
Abstract
Objective:
Sonic Hedgehog (Shh)-Gli1 signaling and osteopontin (OPN) play vital roles in pancreatic cancer. However, the precise mechanisms of both signals have not been fully clarified, and whether there is a correlation between them in pancreatic ductal adenocarcinoma (PDAC) is unknown. This study aims to confirm the effect of OPN on human PDAC and assess whether Hh signaling affects pancreatic cancer cells through upregulation of OPN.
Materials and Methods:
OPN expression in human PDAC tissues and cell lines was investigated. Proliferation, apoptosis, migration, and invasion of OPN-knockdown BxPC-3 cells were observed. We analyzed the correlation between Shh or Gli1 and OPN expression in human PDAC. Hh signaling inhibitors and shRNA against Gli1 were used to confirm if OPN expression in BxPC-3 cells was regulated by Hh canonical or noncanonical pathway. We also evaluated the proliferation, apoptosis, migration, and invasion of Gli1-knockdown BxPC-3 cells.
Results:
OPN is highly expressed in human PDAC tissues and cell lines. The proliferation, migration, and invasion of BxPC-3 cell lines were decreased, whereas apoptosis was increased when OPN was knocked down. Correlation analysis showed that Gli1, but not Shh, was associated with OPN expression in human PDAC, and Gli1 regulated OPN production in BxPC-3 cells through a noncanonical pathway because Gli but not Smo inhibitor reduced OPN expression. Similar to above, the proliferation, migration, and invasion of BxPC-3 cells were decreased, whereas the apoptosis was increased when Gli1 was knocked down. Supplement of exogenous OPN protein could partially reverse the effect of both OPN knockdown and Gli1 knockdown on the bio-behavior of BxPC-3 cells.
Conclusion:
Hh signaling promotes proliferation, migration, and invasion but inhibits apoptosis of pancreatic cancer cells through upregulation of OPN in a noncanonical pathway.
Keywords
Introduction
Pancreatic cancer, the majority type of which is pancreatic ductal adenocarcinoma (PDAC), is one of the most malignant human tumors worldwide and presents a very poor prognosis. 1 Over the past three decades, despite improvements in therapeutic techniques, the 5-year overall survival rate only increased from 5% to 6%. 2 There are many causes of the poor outcome of PDAC, such as difficulties in early detection, high relapse rate after surgery, chemo- and radiotherapy resistance, and a high tendency of metastasis.2–4 At present, certain germline or acquired gene mutations have been observed in PDAC, such as KRAS (90%), CDK2NA (90%), TP53 (75%–90%), DPC4/SMAD4 (50%), and so on. However, no apparent progress on targeted therapy against these mutations has been made yet because of the complexity of the signals and microenvironments involved in PDAC.5–7 Although researchers keep attempting newer immunotherapies, such as anti-PD-1 and CAR T cells, to improve the efficacy, they are also less effective.8–12 In addition to optimizing treatment, seeking more effective biomarkers of PDAC to enhance early detection is also a strategy to improve prognosis. Thus, it is essential to further clarify the mechanism of pancreatic cancer and the involved molecules.
The Hedgehog (Hh) signaling pathway, which plays a critical role in embryo development, has been strongly associated with pancreatic cancer incidence. Three Hh ligands have been found in vertebrates: Sonic Hedgehog (Shh), Indian Hedgehog (Ihh), and Desert Hedgehog (Dhh), among which Shh was intensively reported to play a role in malignancies including PDAC. 13 In a canonical Shh pathway, Shh protein binds to its membrane receptor patched 1 (Ptch1), leading to attenuation of the inhibitory effect on the coupled coreceptor smoothened (Smo). Activation of Smo, in turn, activates the transcription factor glioma-associated oncogene (Gli). The Gli family includes Gli1, Gli2, and Gli3. The nuclear translocation of Gli2 and Gli3 caused by the activation of Smo induces the expression of Gli1. 14 Gli1 promotes transcription of its target genes, such as PTCH1, Cyclin D1, c-Myc, and Bcl-2. 14 As Gli1 itself is a target gene of Hh signaling, its expression level can indicate the activation degree of the Hh signaling pathway.
Compared with the quiescent state in normal adult pancreatic tissues, the Hh signaling is highly active in malignant pancreatic tissues and its precursor lesions, pancreatic intraepithelial neoplasia (PanIN). 15 Overexpression of Shh leads to PanIN in mice, and Smo inhibitor can reverse this alternation.15,16 However, in human beings, the involvement of Shh and Smo in pancreatic cancer is controversial, but Gli1 does play an essential role by regulating its target genes to mediate tumor bio-behaviors. 17 It should be a noncanonical Hh pathway, unlike the one mentioned previously. However, the precise mechanism of the Shh pathway in human pancreatic cancer, including the upstream signals and downstream targets of Gli1, remains unclear and needs further clarification.
Osteopontin (OPN) is an integrin-binding ligand. After binding with integrin molecules or/and CD44, it mediates the adhesion of cells and extracellular matrix (ECM), 18 regulates cell proliferation and survival, and participates in various pathological changes, such as chronic inflammation, 19 tumorigenesis, and tumor development and metastasis.20–22 OPN has been upregulated in >30 types of malignant tumors, including pancreatic cancer. 20 It is secreted by macrophages or other cells in the tumor microenvironment23,24 and cancer cells.24,25 The high expression of OPN in pancreatic cancer was confirmed in 2002 by global gene sequence. 26 Some reports has indicated that it is a potential biomarker of pancreatic cancer and mediates tumor advance.25,27,28 A recent study has reported that OPN enhances growth, 29 metastasis,29,30 and chemoresistance of pancreatic cancer cells. 31 However, the precise role of OPN in pancreatic cancer has not been fully clarified, or has the cause of its high expression been confirmed.
According to the reports, Hh signaling and OPN play vital roles in pancreatic cancer. Of note, OPN has been reported to be downstream of Hh signaling. Shh secreted by alveolar type II cells induces the production of OPN by macrophages, which promotes pulmonary fibrosis to progress. 32 In melanoma, OPN, upregulated by Gli1, promotes the malignant behavior of cancer cells. 33 However, there are no reports that expression of OPN is upregulated by canonical or noncanonical Hh pathway in pancreatic cancer. Hence, in this study, we intended to explore whether activation of the Hh signaling influences the bio-behavior of pancreatic cancer cells through upregulation of OPN. We investigated the role of OPN in proliferation, apoptosis, migration, and invasion of pancreatic cancer cells and identified if OPN was a downstream molecule of Shh. We demonstrated that OPN affected these bio-behaviors mentioned previously and was upregulated by Gli1 through a noncanonical Hh pathway. Our study further elucidated the molecular signaling mechanisms that mediated pancreatic cancer and may provide some new clinical diagnosis and treatment ideas.
