
Abstract
Purpose:
The study aims to explore the roles and underlying mechanisms of long noncoding RNAs endogenous bornavirus-like nucleoprotein (lncRNA EBLN3P) in colon cancer, emphasizing the potential impact of these insights on advancing colon cancer treatment strategies. By shedding light on lncRNA EBLN3P’s involvement, this research could contribute to the development of novel therapeutic approaches, enhancing the efficacy of interventions for colon cancer patients.
Methods:
We employed quantitative reverse transcription polymerase chain reaction to assess the levels of lncRNA EBLN3P, zinc finger protein (ZFP91), and miR-519d-3p, alongside CCK-8 and EdU assays for cell proliferation, flow cytometry for apoptosis, and Transwell and wound healing assays for migration and invasion. The in vivo function of lncRNA EBLN3P was investigated through a xenograft model, and protein levels were evaluated via Western blot analysis.
Results:
LncRNA EBLN3P was found to be upregulated in colon cancer tissues and cells, promoting cell proliferation and metastasis while inhibiting apoptosis. Downregulation of lncRNA EBLN3P reduced tumor size, volume, and weight in a mouse model. MiR-519d-3p, which negatively interacts with lncRNA EBLN3P, was found to be downregulated in colon cancer tissues and cell lines. Its upregulation hindered cancer cell proliferation and metastasis while enhancing apoptosis. ZFP91, a binding partner of miR-519d-3p, was upregulated in colon cancer and inversely related to miR-519d-3p levels. Rescue experiments indicated that the effects of lncRNA EBLN3P silencing could be reversed by miR-519d-3p suppression, but were mitigated by ZFP91 downregulation.
Conclusion:
LncRNA EBLN3P facilitates colon cancer progression via the miR-519d-3p/ZFP91 axis, presenting a novel understanding of lncRNA EBLN3P’s role and offering potential therapeutic insights for colon cancer treatment. This study fills a critical gap by linking lncRNA EBLN3P with the miR-519d-3p/ZFP91 axis in the context of colon cancer, thereby broadening our understanding of the molecular mechanisms underlying colon cancer progression.
Introduction
Colon cancer is the third most prevalent malignant tumor in the digestive tract, with a mortality rate of 9% USA. 1 It has been estimated that there were approximately 1,148,515 confirmed cases of and 576,858 deaths due to colon cancer in 2020 worldwide. 2 The incidence of colon cancer varies greatly in different regions of the world, and the incidence of colon cancer has gradually increased in the world in recent decades. In China, the rising trend in the incidence of colon cancer cannot be ignored.3,4 Colon cancer can not only develop along the intestinal wall but also spread above and below the primary position of the longitudinal and radial intestinal canal and even infiltrate deep into the intestinal wall. In addition to blood, lymph, and local metastases, colon cancer can also directly implant into the abdominal cavity, spread along the incision surface, and metastasize along the suture line. 5 Currently, the main treatments for colon cancer are surgical resection, chemotherapy, radiation therapy, and immunotherapy. 6 However, since invasive colon cancer is prone to metastasis in the early stage and recurrence after treatment, it seriously affects the diagnosis, treatment, and prognosis of colon cancer patients.7,8 The occurrence and development of colon cancer are the result of the interaction of many pathogenic factors. A high-fat diet, food fiber deficiency, and intestinal flora disorder are the main environmental factors involved in colon cancer development. 9 Familial adenomatous polyposis and familial hereditary nonpolyposis are typical genetic factors leading to colon cancer, and gene mutation is also an important pathogenic factor that has been increasingly studied in recent years. 10 With the development of molecular biology and research, an increasing number of studies have shown that abnormal gene expression and regulation play an important role in the development of colon cancer. 11 KRAS and BRAF gene detection is of great significance to guide targeted therapy. Patients with intestinal tumors with KRAS gene mutation have poor efficacy to cetuximab or panitumumab. 12 BRAF gene mutation is closely related to the effect of tumor-targeted therapy. In another study, the authors showed that BARF gene mutation is the most important prognostic indicator of colorectal cancer, patients with BARF gene mutation have poor overall survival. 13 The BRAF gene wild type and cetuximab or panitumumab on the efficacy of colorectal cancer was positively correlated. 14 Therefore, the detection of KRAS and BRAF genes can be used to predict the efficacy of targeted drug-EGFR monoclonal antibodies. Thus, exploring new stable and reliable novel biomarkers can aid the development of effective prevention and treatment plans for colon cancer and ultimately prolong overall survival.
