
Abstract
Background:
Pancreatic adenocarcinoma (PAAD) is a type of malignant tumors in the digestive tract. It is extremely aggressive. However, the molecular mechanism of the occurrence and development of pancreatic cancer has not yet been elucidated. New evidence shows that the dysregulation of small nucleolar RNAs (SnoRNAs) plays an important role in tumorigenesis and has a certain connection with tumor stem cells. In this study, the authors screened differentially expressed SnoRNAs in pancreatic cancer, further explored whether the HGF/C-Met pathway is involved in the regulation of SNORD35A in pancreatic cancer stem cells.
Materials and Methods:
AffymetrixmiRNA 4.0 and QRT-PCR was used for differential screening of SnoRNA. CCK8, wound healing assay and TransWell chamber were used to detect cell proliferation, migration and invasion. QRT-PCR was used to detect the changes of epithelial - mesenchymal Transition (EMT) related genes of tumors. The authors detected the expression levels of HGF/C-Met pathway and its related proteins by Western blotting.
Result:
The authors found that SNORD35A is significantly overexpressed in pancreatic cancer. After disturbing the expression of SNORD35A, the epithelial markers increased and the mesenchymal markers decreased during the EMT process. At the same time, down-regulation of SNORD35A inhibited the proliferation, migration and invasion of pancreatic cancer stem cells in cellular level. In nude mouse transplanted tumor models, low expression of SNORD35A reduced tumor growth volume and attenuated its pathological features. Finally, the authors found that silencing SNORD35A reduced the expression levels of C-Met and its phosphorylated proteins.
Conclusion:
These results suggest that the regulation of SNORD35A on proliferation, migration, invasion and EMT of pancreatic cancer stem cells involves HGF/C-Met signaling pathway. SNORD35A has carcinogenic effects in pancreatic cancer and may become a prognostic biomarker and therapeutic target for pancreatic cancer patients.
Introduction
Pancreatic cancer represents only 3% of all cancers in the United States but accounts for 7% of all cancer-related deaths. 1 Pancreatic cancer has one of the worst prognoses of all tumors, with a 5-year survival rate after diagnosis of only 10%. 2 Many studies have demonstrated abnormal regulation of specific small nucleolar RNAs (snoRNAs) in certain cancer cells. 3 For example, snoRNA 113-1 inhibits hepatocellular tumorigenesis 4 ; the abnormally high expression of snoU2_19, SNORD72, and ACA11 facilitate the proliferation and invasion of liver cancer cells. 5,6 NOP10 can predict the prognosis of lung cancer, and its related SnoRNAs SNORA65, 7A and 7B promote the proliferation and migration of lung cancer cells. 7 Kitagawa et al. found that SNORA74A and SNORA25 are beneficial for the early detection of pancreatic cancer. 8 The initial oncogenic mutations in many cancers often occur in stem cells.
In 1997, Dick and Bonnet of the University of Toronto isolated leukemia cancer stem cells (CD34++ CD38−) for the first time. 9 Recent findings have elaborated that SNORD89 and SNORA72 facilitated cell stemness through the Notch1/c-Myc pathway. 10,11 Moreover, the HGF/C-Met pathway is associated with the occurrence of cancer stem cells. Therapies targeting C-Met have also attracted wide attention. The study found that STMN1 regulates tumor stem cell properties in vitro through HGF/Met signaling and slows hepatocellular tumor growth. 12 And inhibition of C-Met reversed the malignant potential of pancreatic cancer cells. 13 However, conventional therapies for pancreatic cancer target tumor cells with limited tumorigenic potential, rather than the cancer stem cell population. The role of snoRNAs in pancreatic cancer stem cells (PCSCs) has not been adequately studied. It is unclear whether snoRNAs affect PCSCs through the c-MET signaling pathway.
The authors previously demonstrated the presence of PCSCs with CD44, CD24, and epithelial specific antigen (ESA) expression on their cell surfaces. 14 On this basis, they report the effects of U35A in pancreatic cancer for the first time. In this study, AffymetrixmiRNA 4.0 and quantitative (q) PCR were used to screen out significantly dysregulated snoRNAs in pancreatic cancer. High expression of U35A promoted epithelial–mesenchymal transition (EMT) in PCSCs and changed the relative phosphorylation levels of C-Met and Erk in the HGF/C-Met signaling pathway. In addition, inhibiting U35A significantly reduced tumor development in vitro and in vivo. Therefore, U35A has profound significance to the prognosis and treatment of pancreatic adenocarcinoma (PAAD).
Materials and Methods
Patients and clinical specimens
To screen and investigate snoRNAs that were associated with pancreatic cancer, the authors obtained surgical specimens from pancreatic cancer patients who were treated at Shanghai Ninth People's Hospital (Detailed information is available in Table 1). This study was approved by the Ethics Committee of Science and Technology, Shanghai University.
Clinical Characteristics of Pancreatic Cancer Patients
Tumorigenicity assays
Male BALB/C nude mice (6–8 week old) were purchased from Shanghai Jiesijie Experimental Animal laboratory (Shanghai, China). All animal experiments were approved in advance by the Animal Protection Department strictly follow Shanghai University's Experimental Animal Institution Care and Use Plan. Male BALB/C nude mice were divided into different groups and injected with 1 × 105 cells that had been transfected with U35A-shRNA. The authors detected tumor growth in the mice after 28 d, tumor metastasis was observed in nude mice using small animal imager. The mice were euthanatized using pentobarbital (Sigma-Aldrich, St. Louis, MO). Tumor volumes were calculated using the formula: V = width × width × length × 0.52. 15
Cell culture
All cell lines (PL-45, Hpde6-C7, and Bxpc-3) were acquired from ATCC. Cells were cultured in DMEM supplemented with 10% FBS and RPMI 1640 medium, including 10% FBS, respectively.