Materials and Methods
Patients and specimens
Fifty human PDAC tissues were collected from the Department of General Surgery, Affiliated Hospital of Jiangsu University (Zhenjiang, Jiangsu Province, China). We removed all the samples between January 2017 and December 2020. Among the PDAC patients, there were 32 men and 18 women, with ages ranging from 28 to 81 years, average age being 61.64 years. All the PDAC patients had not undergone chemo- or radiotherapy or else before surgical resection. Samples were fixed with formalin, and we performed pathological diagnosis after the paraffin section. All the tissue samples were obtained with the patient's informed consent before being used for the present research. The Ethics Committee of Affiliated Hospital of Jiangsu University approved our study (Approval no. KYLS20190205).
Immunohistochemical analysis
Paraffin-embedded samples were sectioned into 4-μm-thick specimens and were deparaffinized in xylene and rehydrated by a series of 5-min washes in 100%, 95%, and 80% ethanol and distilled water. Then sections were treated with 0.3% Triton X-100 at room temperature for 30 min. Endogenous peroxidase was blocked in 3% hydrogen peroxide for 30 min. Next, all samples were autoclaved in citric acid buffer (pH 6.0) and 120°C for 3 min for antigen retrieval. After blocked with 5% bovine serum albumin (BSA) at room temperature for 1 h, samples were incubated with primary rabbit anti-human OPN (Abcam; ab8448, 1:500), rabbit anti-human Gli1 (Abcam; ab17326, 1:500), or rabbit anti-human Shh (Boster; BA2171, 1:200) at 4°C overnight, and then with biotinylated goat anti-rabbit IgG (Boster; BA1003, 1:500) at room temperature for 1 h, followed by treatment with avidin–biotin–peroxidase (Boster; SA1022) complex for 45 min. Immunostaining was developed by a DAB kit (Boster; AR1027). Between each of the steps mentioned previously, slides were washed three times with PBS for 5 min. Finally, the sections were counterstained with hematoxylin, dehydrated, cleared, and mounted. The structures exhibiting brown staining in the tissues were considered positive.
Cell culture and treatment with drugs
The pancreatic cancer cell lines, BxPC-3, PANC-1, and SW1990, were kindly provided by Prof Min Xu (Affiliated Hospital of Jiangsu University, Zhenjiang, China). 34 BxPC-3 cells were cultured in RPMI-1640 medium (Wisent, Inc.), and PANC-1 and SW1990 cells were cultured in Dulbecco's modified Eagle's medium (DMEM) (Wisent, Inc.). Both medium were supplemented with 10% inactivated fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.), and cells were maintained in 1% penicillin/streptomycin (HyClone, GE Health Care) at 37°C with 5% CO2. BxPC-3 cells were seeded into six-well plates at a density of 106/well, and cyclopamine 35 (10 μM; MedChemExpress LLC) or GANT6136 (20 μM; MedChemExpress LLC.) were added into the corresponding well. DMSO, the solvent of the drugs, was kept at a constant concentration in both the test and control cultures. We purchased recombinant human OPN protein from R&D Systems, Inc., and BxPC-3 cells were treated as previously described 36 to explore whether the effects caused by OPN or Gli1 knockdown could be reversed by it.
RNA extraction and reverse transcription–quantitative polymerase chain reaction analysis
Total RNA was extracted from cells with TRIzol (Takara) according to the manufacturer's protocol. The cDNA was generated using a reverse transcriptase kit (Promega Corp.). Reverse transcription–quantitative polymerase chain reaction (RT-qPCR) was performed as previously described, 37 and each experiment was conducted in triplicate. The primers used for RT-qPCR are given in Table 1. All the primers were designed using Primer Blast of PubMed.
Primers of Human OPN, Gli1, Shh, and β-Actin for Real-Time–Quantitative Polymerase Chain Reaction

Western blot
Cells were lysed with RIPA buffer (Merck KGaA; Sigma-Aldrich), and a protease inhibitor cocktail containing PMSF, E-64, aprotinin, and Leupeptin (Sangon Biotech) was added. An equal amount of protein sample was loaded and separated by SDS-PAGE (10% gel) and transferred to PVDF membranes. Membranes were blocked with 5% BSA for 2 h at room temperature and immunoblotted overnight at 4°C with primary antibodies against full-length human OPN (AF1433, 1:4000; R&D), Gli1 (ab17326, 1:2000; Abcam), Shh (#2207, 1:1000; Cell Signaling Technology), and anti-β-actin (BM0627, 1:4000; Boster) used to confirm equal loading. The membranes were then incubated with horseradish peroxidase-conjugated secondary antibodies at room temperature for 1 h. The target proteins were subsequently visualized using enhanced chemiluminescence. 38
Plasmid constructs, transfection, and production of a stable cell line
To generate the short hairpin RNA (shRNA) construct against human OPN (sh-OPN), three targeting sequences from the OPN gene were selected, and three corresponding pairs of primers were designed as given in Table 2. The plvx-hOPN-shRNA-Zsgreen-T2A-Puro plasmid was constructed by inserting an annealed short hairpin oligonucleotide targeting OPN into plasmid plvx-shRNA-Zsgreen-T2A-Puro Vector between the EcoR I and BamH I restriction endonuclease sites. The empty plasmid vector (plvx-shRNA-Zsgreen-T2A-Puro) was used as the negative control. According to the manufacturer's protocol, all transfections were performed using the Lipofectamine 2000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.). We used the same method as mentioned previously to construct shRNA targeting human Gli1 (sh-Gli1), and primers for the synthesis of candidate sh-Gli1s are given in Table 2.
