells at 24 and 48 h after infection. As the proliferation inhibition rate showed an increasing trend over time and was significantly higher at 48 h than at 24 h, 48 h was selected as the time point for all subsequent experiments.
Determination of cell migration ability, which is crucial in pathological processes such as tumorigenesis, leads to better analysis and understanding of multiple pathological processes. To elucidate whether Gag-CASP8-VLPs could inhibit the proliferation of human breast cancer cells and affect their migration ability, a scratch assay was used. Gag-CASP8-VLPs significantly inhibited the scratch-healing ability of monolayers of cultured human breast cancer cells, suggesting that Gag-CASP8-VLPs inhibited the proliferation of human breast cancer cells and reduced their migration ability.
The authors conducted cell cycle studies to further investigate the inhibition mechanisms of tumor cell proliferation and migration. With Gag-CASP8-VLP, the proportion of cells in S-phase was significantly higher than in the Gag-VLP and PBS controls. This phenomenon may be due to the ability of Gag-CASP8-VLPs to arrest DNA replication or impact the cells' ability to repair DNA damage. As a result, most cells cannot normally proceed into the next phase and are retained in S-phase, resulting in the inhibition of cell proliferation and migration.
Apoptosis is triggered through two main signaling mechanisms—the intrinsic and extrinsic pathways. 30,31 In both, signal transduction activates the cysteine protease, or caspase, family. Among the 11 human caspases, activation of CASP8 is central to apoptosis. 32 –34 Cleaved-CASP8 activates downstream caspase-3 and caspase-7, causing apoptosis. 35 Therefore, CASP8 is an effective proapoptotic molecule. Herein, lentivirus-like particles were established that could directly deliver activated CASP8 into tumor cells. The CASP8 protein was effectively released from the Gag-CASP8 fusion protein by cleavage with the HIV protease to assemble CASP8-VLPs. Most of the CASP8 protein in VLPs was present in the active state (p41/37 and p18) and induced apoptosis (Figs.1A and 3B, C, E). Caspase-3, an apoptosis-executing molecule downstream of the apoptosis promoter CASP8, had higher expression in the experimental apoptosis group than in the control group (Fig. 4A, B). These findings suggest that one of the mechanisms for induction of tumor cell apoptosis involves the activation of caspase-3 downstream of CASP8.
Previous studies have shown that activation of CASP8 may occur through multiple mechanisms, in addition to FAS-L stimulation. Under certain physiological conditions, high expression levels of CASP8 precursor proteins may lead to autooligomerization, self-cleavage, and activation of CASP8 in the absence of any external apoptotic signal. 36 To effectively mediate the entry of lentiviral particles into cancer cells, researchers have proposed using the VSV-G pseudotype, a gene transfer mechanism commonly used in lentiviral vectors. In contrast to general lentiviral vector systems, Gag-CASP8-VLPs do not contain any viral genomic material, precluding the potential risk of viral genome integration into the host chromosome. This feature is critical, as Gag-CASP8-VLPs may be used as anticancer agents if their anticancer effects and safety are demonstrated in animal models. Gag-CASP8-VLPs effectively induced tumor cell death and inhibited tumor formation in a mouse model, suggesting their suitability as a new antitumor strategy. 22
As Gag-CASP8-VLPs can enter cells in the presence of VSV-G on VLPs, these VLPs are expected to target both tumor cells and normal cells. Although there were no significant side-effects after injection of Gag-CASP8-VLPs into mice, further research is required to optimize their targeting. 37 A key aspect of this strategy is the specific or preferential targeting of cancer cells by CASP8-VLPs. Previous studies have revealed that several unique receptor proteins, such as HER-2, are overexpressed in multiple tumor cells by ligands. 38,39 HER-2 is one of the most characteristic tumor-associated antigens in cancer therapy and plays a vital role in breast cancer growth and progression. In patients with drug resistance, both primary and secondary tumors exhibit poor prognosis. The resistance mechanism is highly complex, with multiple drug-resistance mechanisms coexisting and multiple factors remaining unexplored. Therefore, HER-2 overexpression in cancers and its availability in the extracellular domain make it a clinically useful target for developing tumor-specific drugs.
Placing HER-2 on the surface of vaccinia viruses coated with monoclonal antibodies against tumor-associated antigens is a step toward targeted therapy of selective tumor cells. 39 Recent studies have shown that biocatalytic phage P22 VLPs can be chemically coated with targeted ligands containing polyethylene glycol–folic acid or estradiol. These studies also demonstrated that VLPs with targeted portions on their surfaces were recognized and internalized into tumor cells, including ER+ breast cancer cells. Therefore, this system could provide an opportunity to attach tumor-specific targeting molecules to the surface of VLPs and specifically or selectively drive active CASP8 into cancer cells to achieve specific anticancer activity.
Conclusions
Gag-CASP8-VLPs could be assembled using an HIV Gag-CASP8 fusion protein and effectively introduced in vitro into HER-2 overexpressing primary breast cancer cells, causing cells to be stagnated in the S-phase of the cell cycle, while their proliferation and migration were inhibited. As the study was limited to the cellular level and insufficient research was performed on the underlying mechanism, further animal experiments and in-depth mechanistic research are needed to evaluate the feasibility of this new antitumor strategy. Sufficient evidence is required to optimize the tumor cell-targeted specificity of this antitumor system and provide an effective anticancer treatment.
Footnotes
Authors' Contributions
D.T. and M.W. conceived and designed this study; M.W., X.L., W.X., Y.L., and L.Z. performed the experimental work; X.L., Z.Z., X.C., and L.L. maintained the primary cell culture; M.W., X.L., L.Z., and Y.L. were involved in Western blot analysis. X.C., M.Y., and L.Q. performed the cell viability assay; M.W. and X.L. contributed to flow cytometry analyses, Hoechst AO-EB staining, and drafted the article. D.T. and W.X. critically revised the article. M.W. and X.L. contributed equally to this study. All authors reviewed and approved the article before submission.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Natural Science Foundation of China Grant No. 81660749, by the Science and Technology Foundation of Guizhou Provincial Health Commission Grant No. GZWJKJ2020-1-241, and by the Science and Technology Foundation of Guizhou Provincial Health Commission Grant No. GAWKJ2021-521.