Long noncoding RNAs (lncRNAs) are a class of restriction RNAs composed of 200–600 nucleotides that generally do not encode proteins, but regulate gene expression levels in the form of RNA through the transcriptional regulation of epigenetic regulation and posttranscriptional regulation. 15 LncRNAs have recently received significant attention due to their regulation of epigenetics at the transcriptional and posttranscriptional levels in many physiological processes, affecting tumor initiation, development, invasion, and metastasis and their relationship with prognosis. 16 LncRNAs have been extensively proposed to be implicated in colon cancer development. For instance, LINC00657 was enriched in CRC stem-like cells (CSCs) and significantly promoted CSCs’ invasion ability. 17 At the posttranscriptional level, lncRNAs control cell fate as precursors of microRNAs (miRNAs/miRs) and competing endogenous RNAs (ceRNAs). Among those regulatory RNAs, ceRNAs, which regulate gene expression via miRNA mediation, have been extensively studied and recognized. 18 An increasing number of researchers have proposed the hypothesis of competitive endogenous RNA (ceRNA) in previous studies, which revealed that endogenous RNAs could mediate mRNA levels by sponging miRNAs. 19
LncRNA endogenous bornavirus-like nucleoprotein (LncRNA EBLN3P) has been identified to be involved in the progression of several cancers, such as osteosarcoma, 20 liver cancer, 21 and colorectal cancer. 22 It has been reported that the interaction of Ras-related protein 10 (Rab10) with miR-224-5p may underlie the oncogenic roles of the lncRNA EBLN3P in osteosarcoma progression. 20 Similarly, another study illustrated that EBLN3P may act as a ceRNA to modulate the dedicator of cytokinesis 4 (DOCK4) expression via downregulation of sponging miR-144-3p, leading to the regulation of hepatocellular carcinoma cell proliferation, migration, and invasion in vitro. 21 A recent study investigated whether EBLN3P is an oncogenic lncRNA facilitating CRC tumor formation by acting as a ceRNA to regulate the U2AF homology motif kinase 1 (UHMK1) expression state by sponging miR-323a-3p. 22 However, the molecular mechanism of lncRNA EBLN3P and miR-519d-3p in colon cancer remains largely elusive.
Building on these findings, our study introduces a novel aspect by exploring the role of lncRNA EBLN3P within the ceRNA framework. Unlike previously characterized ceRNAs, lncRNA EBLN3P exhibits unique regulatory features in the context of colon cancer. Preliminary data suggest that EBLN3P may serve as a molecular scaffold, orchestrating a complex ceRNA network that fine-tunes the regulatory effects of miR-519d-3p on key signaling pathways involved in colon cancer pathogenesis. This investigation not only sheds light on the multifaceted interactions within the ceRNA landscape but also highlights the potential of lncRNA EBLN3P as a therapeutic target, offering a fresh perspective on the intricate regulatory mechanisms that govern colon cancer progression.
Zinc finger protein (ZFP91) plays an important role in inflammation, tumors (gastric cancer, prostate cancer), and leukemia.23,24 A previous study indicated that the expression of ZFP91 was upregulated in colon cancer patients. ZFP91 upregulated hypoxia-inducible factor-1α (HIF-1α) protein levels in colon cancer cells. ZFP91 enhances colon cancer cell proliferation by upregulating HIF-lα in vitro and in vivo. Herein, whether miR-519d-3p participates in mediating the functions of lncRNA EBLN3P in colon cancer cells aroused our interest. In this study, we aimed to uncover the role of lncRNA EBLN3P in colon cancer, and its potential regulatory mechanisms. The data unveiled that lncRNA EBLN3P accelerated colon cancer by regulating the miR-519d-3p/ZFP91 axis, which may provide clues for the targeting therapy of colon cancer.