The Sequences of the Primers to Create Candidate shRNAs Against the Selected Target Sequences Within hOPN and hGli1 Genes, Respectively

To generate lentiviral particles, HEK 293T cells cultured in 100 mm culture dishes were cotransfected with 15 μg Plvx-shRNA-T2a-puro, 11.25 μg psPAX2, and 3.75 μg pMD2.G. The supernatant was harvested at 48 h post-transfection, and then clarified by filtration through a 0.45-μm membrane filter (EMD Millipore), and concentrated through ultracentrifugation at 50,000 g for 2 h at 4°C (AvantiTM J-30I; Beckman Coulter). Infection was performed by adding 1 mL of the lentiviral supernatant containing 1 × 106 recombinant lentivirus transduction units to BxPC-3 cells in a 60 mm culture dish with 4 mL of growth medium supplemented with 8 μg/mL polybrene (Merck KGaA; Sigma-Aldrich). After 24 h, the viral particle-containing medium was removed and replaced with a fresh medium containing 1 μg/mL puromycin. From day 4, a new medium was replaced when necessary, and we evaluated transfection efficiency under a microscope according to the ratio of Zsgreen-positive cells. Finally, the cells were collected for further experiments.
Cell counting kit-8 assay
The BxPC-3 cells transfected with sh-OPN or sh-Gli1 (or sh-NC as the control) were seeded into 96-well plates with 3 × 103 cells/well, and grown in 200 μL culture medium at 37°C, 5% CO2. Following culture for 20, 44, and 68 h, we added 10 μL of Cell counting kit-8 (CCK-8) solution (Dojindo Molecular Technologies, Inc.) into each well. The cells were cultured for another 4 h, and the absorbance values were analyzed at 450 nm using a microplate reader (Elx800; BioTek Instruments, Inc.). All cell samples were tested in triplicate.
Flow cytometry
The control cells (sh-NC group), OPN knockdown cells (sh-OPN group), and OPN knockdown cells incubated with rhOPN protein (sh-OPN + rhOPN group) were plated in a six-well plate. Following culture for 24 or 48 h, the cells were digested with trypsin (Merck KGaA; Sigma-Aldrich) and washed with PBS twice. The cells were resuspended at a density of 5 × 105/mL with 100 μL 1 × binding buffer and incubated with 5 μL of Alexa Fluor 647-Annexin V (4A Biotech Co. Ltd.) for 10 min in the dark at room temperature, and then 5 μL of propidium iodide (4A Biotech Co. Ltd) was added for 3 min under the same condition. Next, 500 μL of 1 × binding buffer was added into each tube and analyzed by flow cytometry to evaluate the role of OPN in BxPC-3 cell apoptosis. The same procedures were performed to evaluate the apoptosis of BxPC-3 cells that had undergone Gli1 knockdown. The rhOPN was also supplemented to see if exogenous OPN might reverse the effect of Gli1 knockdown on apoptosis, at least in part.
In vitro cell migration and invasion assays
The ability of cells to migrate was tested using scratch wound healing. Cells were seeded into six-well culture plates at 5 × 105 cells/well and cultured until they reached 80%–90% confluence. A 10-μL pipette tip was used to scratch the confluent monolayers in the shape of a straight line, and floating cells were washed with PBS, then the medium was replaced with RPMI-1640 with no FBS. Pictures of the same wound position were taken after 0, 12, 24, and 48 h microscopically.
In addition, transwell insert chambers (Corning, Inc.) were also used to evaluate the migration and invasion of cancer cells. The cells were suspended at a density of 2 × 105/mL in total 150 μL of serum-free RPMI-1640 and added into the upper chamber containing an insert filter with or without matrigel and 800 μL medium containing 10% FBS was placed in the lower chamber. After incubation at 37°C for 36 h, migrated or invaded cells were fixed in 4% paraformaldehyde for 20 min and then washed with PBS. The cells in the upper chambers were removed, and the inserts were stained with crystal violet for 15 min. After washing the sides twice with PBS, the migrated or invaded cells were counted under a contrast microscope at × 10 magnification. The results in the bar diagram represent the mean ± standard deviation (SD) of the number of cells from five randomly selected fields.
Statistical analysis
Data are presented as the mean ± SD from at least three independent experiments. Statistical comparisons of the results between two groups were performed using the Student's t-test, and one-way analysis of variance (ANOVA) was used when samples were more than two groups, except for the CCK-8 assay results, which were analyzed using repeated-measures ANOVA. p < 0.05 was considered as a statistically significant difference. All statistical analysis was performed using SPSS software.
Results
OPN is highly expressed in human pancreatic cancer tissues and cell lines
Before confirming the role of OPN in pancreatic malignancies, the expression of OPN was detected in human pancreatic cancer tissues and cell lines. Immunohistochemistry showed that 70% (35/50) of PDAC tissues strongly expressed OPN, whereas only 32.0% (16/50) of adjacent pancreatic tissues did (Fig. 1A). OPN-positive staining in cancerous tissues was mostly concentrated in cancer cells. Still, there were also positive structures (Fig. 1A, indicated by the triangle), following the idea that OPN is a component of ECM. Expression of OPN by the three PDAC cell lines (BxPC-3, PANC-1, and SW1990) was detected using qPCR and Western blot, and the normal pancreatic ductal cell line hTERT-HPNE (HPNE) was used as the control. As given in Figure 1B, BxPC-3 cells expressed the most abundant OPN mRNA and the other two cancer cell lines both expressed more OPN than HPNE cells. The Western blot result further confirmed that BxPC-3 cells expressed the highest level of OPN (Fig. 1C). These results suggest that OPN is frequently expressed in human pancreatic cancer tissues, especially in the cytoplasm of cancer cells. According to the abundance of expression, BxPC-3 cell line was selected for subsequent assays.
