
Abstract
Objective:
To investigate the effects of M2 macrophage-derived exosomes (M2-Exos) on proliferation, migration, and apoptosis of gastric cancer cells in the tumor microenvironment and to further explore their possible molecular mechanism.
Materials and Methods:
M2 macrophages were induced from THP-1 cells and identified by qRT-PCR. Exosomes were extracted by ultracentrifugation and identified by transmission electron microscopy (TEM), nanoparticle tracking analysis (NTA), and Western blot analysis. Fluorescence labeling was used to detect the internalization of exosomes in receptors. The proliferation, migration, and invasion of AGS and HGC27 cells were determined by EdU and MTS, wound healing and Transwell assay, and flow cytometry, respectively. Proteins in the related pathway of M2-Exos affecting the progression of gastric cancer were detected by Western blot analysis.
Results:
In this study, M2 macrophages and M2-Exos were successfully obtained. The purified M2-Exos were observed as small round vesicles with diameters of 50–90 nm and positive expression of CD63, CD9, and TSG101. Besides, M2-Exos can be effectively taken up and internalized by AGS and HGC27 cells. Cell behavior studies showed that M2-Exos promoted proliferation and migration and inhibited the apoptosis of AGS and HGC27. Further research illustrated that M2-Exos promoted the phosphorylation of P38 and high expression of programmed death ligand 1 (PD-L1).
Conclusions:
This study demonstrated that M2-Exos promoted proliferation and migration and inhibited the apoptosis of gastric cancer cells. Mechanically, M2-Exos may promote gastric cancer progression through the P38MAPK signaling pathway and achieve immune escape through elevating the expression of PD-L1.
Introduction
Gastric cancer is a highly prevalent disease worldwide and the third leading cause of cancer-related death. Meanwhile, gastric cancer ranks as an important barrier to increasing life expectancy. 1 It is reported that its morbidity and mortality rates in China account for more than 40% of the world's total, seriously threatening the health of people. 2 Different from gastric cancer in other East Asian regions, the early diagnosis rate of gastric cancer in China is low and, once diagnosed, it is usually at an advanced stage. Besides, due to lack of treatment strategies for advanced gastric cancer, coupled with the high aggressiveness and drug resistance of cancer, the treatment result is not ideal, and the prognosis is poor. 3 Therefore, there is urgent need to develop new treatment methods and more indicators of risk and prognosis for gastric cancer.
With the rise of tumor immunotherapy, the application of immunotherapy in treatment of advanced gastric cancer has also shown promising effects. In the ATTRACTION 2 study, programmed death-1 (PD-1) inhibitor nivolumab improved the objective remission rate and prolonged the median survival time of advanced third-line gastric cancer. 4 In the TOGA study, trastuzumab combined with chemotherapy extended the survival of patients with HER2-positive advanced gastric cancer to more than 16 months. 5 Although immunotherapy has made great progress in the treatment of advanced gastric cancer, it is not 100% effective. Tumors do not exist independently in the human body and often interact with the surrounding growth environment, which is often referred to as the tumor microenvironment (TME). The TME undergoes adaptive changes with formation and metastasis of tumors. Therefore, the TME has become a key point in the study of cancer progression.
Recently, a large number of studies have emphasized the importance of the interaction between tumor cells and the corresponding microenvironment in tumor progression. 6 Among them, tumor-associated macrophages (TAMs) play a particularly significant role and their performance varies greatly in different microenvironments. 7 According to the activation state and function, TAMs can be divided into classically activated (M1) macrophages and alternatively activated (M2) macrophages. 8 M1 macrophages participate in the positive immune response, play a role in proinflammatory and antitumor responses, and prevent tumor immune escape. 9 M2 macrophages are involved in the negative immune response, which has anti-inflammatory and protumor effects and helps the immune escape of tumor cells. 10 In fact, in most solid tumors, TAMs are usually a macrophage subpopulation with an M2 phenotype, and the positive correlation between TAM density and poor prognosis has been demonstrated in a variety of cancers. 11
Exosomes are an important communication medium between different cell types in the TME, carrying information from one cell to another and reprogramming the recipient cell. 12 Studies have reported that the exosomes secreted by M2 macrophages can not only affect some inflammation-related diseases but also affect the TME and have an impact on tumors by carrying the information of their source cells.12,13 However, most current studies have focused on exosomes secreted by cancer cells, and little is known about TAM-derived exosomes and their effects on cancer cells.
In this study, the authors sought to determine the effect of M2 macrophage-derived exosomes (M2-Exos) on gastric cancer. Furthermore, the potential molecular mechanisms related to M2-Exos in progression of gastric cancer were investigated, which will lay a foundation for the study of immune escape using TEM.
Materials and Methods
Cell culture and induction
The human acute monocytic leukemia cell line (THP-1) and gastric cancer cell lines (AGS and HGC27) were obtained from American Type Culture Collection (ATCC, Manassas, VA) and cultured in RPMI 1640 medium. For the induction experiment, 4 μL of PMA (100 ng/mL) was first added into THP-1 cells and cultured in an incubator to obtain adherent M0 macrophages. Subsequently, 40 mL of RPMI-1640 medium containing 10% FBS, IL-4 (20 ng/mL), and IL-13 (20 ng/mL) was added to obtain M2 macrophages. This study was approved by the Ethics Committee of China.
Isolation of exosomes
Exosomes were extracted from the supernatant of M2 macrophages by ultracentrifugation. Briefly, the supernatant was filtered with a 0.22-m Steritop™ sterile membrane and then centrifuged at 2500 g for 30 min, 8500 g for 30 min, and 120,000 g for 1.5 h. Subsequently, the precipitate was then resuspended in 15 mL of sterile PBS and centrifuged at 4000 g until the final volume was reduced to about 200 μL. Finally, the purified exosomes were stored at −80°C for later use.