SnoRNA profiling
The authors used surgically resected tumor tissues from patients with pancreatic cancer to construct tumor xenograft models 16 using single cell suspensions of tumors to generate the xenografted mice. As in Li et al., the antibodies used included anti-CD44 (APC, PE, or biotin), anti-CD24 (PE or FITC), anti-B38.1 (APC), and (ESA)-FITC (Biomeda, Foster City, CA). The authors used flow cytometry to isolate pancreatic cancer cells that were positive or negative for all three markers (CD44+CD24+ESA+ and CD24−CD44−ESA−). To identify differentially expressed snoRNAs in tumor tissues, total RNA was isolated and analyzed using the GeneChip™ miRNA 4.0 Array (Affymetrix, Shbio, Shanghai). The normalized pancreatic ductal epithelial cells were used as negative controls.
Quantitative PCR
U35A expression was identified in PCSCs using real-time SYBR green quantitative PCR to test for changes in U35A that had been treated with RNA interference. Then the authors observed the differential expression of related genes during EMT in the PCSCs. According to the manufacturer's recommendations, total RNA was extracted from PCSCs using the TransZol Up protocol (Qiagen, TransGen Biotech, China). The authors used a multiplate reader (Thermo Fisher Scientific, Waltham, MA) to explore the concentrations of RNA (or DNA) using the A260 nm/A280 nm ratio.
The HifairII 1st Strand cDNA Synthesis Kit and Hieff UNICON qPCR SYBR Green Master Mix (Yeasen BioTechnologies Co. Ltd.) were used for reverse transcription PCR. All PCR tests were performed using a IQ5 Multicolor Real-Time PCR exploration system (Bio-Rad) and quantified through the 2−ΔΔCt method. GAPDH was used as a control for normalization in each sample. Stem-loop primers were designed to reverse transcribe snoRNAs and increase the specificity of the assay. The stem-loop primer sequences are listed in Table 2. The primer sequences for EMT-related genes are listed in Table 3.
The Stem-Loop Primers for Small Nucleolar RNAs
The Primer Sequences for Measuring Expression of Each Gene by Reverse Transcription-Quantitative Polymerase Chain Reaction
Transfection
The authors designed a 19-nucleotide special siRNA that could target 5′-GAAUGGCGACAAUGCCAAUTT-3′ in the sequence of U35A (NCBI gene ID: 26816), which was synthesized as the corresponding interfering sequences. Control oligo RNA and siRNA for U35A were purchased from Shanghai GenePharma. Transfections were performed using Opti-MEM medium and Lipofectamine 2000 in accordance with the manufacturer's protocol. The short hairpin (SHRNA) sequences were designed using the LV3 (H1/GFP and Puro) or LV16 (U6/Luciferase17 and Puro) vector systems (GenePharma) and were used to establish stable Bxpc-3 and PL-45 or mutant Bxpc-3 and PL-45 cell lines. Vector plasmid maps are shown in Supplementary Figure S1. Bxpc-3 and PL-45 cells were cultured and used for shRNA transfection (dilution ratio 1:1000). All cell lines were cultured in the corresponding medium with 10% FBS (Gibco, Grand Island, NY). After 72 h, 4 μg/mL puromycin (YEASEN) was added to the stable cell lines for the next 7 d.
Proliferation assay
Cell proliferation assays were used to assess cell viability and cell proliferation. The authors used the Cell Counting Kit-8 (CCK-8; Sangon Biotech) in accordance with the supplied instructions to determine cell proliferation. In brief, 100 μL of a cell suspension was dispensed in 96-well plates and incubated for 24 h in a humidified incubator. Then, 10 μL of CCK-8 solution was added to each well for 20 min. Finally, absorbance at 450 nm was measured using a microplate reader.
Migration and invasion assays
Transwell chambers were used to evaluate the migration and invasion ability of tumor cells. Under serum-free conditions, cells transfected with U35A shRNA were added in the upper well of a transwell chamber (Nunc, China) containing 8-μm pore size of polycarbonate filters for the cell migration assay. Using Corning® BioCoat™ Matrigel® Invasion Chamber (Corning, NY) for the invasion test that also included an 8-μm pore size PET membrane with a thin layer of MATRIGEL basement membrane matrix. Into the lower chamber was added 500 μL of medium containing 15% FBS, and then the cells were cultured at 37°C with 5% CO2 for 24 h. After incubation, a cotton swab was used to wipe the nonmigrated cells from the upper surface of the membrane. Then, 100% methanol and 1% crystal violet were used to fixed and stain the cells, respectively. The invading cells were then observed and photographed under a microscope, and then cells in several fields of three membranes were counted.
Wound healing assay
Cells were cultured in six-well plates, and then transfected at a certain density. Cells were scraped with a pipette tip at different timepoints and washed with PBS. Migration rates were calculated using the following formula: wound healing area (%) = (initial scratch distance − final cell-free imaging distance) ÷ initial scratch distance.