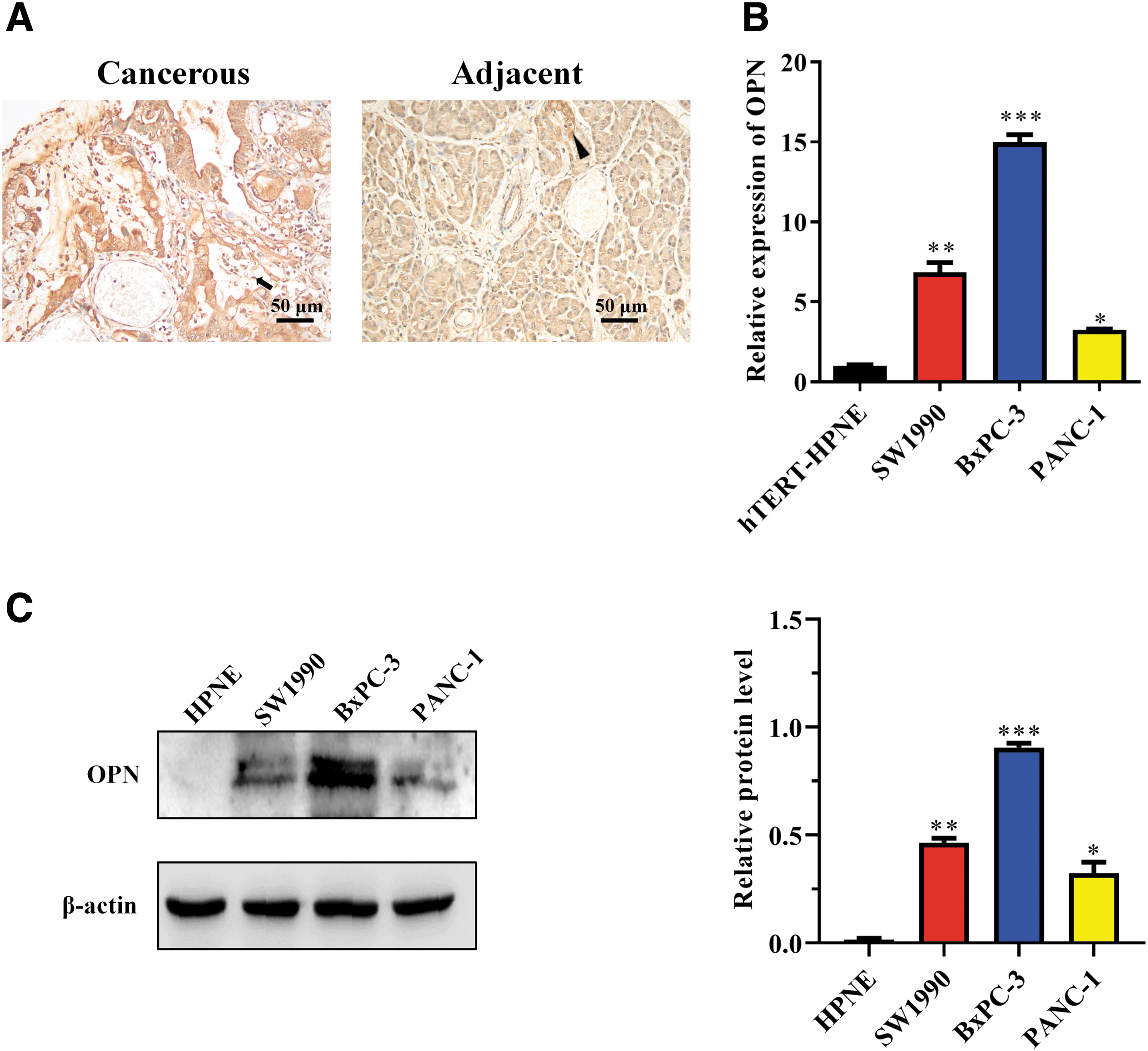
Expression of OPN in human pancreatic cancer tissues and cell lines.
Knockdown of OPN inhibits cell proliferation, migration, and invasion but induces apoptosis of BxPC-3 cells in vitro
To further analyze the functional role of OPN in the biological behaviors of BxPC-3 cells, the three candidate shRNAs targeting the human OPN gene (sh-OPN) were transfected into the cell line using a lentivirus vector, respectively. The empty vector (sh-NC) was used as the control. PCR showed all the shRNAs reduced expression of OPN, among which sh-OPN-2 reduced the most (81.2%, ***p < 0.001, compared with sh-NC), and sh-OPN-1 and sh-OPN-3 reduced by 42.2% (**p < 0.01, compared with sh-NC) and 75.9% (***p < 0.001, compared with sh-NC), respectively (Fig. 2A). Expression of OPN protein demonstrated by Western blot further confirmed the result of qPCR (Fig. 2B). Thus, sh-OPN-2 was used in our study to interfere with OPN expression by BxPC-3 cells, and the “sh-OPN” undermentioned was sh-OPN-2.
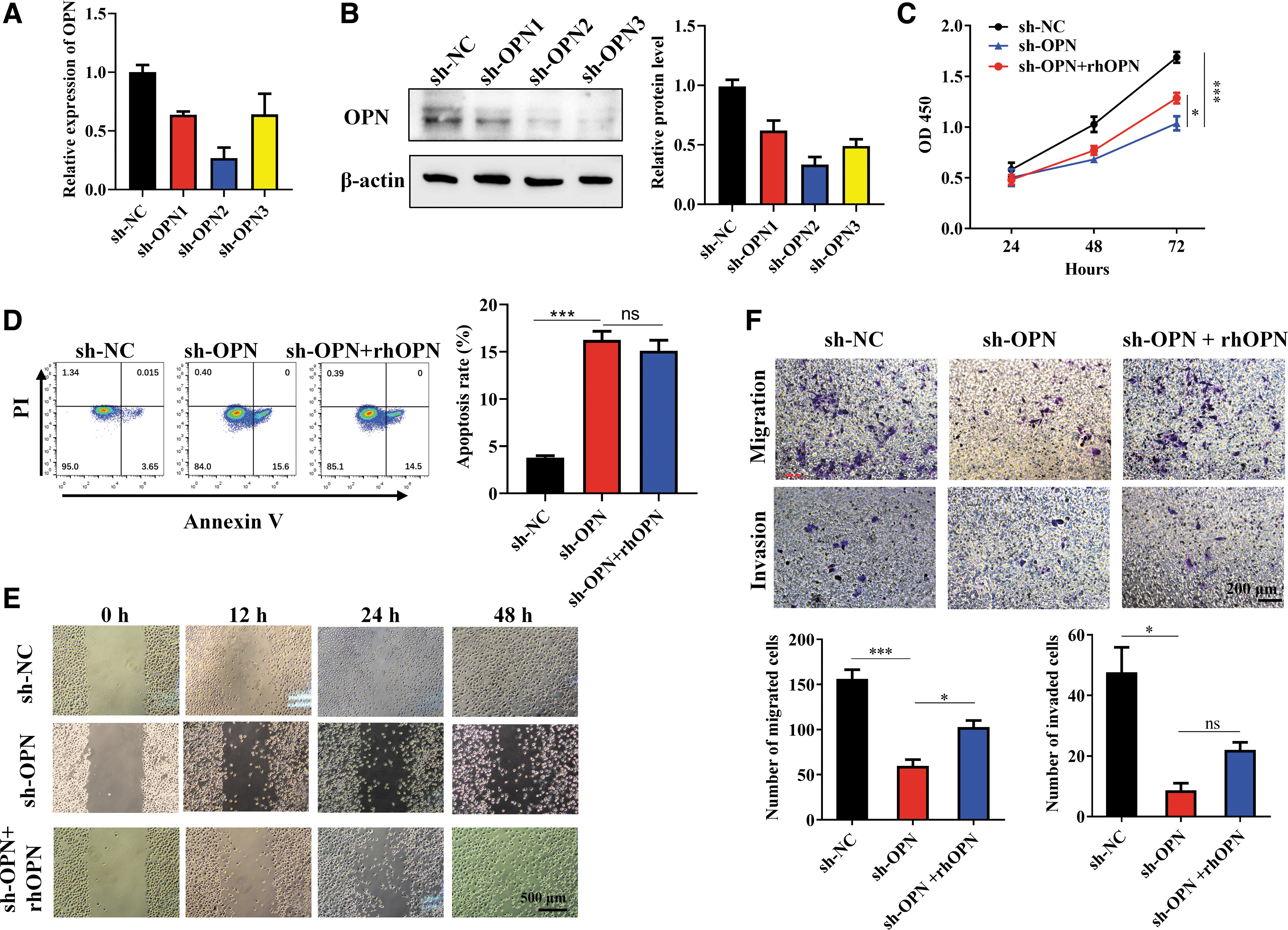
Knockdown of OPN led to slowed proliferation, promoted apoptosis, and inhibited migration and invasion of BxPC-3 cells. BxPC-3 cells were transfected with the three candidate shRNAs targeting OPN (sh-OPNs), and effect of OPN knockdown on bio-behaviors of BxPC-3 cells were tested.