Quantitative real-time PCR
The TRIpure reagent (Invitrogen) was applied to isolate the total RNA from samples, and the PrimeScript RT Kit (TaKaRa, Otsu, Japan) was used for reverse transcription. After the sample was prepared, the gene expression was detected with FAST SYBR™ Green Master Mix, and GAPDH was used as the internal control parameter. The 2−ΔΔCt method was used to calculate the fold changes in gene expression, and the experiments were conducted three times. Primers used were as follows: Arg1 forward, 5′-TGGACAGACTAGGAATTGGCA-3′, and reverse, 5′-CCAGTCCGTCAACATCAAAACT-3′; CD206 forward, 5′-GGGTTGCTATCACTCTCTATGC-3′, and reverse, 5′-TTTCTTGTCTGTTGCCGTAGTT-3′; IL-10 forward, 5′-CAGCTCAGCACTGCTCTGT-3′, and reverse, 5′-TCTTCACTCTGCTGAAGGCA-3′; and GAPDH forward, 5′-GGCTGCTTTTAACTCTGGTA-3′, and reverse, 5′-ACTGTGGTCATGAGTCCTT-3′.
Transmission electron microscopy
Transmission electron microscopy (TEM) analysis was conducted to observe the morphology of extracted exosomes. First, the purified exosomes were stained with 2% sodium phosphowolframate for 10 min and then washed with PBS. After drying for 35 min at room temperature, exosomes were stained with 1% uranyl acid and observed using TEM (Hitachi H7500 TEM, Tokyo, Japan).
Nanoparticle tracking analysis
The size distribution and concentration of the extracted exosomes were detected using NanoSight LM10. First, M2-Exos were diluted 10,000 times with PBS and mixed upside down.
Then, the diluted M2-Exos were tested, photographed, and recorded under professional guidance.
Western blot analysis
The target cell protein was obtained by the RIPA method and protein concentration was determined using the BCA kit. Then, 50 μg of prepared protein was taken for gel electrophoresis separation, and the isolated protein was electrically transferred to a PVDF membrane (Roche, Switzerland). After blocking with 5% nonfat dry milk, the PVDF membrane was subjected to incubation with primary antibodies, CD63 (1:1000, ab134045; Abcam), GM130 (1:1000, ab92726; Abcam), CD9 (1:1000, ab2787; Abcam), tumor susceptibility gene 101 (TSG101, 1:1000, ab125011; Abcam), p-P38 (1:1000, ab170099; Abcam), PD-L1 (1:1000, ab205921; Abcam), and GAPDH (1:1000, ab9485; Abcam), at 4°C overnight. On the following day, the membrane was incubated with the secondary antibody at 37°C for 1 h. After washing the membrane film with TBST, a luminescent solution was added for exposure in the gel imaging system. The protein content was analyzed using Quantity One software.
Fluorescence labeling
Fluorescence labeling was performed when AGS and HGC27 cells were fused to 50%. The purified exosomes were labeled with PKH-67 (green fluorescence) under dark conditions. After 24 h of culture, PKH-67-labeled exosomes were cocultured with AGS and HGC27 and then stained with rhodamine phalloidin (red fluorescence) and DAPI (blue fluorescence). Finally, the samples were observed under a fluorescence microscope.
5-Ethynyl-2-deoxyuridine assay
AGS and HGC27 cells at the logarithmic growth stage were spread on a 24-well plate and then cultured in different concentrations of M2-Exos (PBS and M2-Exos 40 μg/mL) for 48 h. The 5-ethynyl-2-deoxyuridine (EdU) solution was diluted with cell complete medium at the ratio of 1000:1 to prepare an appropriate amount of 50 μM EdU culture medium. In each well, 300 μL of EdU medium was added, which was then incubated for 2 h. After cell fixation, Apollo staining, DNA staining, and the proliferation activity of cells were observed and detected under a fluorescence microscope.
MTS assay
In brief, the cells adjusted to the appropriate concentration (1000–2000 cells/100 μL) were inoculated on 96-well plates. Twenty-four hours after the cells were completely attached to the well, the media containing PBS, M0-Exos, and M2-Exos were added, respectively, and the culture was set for different days (days 1, 2, 3, and 4). MTS solution was added into each well and the absorbance value was read at 490 nm as day 0. The values of days 1, 2, 3, and 4 were measured continuously at the same time interval.
Transwell assay
AGS and HGC27 cells were first adjusted to the appropriate concentration (2 × 104 cells/well). Subsequently, the upper chamber was filled with cells treated with PBS, M0-Exos, and M2-Exos, whereas the lower chamber was replaced by a medium supplement, 15% FBS. After 48 h, 4% paraformaldehyde was used for fixation and 0.1% crystal violet for staining, respectively. Five fields were randomly selected under an inverted microscope to represent the migration ability of cells in each group.
Wound healing assay
First, AGS and HGC27 cells were inoculated in a six-well plate for 24 h. When the cells were fully fused, a pipette tip was used to create a scratch wound on confluent cells in the center and the medium containing PBS, M0-Exos, and M2-Exos was added simultaneously. Migration of the entire wound area was observed with an inverted optical microscope (Oberkochen, Germany), and images were taken at 36 h with a camera connected to the microscope (Sony Cyber-Shot, Shanghai Suoguang Visual Products Co., Ltd., China). The cell migration ability was statistically analyzed according to cell healing.