Western blotting assay
Western blotting was performed using standard protocols. The antibodies used were as follows: phospho-p44/42 MAPK (Erk1/2), p44/42 MAPK (Erk1/2), phospho-Akt, Akt, phospho-Met, Met, phospho-Stat3, Stat3, and β-actin. All antibodies were purchased from Cell Signaling Technology (Danvers, MA).
Statistical analysis
All statistical analyses were informed by three separate replicate tests, and data are shown as mean ± SEM. One-way analysis of variance (ANOVA) and the Student's t-test were used for data analysis. p < 0.05 was statistically significant.
Results
U35A was overexpressed in PAAD
To evaluate the differential expression of genes in pancreatic cancer tissues, the authors sorted CD24+CD44+ESA+ PCSCs and CD24−CD44−ESA− pancreatic cancer cells by flow cytometry from a mouse PDX model of pancreatic cancer. AffymetrixmiRNA 4.0 was used for snoRNA screening (Fig. 1A). Compared with normal pancreatic ductal epithelial cells, there are 29 upregulated and 12 downregulated snoRNAs in CD24+CD44+ESA+ PCSCs. The authors found nine snoRNAs, including SNORD14B, SNORD32A, U35A, SNORD35B, SNORD58B, SNORD83A, SNORD97, and SNORD101, which showed significant changes. The most downregulated gene was SNORD14B and the most upregulated gene was U35A. Quantitative PCR was then used to verify the expression of the snoRNAs (Fig. 1B). Consistent with the snoRNA screening, the expression of U35A was significantly increased.
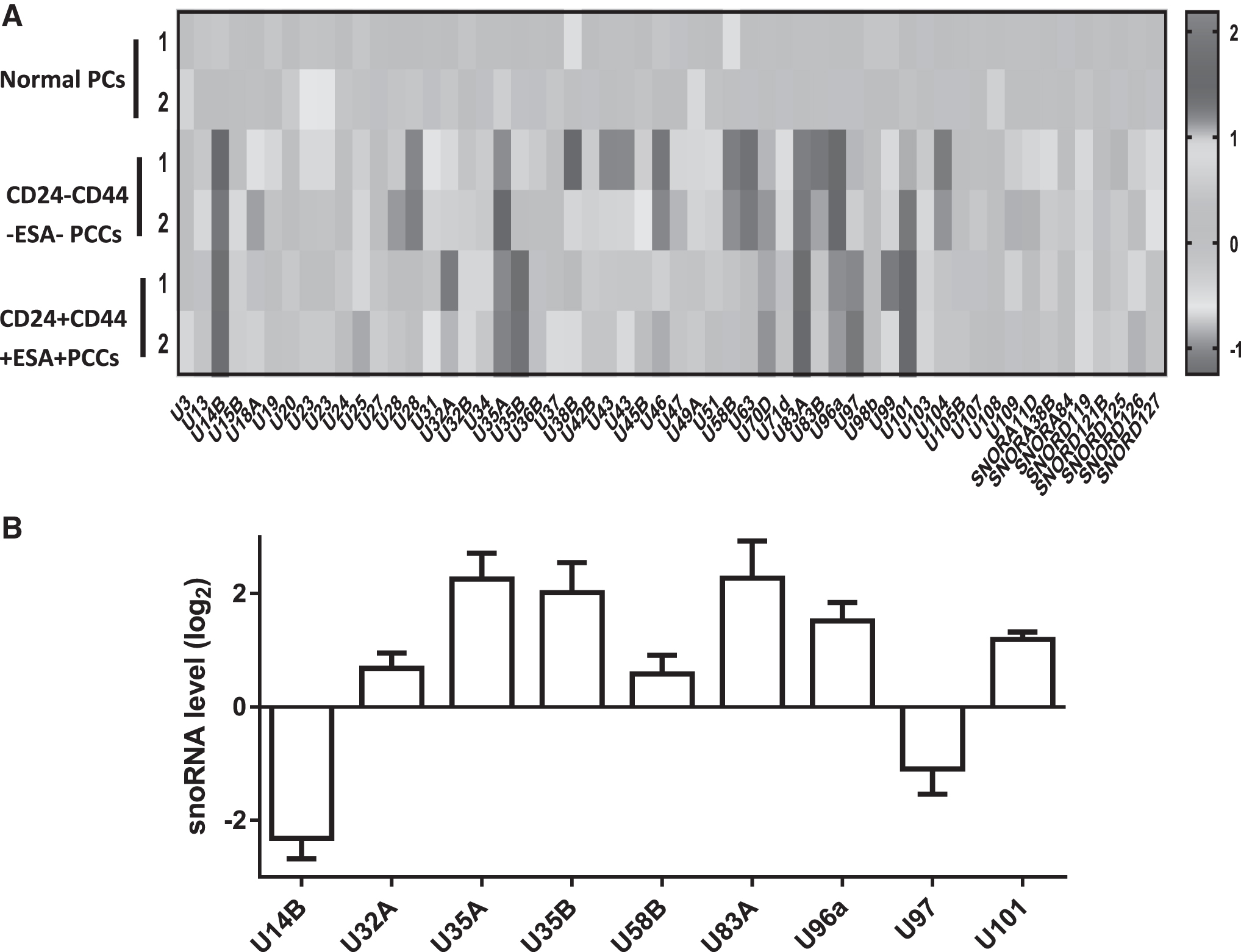
Differential expression of SnoRNAs in PAAD.