Knockdown of OPN using sh-OPN led to multiple changes in BxPC-3 cell bio-behaviors. As indicated by the CCK-8 assay, OPN knocked-down BxPC-3 cells proliferated more slowly than the sh-NC group (***p < 0.001; Fig. 2C), and apoptosis of BxPC-3 cells improved after OPN expression was inhibited (***p < 0.001), as given in Figure 2D. Wound healing assay showed that at both 24 and 48 h after scratching, the cells in sh-OPN group migrated less than those in the sh-NC group (Fig. 2E). Transwell assay further confirmed, as demonstrated by Figure 2F (the upper panel), that sh-OPN transfected BxPC-3 cells migrated much less than the control cells after 24 h (***p < 0.001). Similarly, the invasion of BxPC-3 cells was also reduced when OPN expression was knocked down (*p < 0.05; the lower panel of Fig. 2F).
A recent study reported that BxPC-3 cells express integrins such as αVβ3, αVβ5, and αVβ6, 39 and αVβ3 and αVβ5 are known OPN receptors. 40 Therefore, exogenous rhOPN (200 ng/mL) was used to stimulate the OPN-knocked-down BxPC-3 cells further to validate the role of OPN in the above bio-behaviors. As given in Figure 2C, rhOPN partially restored the reduced proliferation (*p < 0.05), and the inhibited cell migration also improved in part (*p < 0.05; Fig. 2E, F). However, no significant changes were seen in the apoptosis and invasion of OPN knocked-down BxPC-3 cells treated with rhOPN (Fig. 2D, F), suggesting that the two behaviors mediated by OPN may not be achieved by binding the integrin receptors.
On the whole, these findings suggest that OPN produced by pancreatic cancer cells may play an essential role in promoting the growth, survival, and progression of PDAC.
OPN expression is positively correlated with Hh signaling in human pancreatic cancer
It has previously been shown that OPN expression is regulated by Shh signaling, which is considered an essential pathway in pancreatic cancer tumorigenesis and progression. 32 Thus, this study postulated that the high expression of OPN in pancreatic cancer cells is caused by activation of the Shh signaling. To validate the correlations of expression of Shh and OPN and Gli1 (the main terminal effector of Hh signaling) and OPN in human pancreatic cancers were analyzed by the GEPIA database. The correlation between IL-6 and OPN was also examined because reports suggest that IL-6 is an OPN inducer in some malignancies or nontumor diseases, such as head and neck cancer, osteosarcoma, hyper IgE syndrome,41–43 and so on. Spearman correlation analysis showed that among Shh, Gli1, and IL-6, only Gli1 was positively correlated with OPN expression in human pancreatic cancer tissues (R = 0.27, p = 0.00031; Fig. 3A). This result follows Gu's review. The authors commented that canonical (ligand-dependent) Hh signaling seems not activated in the tumor compartment of PDAC, but Gli does play a role. 44 In addition, the possibility of IL-6 as a regulator of OPN was excluded here owing to the correlation analysis.

OPN expression was related to activation of the Hh signaling in human PDAC.
Next, coexpression of Shh, Gli1, and OPN in human PDAC tissues was observed using immunohistochemistry. We found that 52.0% (26/50) and 64.0% (32/50) of the cases expressed Shh and Gli1, respectively. Shh was concentrated in the cytoplasm of cancer cells, whereas Gli1 was distributed in both cytoplasm and nucleus (Fig. 3B), suggesting the nuclear translocation of this transcript factor. Among the cases, 54.0% (27/50) had coexpression of Gli1 and OPN, and 38.0% (19/50) had coexpression of Shh and OPN. About 32.0% (16/50) expressed all the three molecules (Fig. 3B). However, considering the results of correlation analysis and Gu's review, 44 it is likely that OPN expression in human PDAC is not related to Shh but Gli1.
We further investigated the expression of Shh and Gli1 in BxPC-3 cell line that highly expresses OPN. qPCR indicated that compared with HPNE cells, BxPC-3 cells expressed a higher mRNA levels of Shh (*p < 0.05; Fig. 3C) and Gli1 (*p < 0.01; Fig. 3D). Western blot further confirmed that BxPC-3 cells expressed Shh and Gli1 protein levels more than HPNE cells did (Fig. 3E). Because the expression of Gli1 is indicative of Hh signaling activity, 45 this result indicates that in BxPC-3 cell line, Hh signaling is activated, and the ligand leading to expression of Gli1 might be Shh.
These findings suggest that OPN expression in human pancreatic cancer is associated with activation of the Hh signaling.