Flow cytometry
First, treated cells were suspended and centrifuged for 5 min. Second, cells were washed and incubated in buffer solution with FITC-labeled Annexin V (Carlsbad, CA) for 15 min at 37°C. After that cells were centrifuged for 5 min to remove the supernatant, followed by adding propidium iodide. Finally, cell apoptosis was tested and calculated using a flow cytometer (FACScan, CA) and CELL Quest 3.0 software (BD Biosciences, Beijing, China).
Statistical analysis
All the data were analyzed using GraphPad Prism software (version 5.0). One-way ANOVA (followed by Dunnett's multiple comparison) was applied to assess the differences between groups; p < 0.05 indicated a significant difference between groups.
Results
Characteristics of M2 macrophages and M2-Exos
To identify whether the cells induced were M2 macrophages, qRT-PCR was applied to detect the markers of M2 macrophages. The qRT-PCR data showed that compared with M0 macrophages, M2 macrophages had higher mRNA expression of CD206 (p < 0.001), Arg-1 (p < 0.01), and IL-10 (p < 0.001), indicating that M2 macrophages were successfully induced (Fig. 1A). Subsequently, exosomes were extracted from M2 macrophages and then identified. As presented in Figure 1B, Western blot analysis revealed positive expression of characteristic surface antigens, CD63, CD9, and TSG101, in M2-Exos, while GM130 was not expressed in either M0-Exos or M2-Exos. In nanoparticle tracking analysis, the peak diameter of M0-Exos was bimodal, which was 164.9 and 131.1 nm, and the peak diameter of M2-Exos was unimodal, which was 157.6 nm (Fig. 1C). Using TEM, the purified M0-Exos and M2-Exos were observed as small round vesicles with diameters ranging from 50 to 90 nm (Fig. 1D).

The characteristics of M2 macrophages and M2-Exos.
Uptake and internalization of M2-Exos
To investigate the uptake and internalization of M2-Exos into recipient cells, AGS and HGC27 cells were first incubated with PKH67-labeled M0-Exos and M2-Exos. Then, the cells were stained with rhodamine phalloidin and DAPI. Using confocal microscopy, the authors observed that 90% of the M2-Exos were integrated into the AGS and HGC27 cells, with green fluorescence diffusing throughout the cells. Meanwhile, the authors observed that the blue fluorescent nuclei and green fluorescent exosomes were surrounded by a red fluorescent cytoskeleton, indicating that M2-Exos were taken up by GC cells (Figs. 2 and 3). However, in contrast, M0-Exos were almost not observed in cocultured gastric cancer cells.
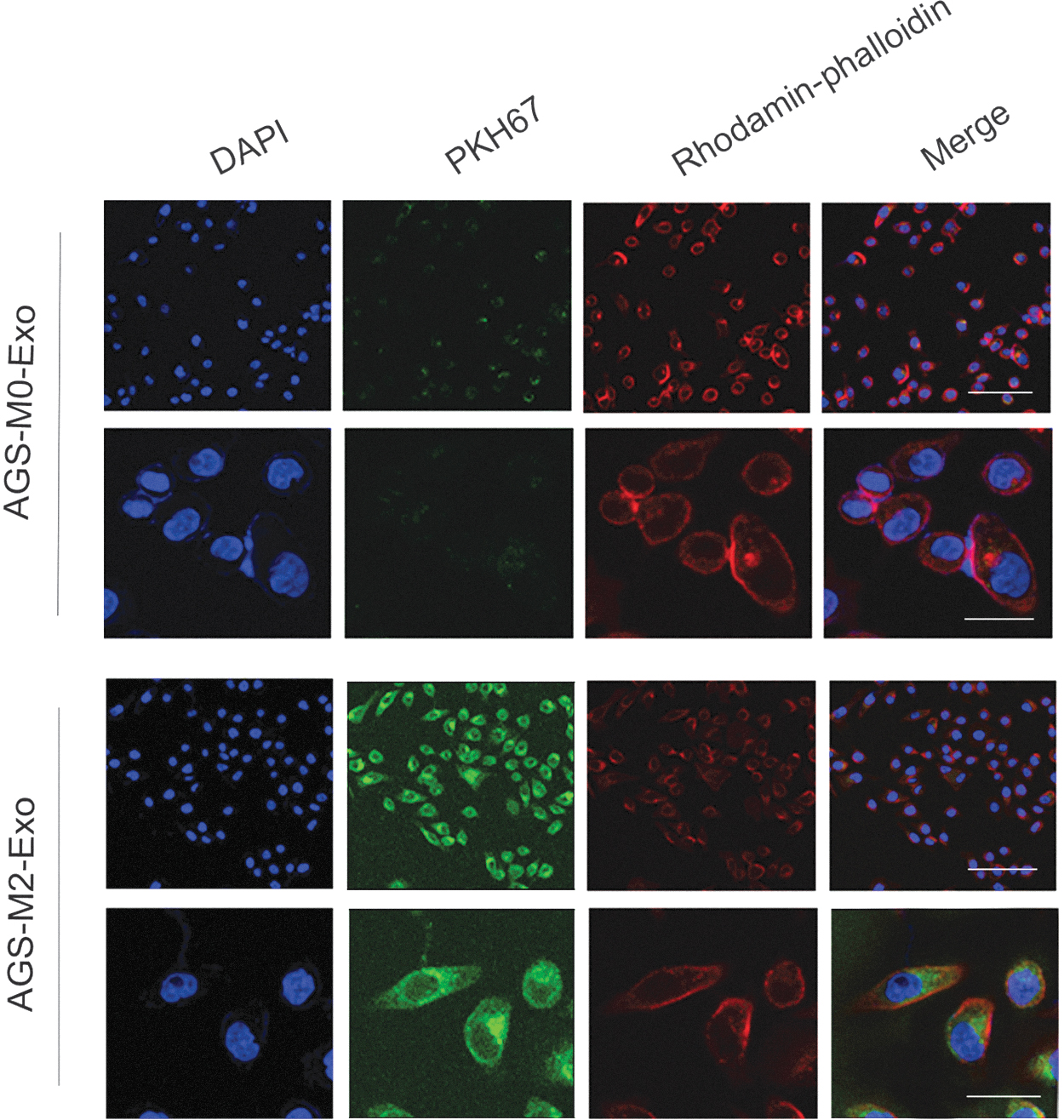
Uptake and internalization of M2-Exos in AGS cells. The uptake efficiency of M2-Exos by AGS cells detected by PKH67 and rhodamine phalloidin. Color images are available online.