U35A promoted PAAD tumorigenesis in vitro and in vivo
The authors next transfected PCSCs (Fig. 2A) and compared the proliferation levels of Bxpc-3 and PL-45 cells after transfection with Si-U35A. CCK-8 results indicated that cells treated with U35A-siRNA had significantly decreased viability compared with cells transfected with scrambled siRNA (Fig. 2B). Subsequently, the authors explored whether U35A affected the migration and invasion in PCSCs. In BXPC-3 and PL-45 cells, they used transwell assays to evaluate cell migration, which was inhibited after transfecting with Si-U35A (Fig. 2C). Meanwhile, they further verified the migration of PCSCs by wound healing assays. Similar to the transwell assay results, Bxpc-3 cells transfected with Si-U35A showed no significant differences from the control group at 0 h; however, after 24 h, there was increased wound closure in the control group compared with the Si-U35A group.

SNORD35A promoted tumorigenesis in vivo.
Scratches in the control group showed complete closure after 48 or 72 h. The similar results were seen in PL-45 cells (Fig. 2D, E). In addition, Matrigel invasion assays were performed to determine the effects of U35A on the invasive behavior of PCSCs. The invasion ability of Bxpc-3 and PL-45 cells were significantly reduced by downregulating U35A (Fig. 2F). In summary, these results suggested that silencing U35A suppressed the growth of PAAD stem cells.
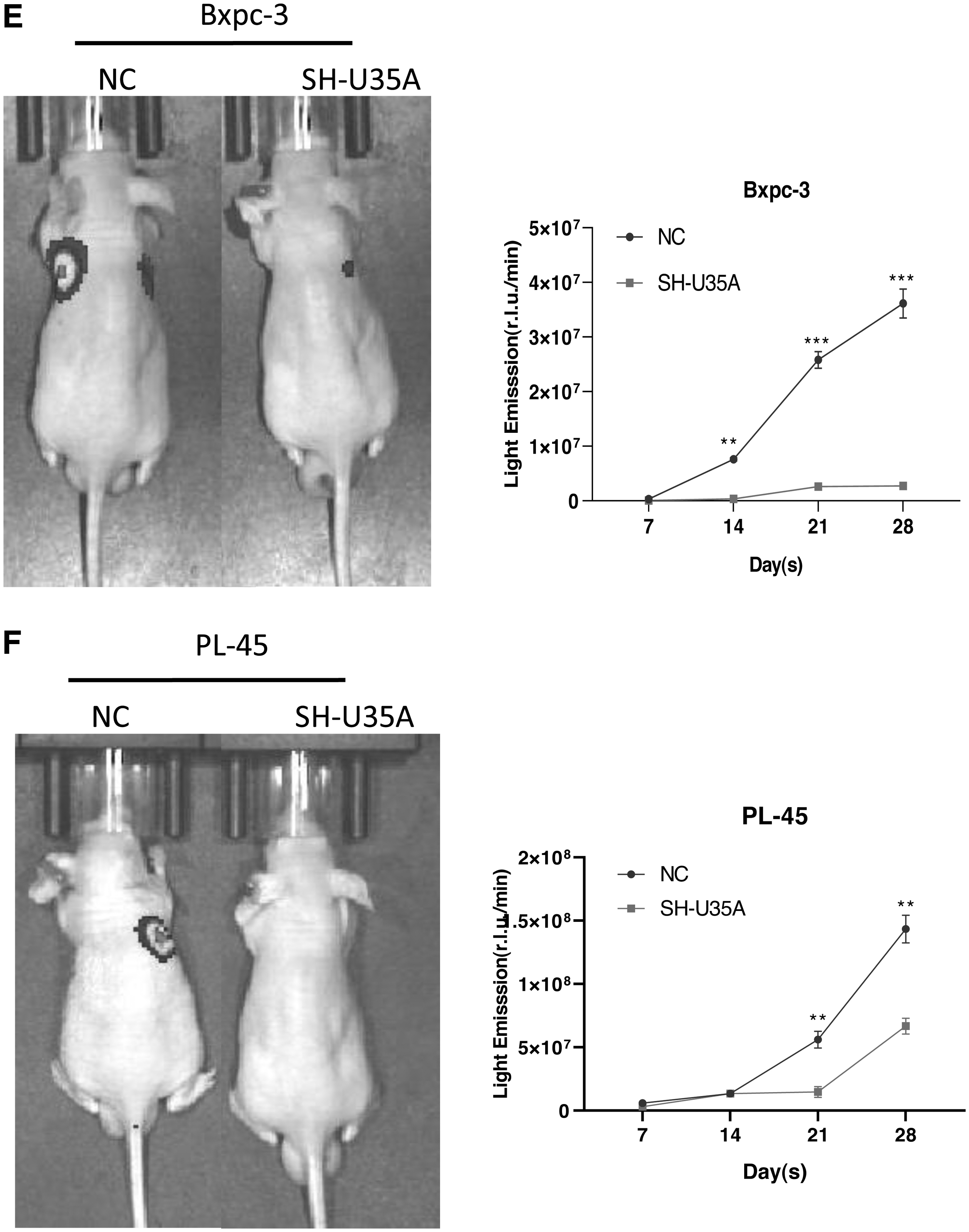
Next, the authors constructed a stable transfection system of SH-U35A (Fig. 3A), and established xenograft mouse models through the subcutaneous injection of PCSCs into postnatal 6–8 week old mice. Compared with the group treated with scrambled SHRNA, Bxpc-3 cells transfected with U35A-SHRNA led to significantly reduced tumor volumes after 30 d. Meanwhile, with the passing of time, the weight of the nude mice also showed a downward trend. The authors observed the similar situation using PL-45 cells (Fig. 3B, C). Compared with the NC group, tumor tissue transfected with SH-U35A showed no obvious tumor atypia and reduced tumor vacuoles (Fig. 3D). The results of tail vein in nude mice showed that the ability of tumor metastasis was decreased after transfection with SH-U35A (Fig. 3E, F). In summary, these results indicated that downregulating U35A can significantly inhibit tumorigenesis in PAAD.