OPN expression in BxPC-3 cells is regulated by Gli1, but not Shh ligand
To evaluate whether OPN expression is regulated by Hh signaling in pancreatic cancer cells, Hh inhibitors were used at first. In a classical Shh signaling, the Shh ligand binds to the membrane receptor Ptch1, leading to activation of Smo, and Smo, in turn, activates Gli through an intracellular process. It was demonstrated by qPCR and Western blot that OPN expression in BxPC-3 cells was not affected by treatment with cyclopamine, a Smo inhibitor, at both mRNA and protein levels (Fig. 4A, B), suggesting that the classical Shh signaling pathway was not involved in OPN expression. In contrast, expression of OPN was significantly decreased by treatment with GANT61 (Fig. 4A, B), a Gli inhibitor that alters the conformation of the Gli molecule and inhibits its binding to DNA. These data suggest that OPN expression in BxPC-3 cells is regulated by Gli, which is not induced by ligand binding but a noncanonical Hh pathway.
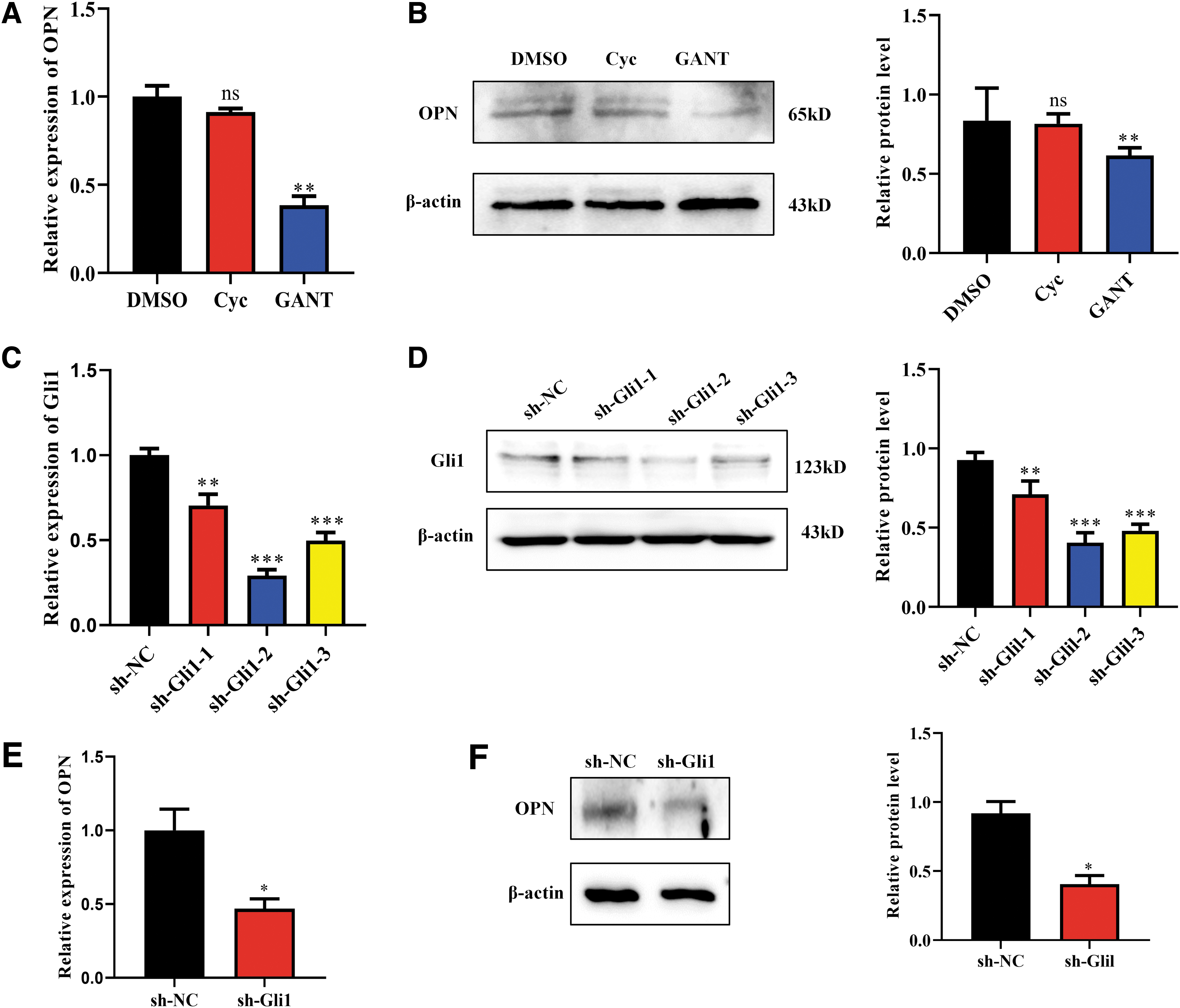
OPN expression by BxPC-3 cells is regulated by Gli1 but not Shh. Hh signaling inhibitors cyclopamine (Cyc) or GANT61 (GANT) were used to treat BxPC-3 cells (the solvent DMSO was used as the control), and Gli1 knockdown BxPC-3 cells were constructed using shRNA.
To further confirm whether Gli1 induces OPN expression, three candidate lentiviral vector-mediated sh-RNAs against Gli1 (sh-Gli1-1, sh-Gli1-2, and sh-Gli1-3) were transfected into BxPC-3 cells, respectively. Both Gli1 mRNA (Fig. 4C) and protein (Fig. 4D) expression was reduced by the sh-Gli1-1/2/3 and sh-Gli1-2 reduced Gli1 more (70.8%, ***p < 0.001 vs. sh-NC group) than sh-Gli1-1 and -3 (29.6%, **p < 0.01 and 50.2%, ***p < 0.001, respectively, both vs. sh-NC group). Therefore, sh-Gli1-2 was selected as the sh-Gli1 used in our subsequent experiments. After Gli1 was knocked down, OPN mRNA (Fig. 4E) and protein (Fig. 4F) levels in BxPC-3 cells both decreased, as shown by qPCR and Western blot. It is further validated that Gli1 is an inducer of OPN expression in this pancreatic cancer cell line.