Uptake and internalization of M2-Exos in HGC27 cells. The uptake efficiency of M2-Exos by HGC27 cells detected by PKH67 and rhodamine phalloidin. Color images are available online.
M2-Exos promoted proliferation of AGS and HGC27 cells
To explore the role of M2-Exos in progression of gastric cancer, the authors first detected the effect of M2-Exos on gastric cancer cell proliferation. In the EdU assay, strong fluorescence was observed in cells cocultured with M2-Exos, indicating that M2-Exos had the ability to promote proliferation of AGS and HGC27 cells. On the contrary, weak fluorescence in gastric cancer cells confirmed that M0-Exos had no effect on cell proliferation (Fig. 4A, B, D, E). Furthermore, MTS was applied to detect the effects of different treatment times with M0-Exos and M2-Exos on proliferation of AGS and HGC27 cells. The results showed that the proliferation activity of AGS and HGC27 cells treated with M2-Exos increased in a time-dependent manner in the first 3 d. By day 4, it was slightly lower, but still higher than the M0-Exo group. Conversely, M0-Exos had no significant effect on promotion of cell proliferation (Fig. 4C, F).
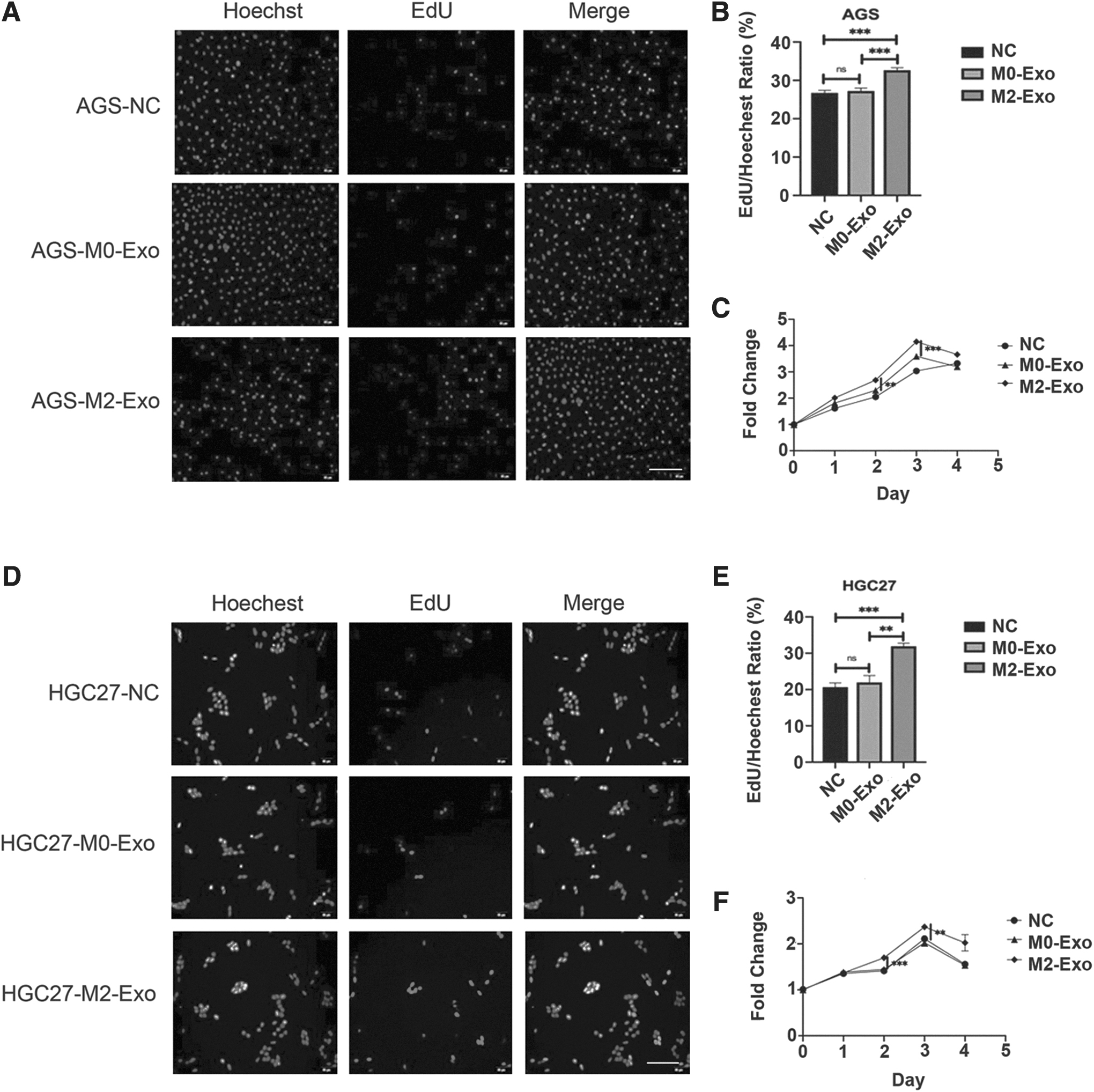
M2-Exos promote proliferation of AGS and HGC27 cells.