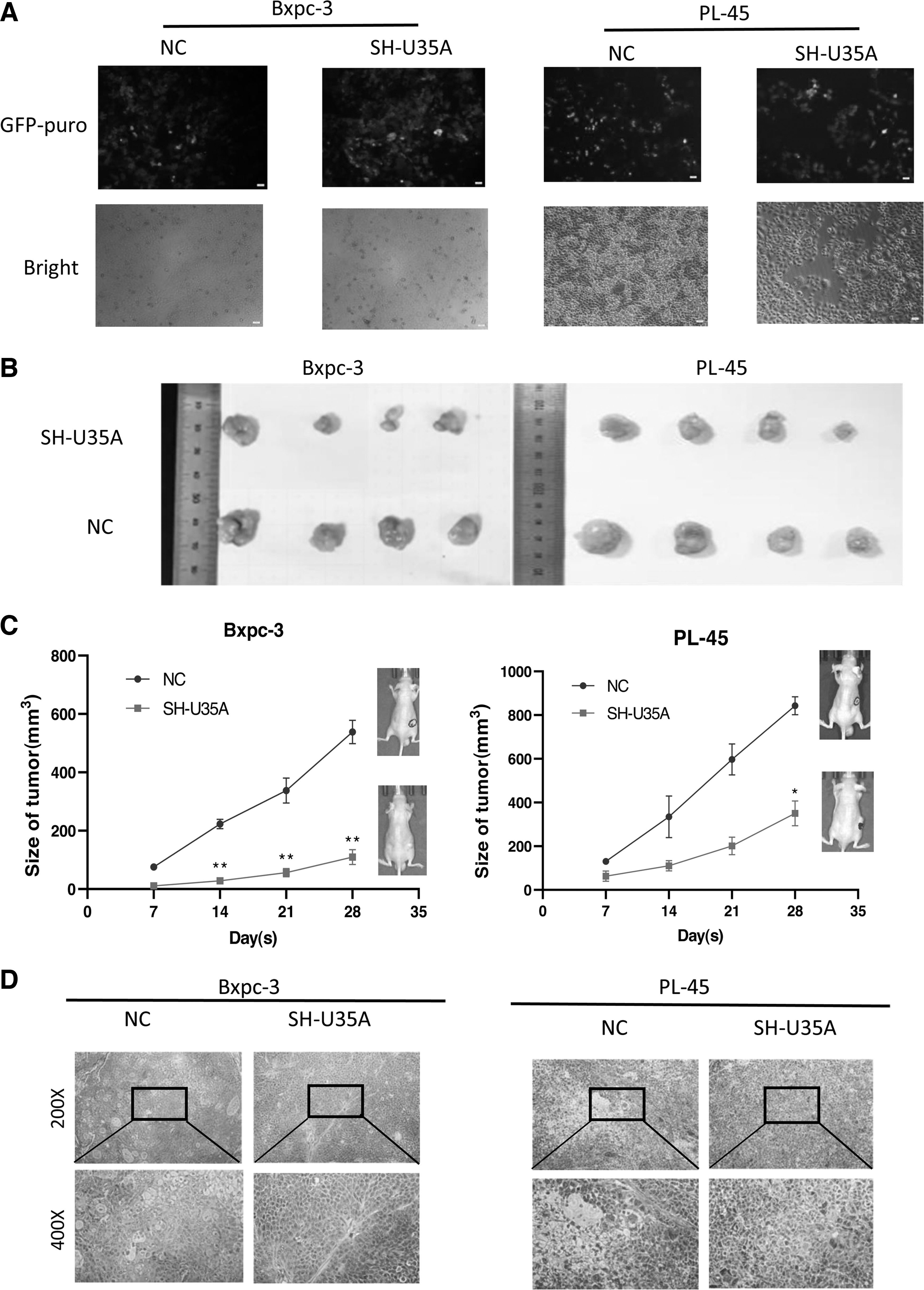
SNORD35A promoted tumorigenesis in vitro.
U35A promoted EMT in PCSCs
The expression levels of snoRNAs in PCSCs extracted from patient tumor tissues were significantly upregulated, indicating that these may play key roles in tumorigenesis. EMT can promote the migration and invasion of cancer cells by enabling cancerous epithelial cells to transform into a mesenchymal-like cell. Therefore, the authors further performed quantitative PCR to demonstrate the levels of EMT-related markers. The data showed that N-cadherin (CDH2) and the interstitial marker Vimentin (VIM) were decreased in PCSCs transfected with si-U35A. In addition, tomoregulin-1 (TMEFF1), Snail Homolog2 (SNAI2), and fibronectin1 (FN1) expression levels suggested a sharp decrease in Bxpc-3 cells, while the levels of matrix metalloproteinase 2 (MMP2), SNAI2, FN1, and TMEFF1 were significantly reduced (Fig. 4A); meanwhile, the expression levels of Occludin (OCLN) were increased in PL-45 cells (Fig. 4B). These results indicated that decreased U35A expression could suppress EMT in PCSCs.