Gli1 affects the bio-behaviors of pancreatic cancer cells by regulating OPN expression
We then evaluated whether knockdown of Gli1 would influence the bio-behaviors of BxPC-3 cells, which was demonstrated above to be regulated by OPN, and rhOPN was supplemented to see if the effects would be reversed. CCK-8 indicated that proliferation decreased after sh-Gli1 transfection (***p < 0.001 vs. sh-NC; Fig. 5A) but increased in part by rhOPN treatment (*p < 0.05 vs. sh-Gli1; Fig. 5A). Flow cytometry showed an improved early apoptosis rate when Gli1 was knocked down. Identically, rhOPN did not reduce such improved apoptosis (Fig. 5B), suggesting a nonparacrine or autocrine manner with which OPN mediated apoptosis of BxPC-3 cells. Wound healing was inhibited by knockdown of Gli1, and rhOPN rescued such inhibition, as given in Figure 5C. Transwell assay indicated that migration and invasion decreased when Gli1 was knocked down, as given in Figure 5D (***p < 0.001, *p < 0.05, ns, no significance, both compared with sh-Gli1).
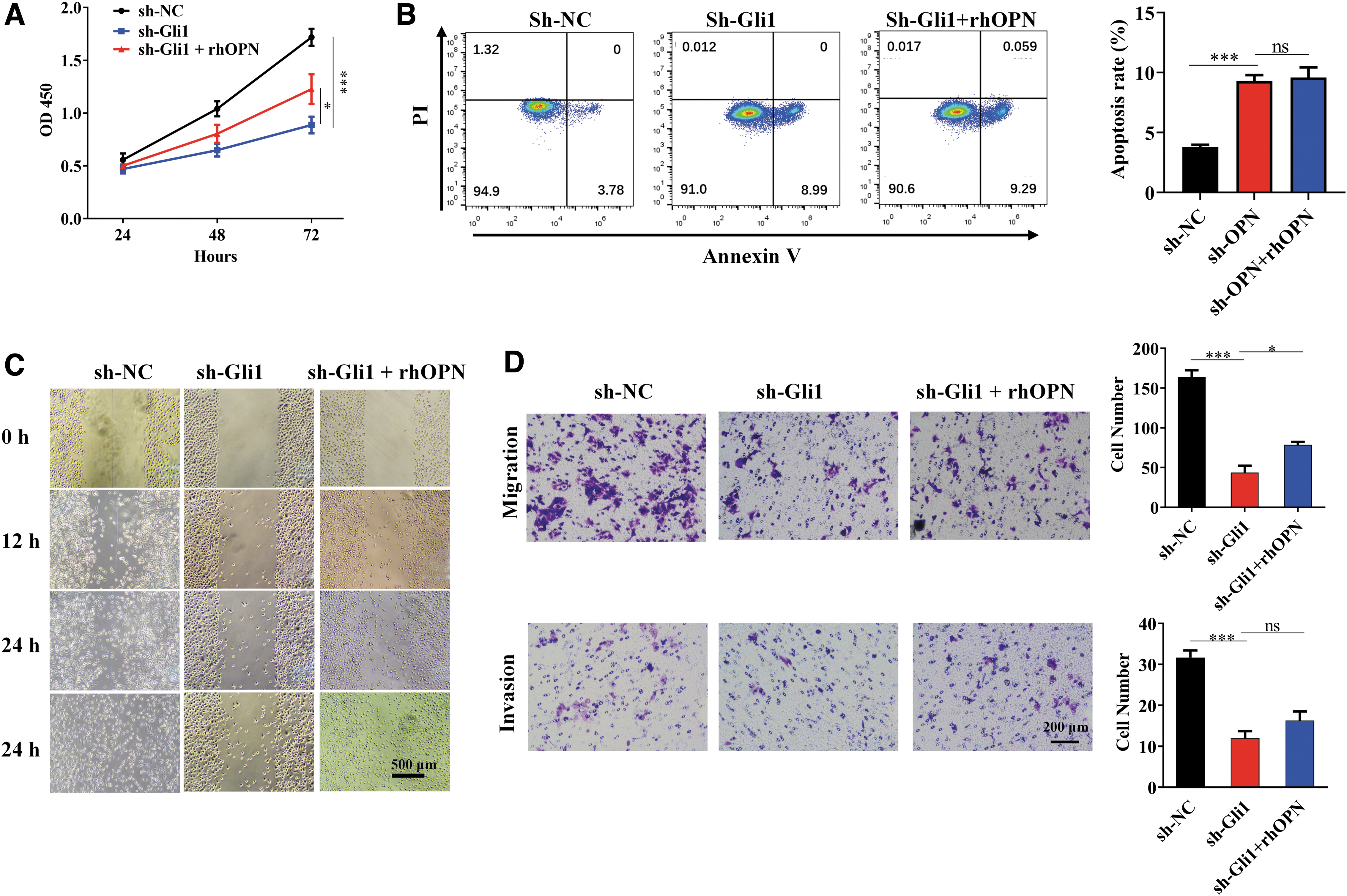
Knocking down of Gli1 led to bio-behavior changes that were also affected by OPN of BxPC-3 cells. Bxpc-3 cells were transfected with sh-Gli1 and cultured with or without rhOPN (sh-Gli1+rhOPN and sh-Gli1, respectively). Cells transfected with an empty vector and cultured with no rhOPN were the control (sh-NC).
Overall, these findings suggest that Gli1 promotes proliferation, migration, and invasion but inhibits apoptosis of BxPC-3 pancreatic cancer cells. The role in proliferation and migration depends on the regulation of OPN expression, at least in part. The effect on apoptosis and invasion of BxPC-3 may be also mediated by upregulated OPN.
Discussion
Pancreatic cancer is an aggressive malignancy with a poor prognosis. Shh was reported to play an essential role in tumorigenesis and the progression of PDAC.46–48 Still, unlike the efficacy in mice models,48,49 Smo inhibitor had not improved prognosis obviously in human cases13,50 because of limitation due to harmful and potentially toxic side-effects 13 or/and ineffectiveness against nonclassical Hh pathways in the disease. However, Gli proteins (especially Gli1 51 ), the downstream transcription factors of Hh ligands, play a vital role in human PDAC, 44 but its downstream signaling is not yet fully elucidated.
OPN was reported to be involved in tumor bio-behaviors, including cell proliferation, angiogenesis, metastasis, chemoresistance, and so on, 52 and was consistently overexpressed by pancreatic tumor patients. It was considered as a potential biomarker of PDAC. 53 However, the precise role and the cause of its high expression in human pancreatic cancer are not fully clarified. Reports have indicated that OPN is a downstream molecule of the Hh signaling pathway.32,33 Given that Hh and OPN are both involved in human PDAC, the correlation between them was investigated in our study.