M2-Exos promoted migration of AGS and HGC27 cells
To evaluate the effect of M2-Exos on cell migration, Transwell and wound healing assays were conducted. As depicted in the Transwell assay, the number of cells in the M2-Exo group was significantly higher than that in the M0-Exo group and NC group (Fig. 5A). In the wound healing assay, the relative wound width of the M2-Exo group was sharply narrower than the M0-Exo group in both AGS and HGC27 cells. Conversely, it was observed that the M0-Exo group inhibited the migration rate of AGS and HGC27 cells when compared with the NC group (Fig. 5B).
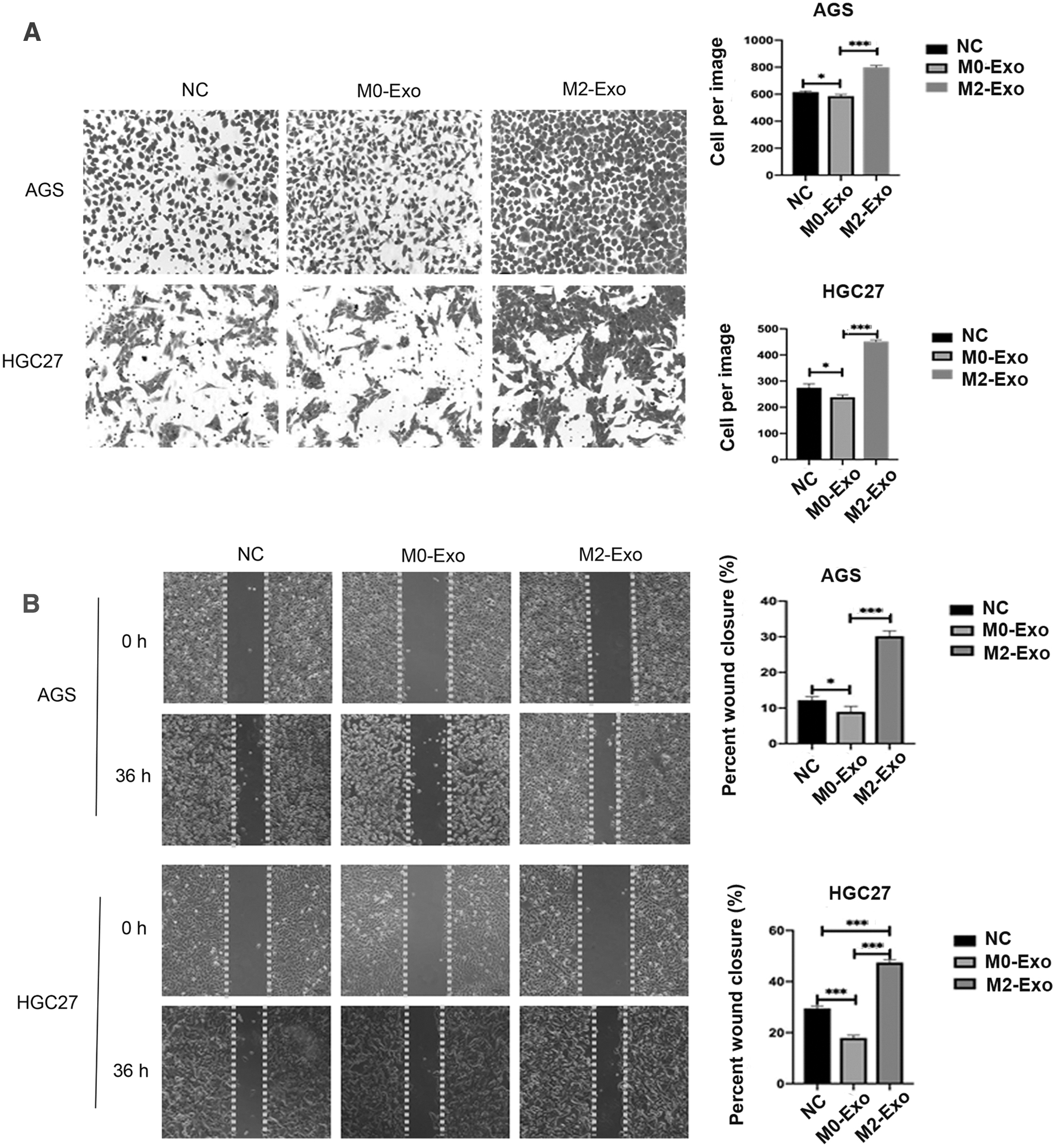
M2-Exos promote the migration of AGS and HGC27 cells.
M2-Exos inhibited the apoptosis of AGS and HGC27 cells
To explore the effect of M2-Exos on cell apoptosis, the authors detected the apoptosis rate of each group of AGS and HGC27 cells. As revealed in Figure 6, M2-Exo treatment led to a significant decrease in the apoptosis ratio of AGS and HGC27 cells. More specifically, after treatment with M2-Exos, the apoptosis rate of AGS and HGC27 cells decreased to 6.04% and 2.01%, respectively. Of note, although the apoptosis rate of AGS and HGC27 cells changed slightly after treatment with M0-Exos, there was no statistical difference.