Analyses of EMT-related genes in pancreatic cancer. After transfecting BXPC-3
U35A affects PCSCs by preventing C-Met signaling
HGF/C-Met signaling plays an important role in the growth and migration of cells. Owing to the binding of HGF and C-Met, Met tyrosine kinase is activated, and tyrosine residues in Met undergo autophosphorylation, which in turn induces the development of tumor cells. To explore the roles of U35A in PCSCs, the authors used Western blotting to detect C-Met expression levels. As shown in Figure 5A–C, si-U35A treatment markedly decreased C-Met phosphorylation. Meanwhile, they observed the expression of key molecules in other related signaling pathways, such as P44/42 MAPK (ERK1/2) of the MAPK pathway, Stat3 of the JAK/Stat pathway, and Akt of the PI3K/Akt pathway.
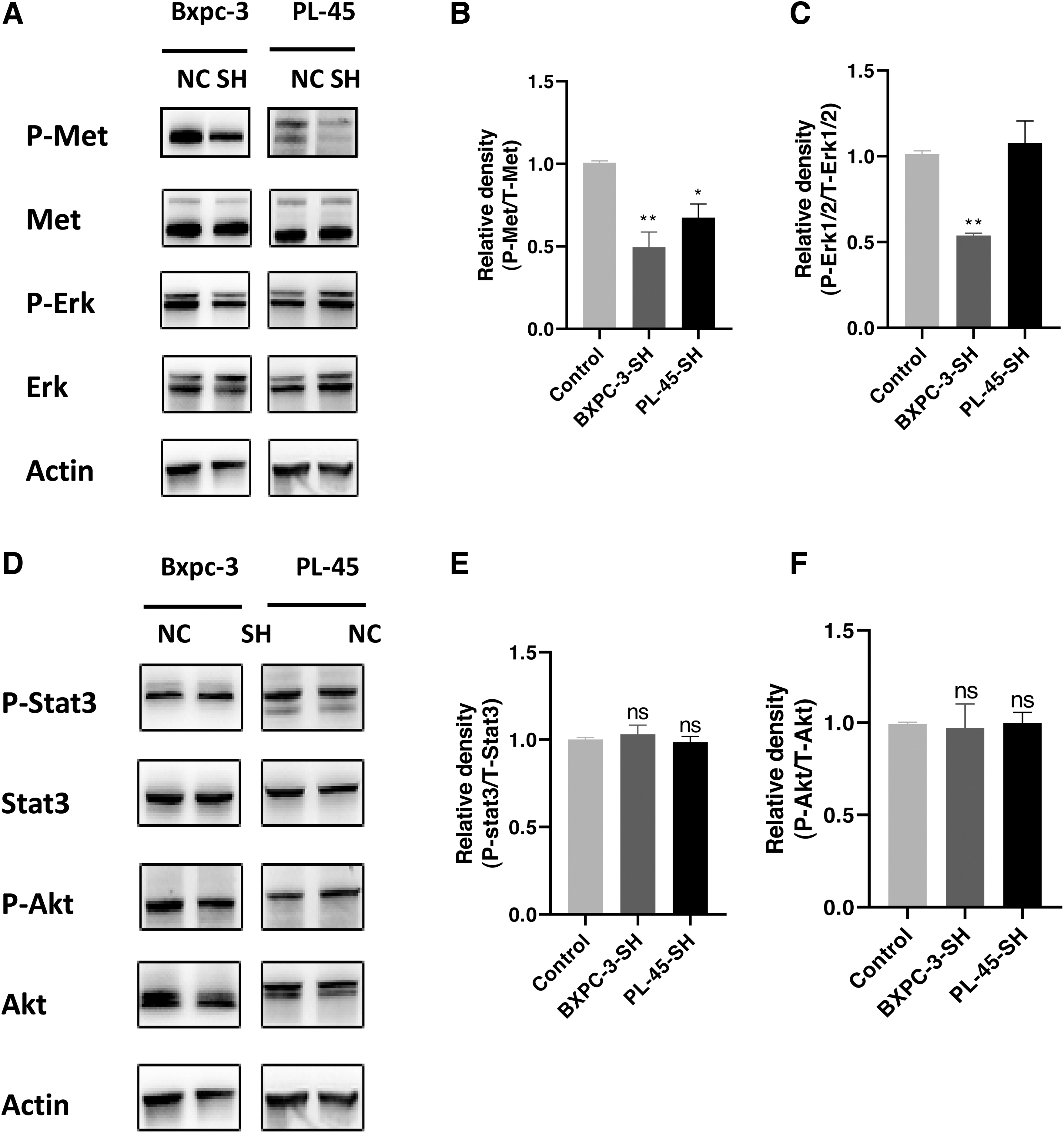
Western blot analysis of c-MET expression and downstream pathway components.
The authors found that silencing U35A not only inhibited the HGF-dependent phosphorylation of C-Met but decreased Erk phosphorylation in Bxpc-3 cells. However, phospho-Erk was elevated in PL-45 cells. The differences in ERK1/2 protein expression may be related to the degree of stemness in PCSCs (Fig. 6). In addition, Stat3 and Akt expression were not significantly changed in PCSCs (Fig. 5D–F). These findings show that U35A regulates the function of C-Met/HGF signaling by influencing C-Met phosphorylation. The mechanism of U35A affecting PCSCs is shown in Figure 7.