Our results revealed that OPN overexpressed in human pancreatic cancer tissues and three PDAC cell lines, BxPC-3, SW1990, and PANC-1. Among the cell lines, BxPC-3 expressed the most abundant OPN. In recent years, OPN has been considered as a tumor metastasis-related molecule because many data have focused on its role in mediating cancer metastasis.54–59 BxPC-3 is derived from a primary PDAC tissue with no metastasis, PANC-1 is also from a primary focus, but the patient had local lymph node metastasis, and SW1990 is entirely from splenic metastatic tissues. Thus, OPN expression by the cell lines seemed discrepant from the idea that OPN is a tumor metastasis mediator. We speculated that OPN in the early stage of pancreatic cancer played a role in the genesis and growth of tumor cells and inhibited apoptosis rather than mediates metastasis. Our experimental results also confirmed that OPN could promote proliferation and inhibit apoptosis of BxPC-3 cells. In addition, metastasized tumor tissues may not necessarily overexpress metastatic markers because after settling, it can undergo a mesenchymal–epithelial transition process opposite EMT. 60
Next, we knocked-down OPN expression in BxPC-3 cells using shRNA, which led to slower proliferation, increased apoptosis, and less migration and invasion, suggesting that OPN does play a role in these bio-behaviors of the PDAC cells. In addition, a supplement of exogenous rhOPN protein partially reversed the reduction of proliferation and migration caused by OPN knockdown, indicating that OPN can affect BxPC-3 cells in an autocrine or paracrine way. However, apoptosis was not influenced by rhOPN. It may be because OPN mediates apoptosis not by binding to its integrin receptors on the cell membrane but by an intracellular mechanism, and the precise mechanism remains to be further studied. After OPN knockdown, the number of invaded cells that underwent rhOPN treatment was more than without rhOPN, but there was no significant difference. The cause may be similar to that of apoptosis. Still, as a primary PDAC cell line, the ability of BxPC-3 cells to invade is weak, so the influence of OPN on invasion is probably but not very obvious.
Hence, our study proved that OPN plays a promoting role in the growth, survival, migration, and invasion of pancreatic cancer. However, a study by Collins et al. performed microarray analysis on 245 resected pancreatic cancer specimens and showed that OPN was significantly overexpressed in the cancer tissues of the patients (p = 0.004). Still, subsequent follow-up showed that patients with higher serum OPN level had longer survival. 61 This result contradicts many reports suggesting OPN as a tumor promoter. We made further analysis regarding this. First, the results of Collins were all followed-up patients who had undergone surgical resection of cancer tissues. At the same time, other studies included cases who could not undergo surgery, so this result does not apply to all issues. Second, the effect of OPN on the pancreas may not be straightforward. OPN secreted by cancer cells in vivo promotes tumor growth, and OPN in the tumor microenvironment also has specific functions. In our study, we found that cancer cells are likely to be the primary source of OPN. However, after tumor resection, OPN should be mainly produced by microenvironmental immune cells, epithelial cells, and so on. It is also worth discussing whether cells are secreting a high level of OPN in the process of wound repair after surgery. Therefore, future studies should focus more on the differences in the effects of OPN from cancer cells and microenvironment on human PDAC to clarify further the exact mechanisms of OPN's impact on the disease.
We then investigated if the high expression of OPN in human pancreatic cancer is caused by activation of Hh signaling. Spearman correlation analysis showed the expression of Gli1, but not Shh is related to OPN. Immunohistochemistry demonstrated that Gli1 coexpressed OPN-positive tumor cases were a bit more than Shh coexpressed ones. Of note, in all the Gli1-positive cases, Gli1 localizes in both cytoplasm and the nuclei of tumor cells, suggesting the nuclear translocation of this transcript factor. As for BxPC-3 cell line, Shh and Gli1 are both expressed, but OPN expression was only downregulated when cells were treated with Gli inhibitor but not Smo inhibitor, suggesting OPN was regulated by a noncanonical Hh signaling rather than a canonical one triggered by Smo activation following Hh protein binding. Thus, it seems strange that Shh expressed in BxPC-3 cells does not affect OPN expression, but its terminal effector Gli1 does. HPNE, a regular pancreatic duct epithelial cell line, also represents a low level of Shh but hardly expresses Gli1. The possible reason may be that Shh cannot activate Hh pathway until it reaches a certain concentration threshold (according to Fig. 3E, the expression level of Shh in BxPC-3 cell line is not high), or some molecule downstream of Shh that is involved in activating Gli1 is not expressed in such cell lines. Thus, in BxPC-3 cells, expression of Gli1 that regulating OPN production may be induced by other factors rather than Shh, which can be regarded as a noncanonical Hh signaling pathway.
So long as Gli1 was suggested to play a more crucial role in OPN expression than Shh, shRNA targeting Gli1 was constructed and transfected into BxPC-3 cells using a lentivirus vector. The shRNA successfully inhibiting Gli1 expression, migration, and invasion in BxPC-3 cells were reduced significantly, but apoptosis was increased. Such effects of sh-Gli1 on BxPC-3 cells were consistent with those of sh-OPN described previously. Moreover, treatment of rhOPN rescued the reduced proliferation and migration in part, further confirming that Gli1 could mediate these behaviors through OPN. However, rhOPN hardly influenced the apoptosis of Gli1 knockdown cells. It also proved that OPN inhibition of tumor cell apoptosis was not achieved through the integrin receptor on the surface. Similarly, the effect of rhOPN on the invasion of Gli1 knocked-down BxPC-3 cells was not significant, although invaded cells were more than the group with no rhOPN. The reason may be the same as apoptosis, but it may also be owing to the weak invasive capability of the cancer cell line, and the effect of OPN supplementation is not apparent.
Overall, this study suggested that the pancreatic cancer cells are regulated by Gli1, an effector molecule of Hh signaling, to produce OPN, which facilitates cell growth and endows migration and invasion of cancer cells. This study may also prove that OPN functions as a diagnostic and therapeutic target of pancreatic cancer.
Footnotes
Funding Information
This work was supported by Graduate Innovation Program of Jiangsu University (Grant No. CXLX12_0675), Jiangsu Provincial Youth Talent (Grant No. QNRCQ2016839) and Young Scientists of Affiliated Hospital of Jiangsu University (Grant No. JDFYRC2016002).
Disclosure Statement
No competing financial interests exist.