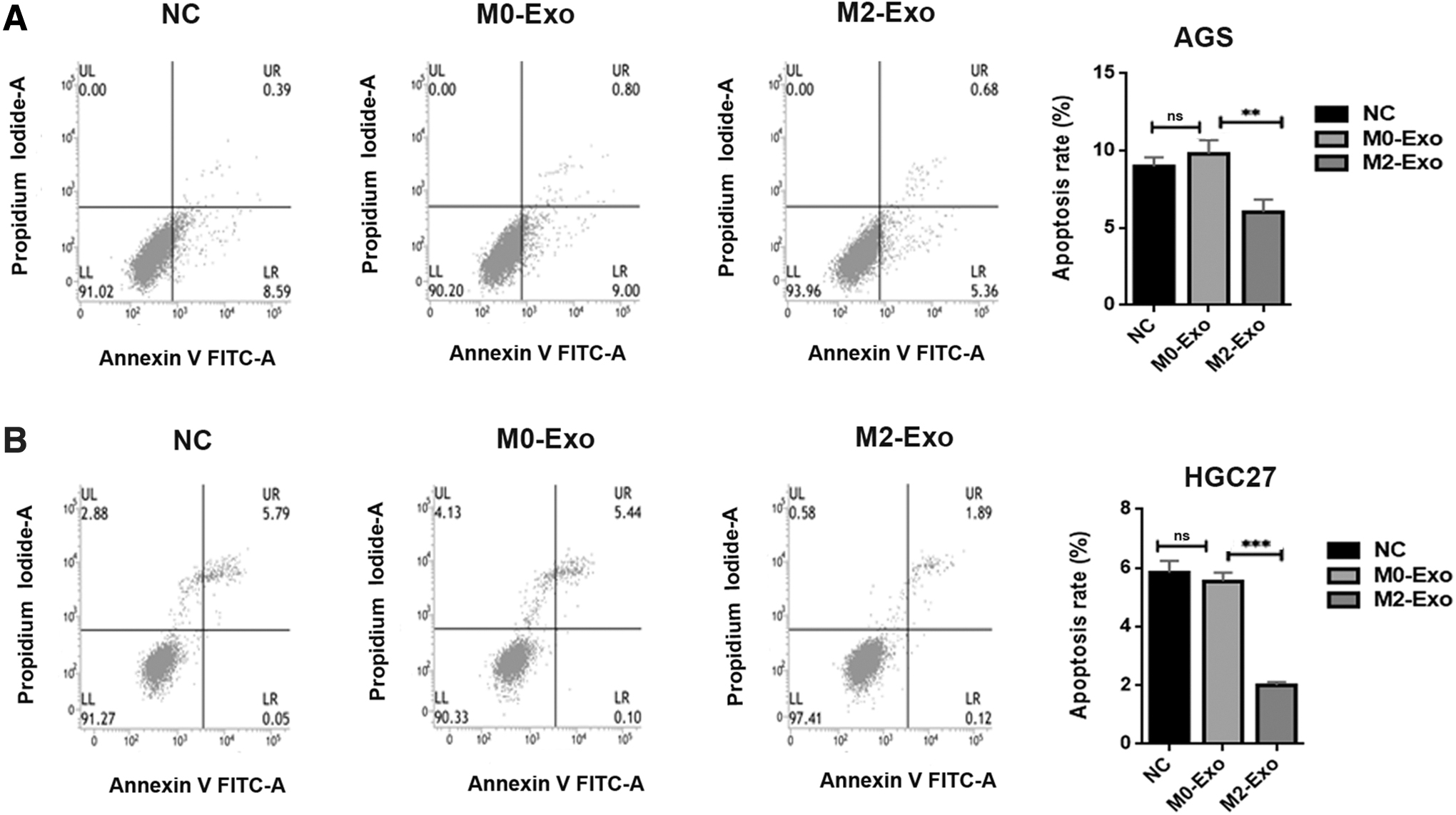
M2-Exos inhibit the apoptosis of AGS and HGC27 cells.
M2-Exos accelerated progression of gastric cancer by activating the P38MAPK pathway and upregulating the expression of PD-L1
To explore the molecular mechanism of the effect of M2-Exos on progression of gastric cancer, the authors detected the expression of P38MAPK pathway-related proteins and the immune checkpoint PD-L1 protein. As presented in Figure 7A, protein levels of p-P38 in AGS and HGC27 cells coincubated with M0-Exos were not significantly different from those in the control group. However, it is worth noting that the protein expression of p-P38 was evidently higher in the M2-Exo group than in the M0-Exo and NC groups. In Figure 7B, it can also be observed that PD-L1 protein expression significantly increased in gastric cancer cells coincubated with M2-Exos, indicating that M2-Exos can achieve immune escape by increasing the expression of PD-L1 in gastric cancer cells. In addition to this, the authors have explored other signaling pathways that play an important role in cancer progression, such as the PTEN/AKT signaling pathway and Ras-ERK1/2 signaling pathway. However, there were no significant differences in the proteins involved (Fig. 7C).