Flow cytometric determination of stem cell dryness in pancreatic cancer. PCSCs were 60% of BXPC-3 and 20% of PL-45 cells.
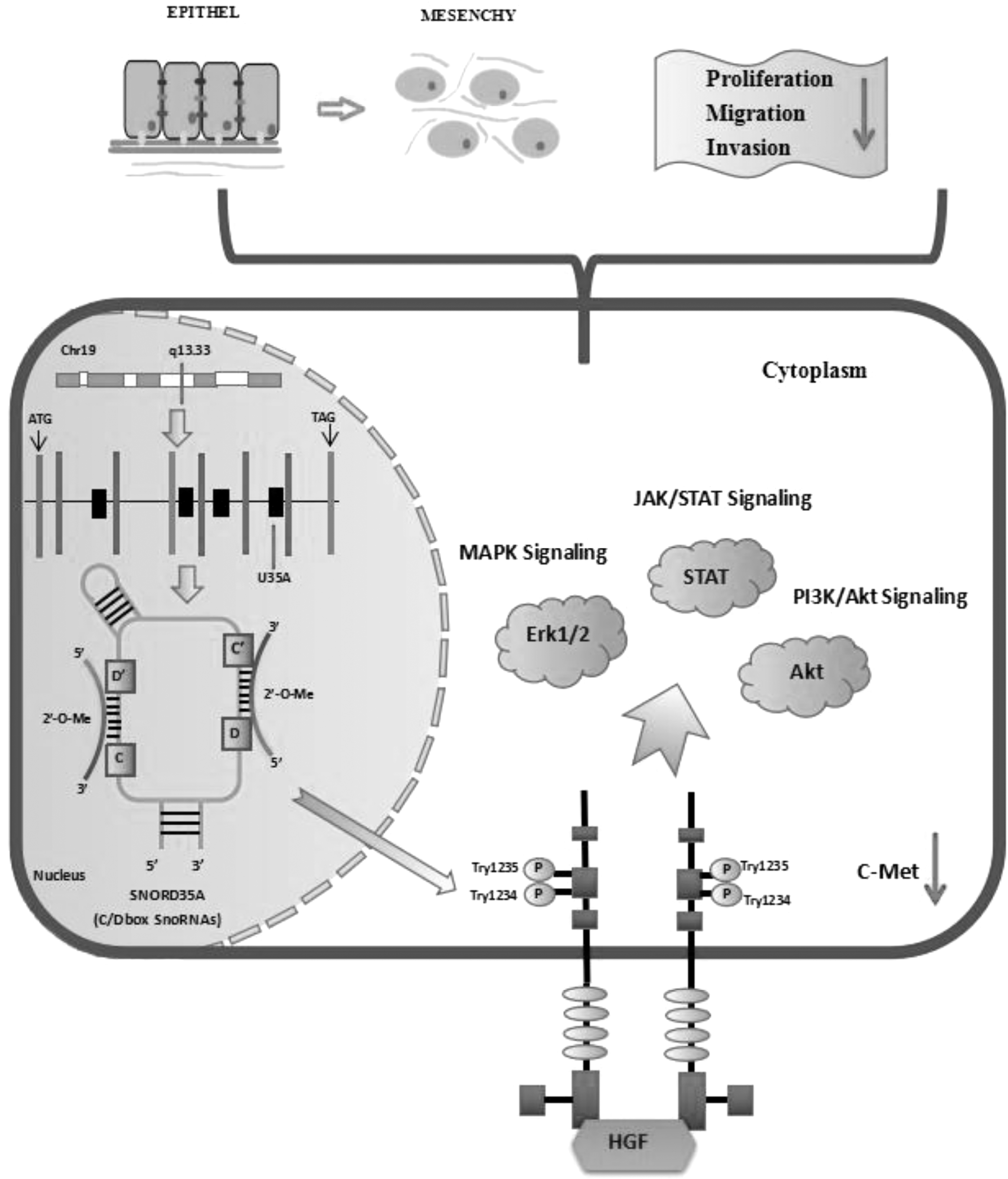
The mechanism through which SNORD35A influences PCSCs. In this study, downregulation of SNORD35A reduced C-Met phosphorylation. Downregulation of SNORD35A reduced the levels of interstitial markers and inhibited EMT and the migration and invasion of PCSCs by blocking C-MET signaling. In addition, knocking down SNORD35A inhibited xenograft tumor growth in mice.
Discussion
In this study, the authors explored the roles of U35A in PCSCs and demonstrated that it regulates the occurrence of PAAD. In contrast with normal pancreatic ductal epithelial cells, they confirmed that U35A is significantly upregulated in PAAD and that silencing U35A can alter the EMT status of PCSCs and decreased the occurrence of tumors. Meanwhile, animal experiments suggested that silencing U35A was associated with decreased tumor size. Western blotting indicated that U35A interfered with the growth of PCSCs through HGF/C-MET. In brief, silencing U35A inhibited the development of pancreatic stem cells.
One of the main findings of this study was that U35A is greatly overexpressed in pancreatic cancer tissues compared with normal paracancerous tissues. AffymetrixmiRNA 4.0 and quantitative PCR were used to screen out the significantly dysregulated U35A and SNORD14B in pancreatic cancer. SNORD14B is the only known intron that encodes multiple genes, but the relevant data are unclear. Therefore, the authors focused on the function of U35A, which is located on chromosome 19q13.33 and is encoded by the sixth intron of RPL13A. 17 The U35A gene is localized to nucleoli and plays a role in 2-O′-ribose methylation of rRNA. 18 Previously, it was generally believed that snoRNAs only had cellular housekeeping functions.