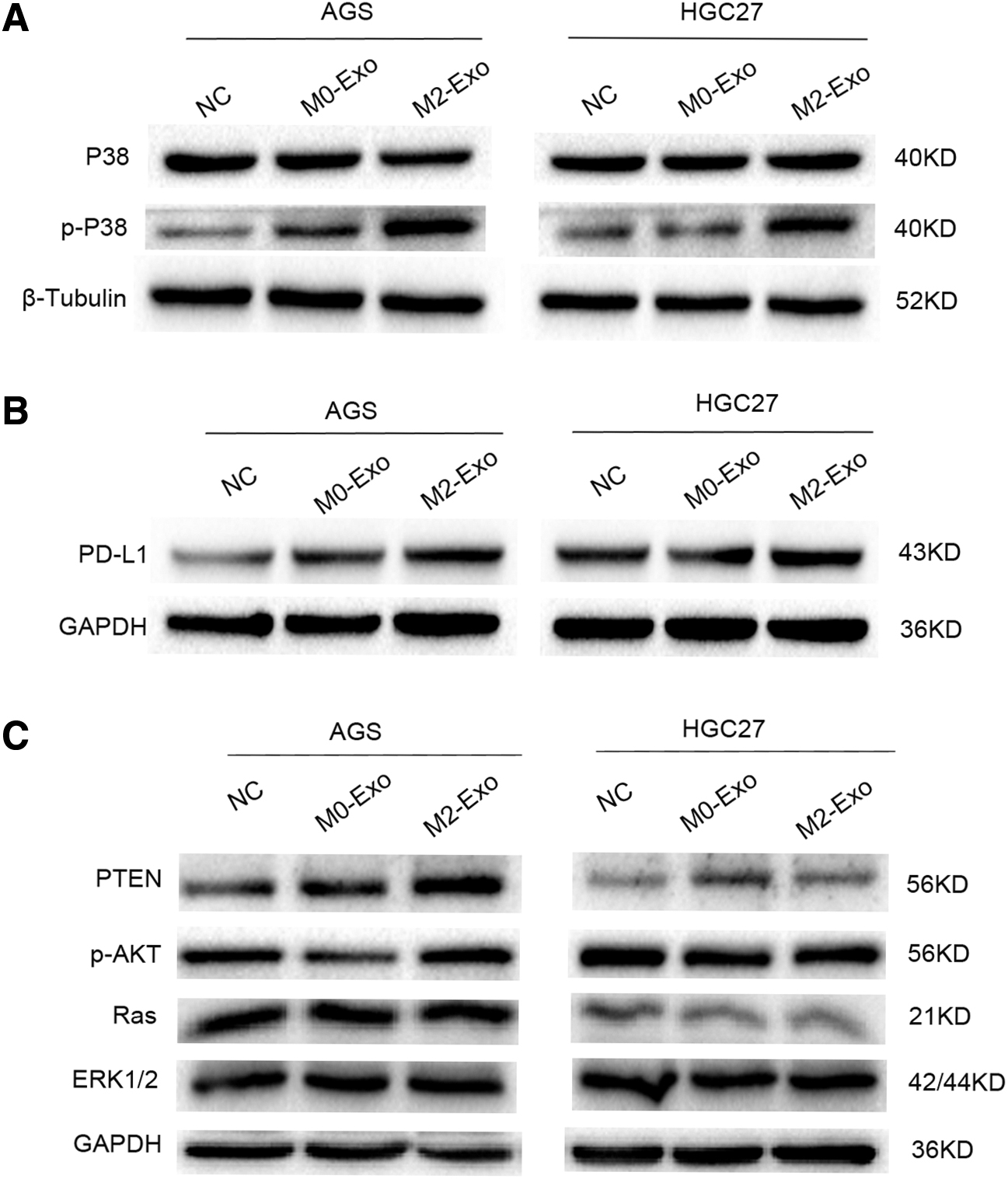
M2-Exos accelerate the progression of gastric cancer by activating the P38MAPK pathway and upregulating the expression of PD-L1.
Discussion
TAMs are major players in the TME and coordinate various factors to promote tumor progression. 14 Typically, macrophages can be polarized into M1-type or M2-type macrophages. M1-type macrophages are activated by cytokines such as IFNG and produce proinflammatory and immune-stimulating cytokines. 8 However, TAMs are thought to be very similar to M2-type macrophages, which are activated by Th2 cytokines. 15 In this study, the authors obtained M2 macrophages by polarization induced by IL-4 and IL-13. A previous study has shown that M2 macrophages express relatively high levels of CD163, IL-10, and CD206. 16 Consistently, in this study, CD206, Arg-1, and IL-10 were found to be highly expressed, indicating that M2 macrophages were successfully induced.
Exosomes are small vesicles containing complex RNA and proteins, typically 40–100 nm in diameter and with a round shape. 17 Similarly, the exosomes that the authors extracted were found to be small and round vesicles with diameters of 50–90 nm. The exosomal membrane is rich in the four-transmembrane protein family, heat shock protein family, and some specific proteins involved in exosomal transport. Among them, CD63, CD9, CD81, TSG101, and HSP70 are the most commonly used exosome markers. 18 In the current study, the authors were pleasantly surprised to find that the extracted exosomes were positive for CD63, CD9, and TSG101, which was consistent with previous studies. Collectively, these data indicated that the exosomes that were extracted are of good purity and biological activity.
Many cells can secrete exosomes under normal and pathological conditions, which can exist in various body fluids and microenvironments, and regulate the biological activity of receptor cells through proteins, nucleic acids, lipids, and so on and participate in the signal transmission between cells. 19 In this study, evidence of fluorescent labeling showed that M2-Exos could be well absorbed and internalized by gastric cancer cells, which laid a foundation for the role of M2-Exos in progression of gastric cancer.
As tumors progress, tumor macrophages have been reported to promote tumor cell proliferation, invasion, and metastasis; stimulate tumor-associated angiogenesis; and inhibit antitumor immune responses. 20 Besides, increasing evidence has revealed that exosomes act as mediators of cell–cell communication within the TME. 21 For example, Lan et al. suggested that M2-Exos accelerated the cell migration and invasion potential in colon cancer. 22 Wu et al. demonstrated that M2-Exos enhanced the motor ability, invasion potential, and metastasis of hepatocellular carcinoma cells. 23 In this study, to explore the effect of M2-Exos on the behavior of gastric cancer cells, the authors examined proliferation, migration, and apoptosis. Similarly, the authors found that M2-Exos accelerated progression of gastric cancer by promoting cell proliferation and migration activity and simultaneously inhibiting gastric cancer cell apoptosis.
GW4869 is a cell-permeable, noncompetitive N-Smase inhibitor commonly used to inhibit the production of exosomes. In the study by Zheng et al., 24 they demonstrated that treatment of M2 macrophages with GW4869 blocked exosome formation. In addition, M2 macrophages failed to promote GC cell migration after GW4869 treatment. In the study by Chen et al., 25 data showed that the coculture system of GW4869 effectively prevented M2 macrophages from regulating collagen synthesis, cell proliferation, and fibroblast invasion. Given that existing data have confirmed that GW4869 can inhibit the release of exosomes and thus affect the behavior of target cells, this research did not conduct a deeper functional inhibition analysis, which is also one of the limitations. In addition, in this study, the authors found that although M0 had no significant effect on gastric cancer cell proliferation and apoptosis, it inhibited cell migration. Therefore, the authors further searched for more information about the role of M0-Exos in cancer. In the study by Zhang et al., 26 data indicate that macrophage-derived exosomes enhanced osteosarcoma cell migration through the miR-29a/NFIA axis, which is different from the conclusions in this study. Given the complexity of biological organisms, the authors speculate that M0-Exos may play different roles in different cancers and the mechanisms involved are also different.
Mitogen-activated protein kinase (MAPK) is a group of serine–threonine protein kinases that can be activated by different extracellular stimuli, such as cytokines, neurotransmitters, hormones, cellular stress, and cellular adhesion. It can transmit extracellular signals into the cell and participate in cell proliferation, differentiation, canceration, metastasis, and apoptosis. 27 P38 is one of the major proteins in the MAPK family, and P38 MAPK is the main pathway by which MAPK regulates the physiological response of cells. When P38 MAPK is stimulated, it forms the stress MAPK signaling pathway, which regulates the progression of cancer by influencing changes of related genes. 28 In the study by Schmieder et al., 29 they describe a novel MS4A8A subset of M2-like TAMs that promote tumor growth and are induced by synergistic activation of P38 MAPK- and GR-dependent signaling pathways. In the study by Kubala et al., LRP1 promoted polarization of M2 macrophages by activating P38MAPK and NF-κB. 30 These data suggest that the P38MAPK pathway plays an important role in M2 macrophages. Exosomes play an important role in cell signal transduction. However, the role of exosomes extracted from M2 macrophages in gastric cancer has been rarely described. Therefore, based on the above background, the authors focus the research on mechanisms of P38MAPK. Interestingly, Western blot data revealed that the protein level of p-P38 was significantly increased in AGS and HGC27 cells coincubated with M2-Exos, suggesting that M2-Exos may promote progression of gastric cancer by activating the P38MAPK signaling pathway.
With the development of immune medicine, it has become increasingly clear that certain cytokines, which were originally described as immune stimulatory and proinflammatory, can also upregulate immunosuppressive molecules. These molecules are key elements of the immunosuppressive pathway, known as immune checkpoints, and they are critical for maintaining self-tolerance and regulating the strength of the immune response. 31 PD-L1 is a 40-kDa transmembrane glycoprotein that negatively regulates the T cell response and plays an important role in controlling the immune response. 32 In addition, PD-L1 expression contributes to tumor escape from immune surveillance and is associated with poor prognosis in cancer patients. 33 For example, Hino et al. showed that the high expression of PD-L1 promoted the vertical growth of malignant melanoma and is an independent prognostic factor. 32 Hamanishi et al. demonstrated that PD-L1 promoted the development of human ovarian cancer. 34
In the current study, the authors found that the PD-L1 protein was significantly increased in cells coincubated with M2-Exos, which was consistent with previous studies. As one of the most promising immune therapy strategies, the PD-L1 inhibitor is a breakthrough for the therapy of some refractory tumors. 35 Deng et al.'s data show that MPDL3280A is a human monoclonal antibody that targets PD-L1 and exerts antitumor activity mainly by blocking PD-L1 interaction with programmed cell death-1 (PD-1) and B7.1. It was regarded as a potential therapy for locally advanced or metastatic malignancies. 36 Next, the immunosuppressive function also be added to reveal the effect of PD-L1 expression in gastric cancer cells induced by M2-Exos. In the study by Noh et al., PD-L1 was proved to be induced by the P38MAPK signaling pathway. 37 Bazhin et al. also confirmed that P38MAPK mediated PD-L1 expression. 31 Similarly, in this study, the authors found that protein expression of both p-P38 and PD-L1 was significantly increased in cells coincubated with M2-Exos. Therefore, the authors speculated that M2-Exos may promote progression of gastric cancer by activating the P38MAPK signaling pathway and elevate the expression of PD-L to achieve immune escape.
Conclusions
In this study, the authors successfully obtained stable M2-Exos and confirmed that M2-Exos promoted proliferation and migration and inhibited the apoptosis of gastric cancer cells. Mechanically, M2-Exos may promote progression of gastric cancer by activating the P38MAPK signaling pathway and simultaneously promote the high expression of PD-L1 in gastric cancer cells to assist gastric cancer cells to achieve immune escape. There are some limitations to this study. First, in the pathway exploration, the authors did not further confirm the involvement of the pathway by adding P38MAPK pathway inhibitors. Second, in immune regulation, the authors did not add PD-L1 inhibitors for further confirmation. Therefore, the next research direction is to verify the regulatory mechanism by adding inhibitors. Furthermore, the authors will continue to explore further downstream targets to enrich the understanding of molecular mechanisms.
Footnotes
Acknowledgment
The authors are thankful to the Gastroenterology Laboratory of Capital Medical University-Affiliated Beijing Friendship Hospital for their guidance.
Funding Information
This work was supported by the Capital Health Research and Development of Special Project (No. 2018-2-2022).
Disclosure Statement
The authors have no financial conflicts of interest.