With more and more attention being paid to snoRNAs, recent reports have converged that their roles are diversified, and it has become a consensus that snoRNAs are dysregulated in a variety of cancers. 19 For example, in pancreatic ductal adenocarcinoma, RNAU91 and SNORA23 levels are significantly increased. 3,20 Moreover, in nude mice of the CDX model, it was found that high SNORA23 expression induced SYNE2 expression, thus promoting the growth and metabolism of tumor tissues. 3 Therefore, the upregulation of U35A in PAAD indicates that it may offer significant implications for the process of carcinogenesis.
Next, the authors transfected pancreatic stem cells in vitro, with the results showing that low U35A expression hindered the proliferation of PCSCs. Meanwhile, it had a significant effect on tumor migration and invasion. It also inhibited the transformation of cells to mesenchyme and reduced tumor volumes in the xenograft mice. Li and colleagues also suggested that changing the expression of snoRNAs can affect the growth and development of tumor cells. 21 The cure rate of cancer will be considerably improved when cells can be prevented from entering a metastatic state from a nonmetastatic state. According to previous studies, HGF/C-Met signaling is involved in the proliferation and invasion of cancer stem cells and is abnormally expressed in many types of solid tumors, including lung cancer, stomach cancer, breast cancer, skin cancer, and colorectal cancer. 22 –24
HGF is a mesenchymal-derived factor that regulates epithelial growth and morphogenesis. HGF induces the expression of EMT markers such as MMPs and is involved in the proliferation and invasion of cancer cells. In this study, U35A knockdown triggered decreased expression of N-cadherin (CDH2) and Vimentin (VIM). In addition, TMEFF1, SNAI2, and FN1 expression levels were greatly reduced in BXPC-3 cells; the authors also found downregulation of MMP2, SNAI2, FN1, and TMEFF1, and upregulation of OCLN in PL-45 cells. Thus, EMT by PCSCs is strongly associated with U35A. However, whether U35A affects EMT in PCSCs via C-Met signaling is unclear.
To focus on the mechanism through which U35A promotes tumorigenesis in PAAD, the authors detected the expression of the key signaling molecule C-Met. The experiments showed that silencing U35A inhibited C-Met and phospho-Met expression in PCSCs. Phosphorylation of Tyr1234/1235 in the Met kinase domain is closely related to kinase activation. First, the binding of Met to HGF induces autophosphorylation of multiple tyrosines, which then recruit multiple downstream signaling components, such as Gab1, c-CbI, PI3 kinase, and the transcriptional activator STAT. 25
Finally, it involves various signaling pathways such as RAS/MAPK, PI3K/Akt, and STAT, which have different effects on the formation and development of malignant tumors. The reduced expression of C-Met inhibited the growth and invasion in pancreatic CSCs, which supports the theory that U35A inhibited the occurrence, development, and metastasis of tumors by blocking the HGF/C-MET pathway. In addition, the C-Met pathway activates Erk1/2. This study suggested that downregulation of U35A significantly reduced the relative phosphorylation levels of Erk1/2 in Bxpc-3 cells.
However, there was no significant difference in PL-45 cells. It has been reported that ectopic expression of SNORA72 and SNORD89 can regulate the stemness of ovarian cancer stem cells and affect the levels of proteins related to the Notch1 pathway. 10,11 Therefore, the authors speculated that the degree of stemness in BXPC-3 and PL-45 affected the expression of ERK1/2 protein. The authors also detected the key signaling molecules Stat3, Akt, and did not observe differential expression of these proteins. There was no correlation between U35A and PI3K/Akt or STAT signaling in PCSCs. Thus, U35A has a crucial effect on tumor progression via HGF/C-Met signaling.
In conclusion, the authors explored the connection between U35A and pancreatic carcinoma stem cells. The results indicated that decreased U35A expression can inhibit HGF/C-Met signaling, which reduces tumor growth and development. This study provides ideas for gene therapy in PCSCs and supports the view that targeting C-Met could be a means of eradicating PCSCs in human tumors. Currently, the FDA has approved three targeted drugs for cancer stem cells, including Vismodegib, Ivosidenib, and Venetoclax. 26 The latest results from the Stanford University research team show that stem cell cancer vaccines can effectively prevent pancreatic cancer. 27 In general, the clinical transformation of tumor stem cells has a bright prospect, but it is still in its infancy, and further research is needed to fully understand the clinical potential of tumor stem cells.
Footnotes
Acknowledgments
The authors thank Fuxue Chen, Chenwei Li, and Huibin Zhou for their help. Finally, they thank James P. Mahaffey, PhD, from Liwen Bianji (Edanz) for editing the English text of a draft of this article.
Authors' Contributions
H.Z. and C.L. conducted the investigations. F.C., Y.Z., H.Z., and C.L. conceived and designed the study. Y.Z. and H.Z. collected and analyzed the data and performed research. Y.Z. wrote the article. F.C. and C.L. supervised the study. All authors approved the final version for submission.
Data Availability Statement
Data supporting the results of this article are available from the corresponding authors upon reasonable request.
Disclosure Statement
No potential conflicts of interest were disclosed by the authors.
Funding Information
This study was supported by the National Natural Science Foundation of China (52002239).
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.