Materials and Methods
Patients and tissue samples
Thirty colon cancer tissues and corresponding adjacent tissues (5 cm from the edge of the tumor, confirmed by rapid pathological examination as normal tissue and tumor cells in the cutting edge remained negative tissue) were collected from Bayinguoleng Mongolian Autonomous Prefecture People’s Hospital from October 2019 to October 2020. All patients were diagnosed with colon cancer by pathological examination first and received neither chemotherapy nor radiotherapy as well as immunotherapy before surgery. The experiment was approved by the local Ethics Committee of Bayinguoleng Mongolian Autonomous Prefecture People’s Hospital in accordance with the Declaration of Helsinki guidelines [No. BZRMYY(2019)016]. The subjects were 30–65 years old, consisting of 19 males and 11 females. Other patient characteristics were as follows: TNM stage: 8 cases of stage II, 22 cases of stage III; pathological grade: 6 cases of grade II, 24 cases of grade III; and lymph node metastasis: positive in 27 cases, negative in 3 cases. Written informed consents were provided by all participants. All collected tissues were promptly frozen at −80°C for storage.
Cell culture and transfection
The American Type Culture Collection offered us the colon cancer cell lines (SW480, HT-29, LoVo, and HCT116) and the regular colonic epithelial cell line (NCM460). Cells were maintained in Roswell Park Memorial Institute—1640 medium with 5% CO2 at 37°C. Cells were grown on six-well plates before transfection, and short hairpin RNA targeting lncRNA EBLN3P or ZFP91 (sh-lncRNA EBLN3P or sh-ZFP91) and miR-30c-5p mimic/inhibitor were designed and synthesized by Gene Pharma (Shanghai, China) and respective controls were transfected into SW480 and T-29 cells with Lipofectamine 2000 (Invitrogen Life Technologies, USA).
Quantitative reverse transcription polymerase chain reaction
TRIzol reagent (Thermo Fisher Scientific) was used to isolate RNAs. Reverse transcription reagents (Applied Biosystems, Foster City, CA) were applied for cDNA reverse transcription for lncRNA or mRNA, while a TaqMan miRNA Reverse Transcription Kit (Applied Biosystems, Foster City) was used for miRNA. Quantitative reverse transcription polymerase chain reaction (RT-qPCR) was performed with SYBR Green (Takara Bio, Japan), and GAPDH and U6 were used as lncRNA/mRNA and miRNA internal controls, respectively. The relative lncRNA/mRNA and miRNA levels were calculated using the 2−ΔΔCt method. The primers are displayed in Table 1.
The Primers Used in This Study for RT-PCR

RT-PCR, reverse transcription polymerase chain reaction.
Flow cytometry analysis
Colon cancer cell apoptosis was evaluated using an Apoptosis Detection Kit (BD Biosciences, San Jose, CA). SW480 and HT-29 cells were incubated for 72 h after transfection for 24 h followed by washing. Annexin V-FITC (5 μL) was used to stain the cells for 15 min, followed by incubation with PI (5 μL) for 10 min in the dark. A FACSCanto II flow cytometer (BD Biosciences) was used to analyze the cells.
Transwell assays
Transwell chambers (Corning, USA) with/without Matrigel (Becton Dickinson, USA) was applied for SW480 and HT-29 cells invasion/migration assessment. After transfection, cells and serum-free medium were placed in the upper chamber, while in the lower chamber, a medium with bovine calf serum (10%) was added. Forty-eight hours later, methanol was used to fix the migrated or invaded cells, and crystal violet (0.1%) was applied for cell staining. Finally, the migrated or invaded cells were counted via a microscope.
Xenograft model
Animal studies were conducted in accordance with the National Institutes of Health (NIH) Guide for the Care and Use of Laboratory Animals (National Academies Press 2011) and approved by the ethical committees of Bayinguoleng Mongolian Autonomous Prefecture People’s Hospital [No.BZRMYY(2021)003]. Twelve BALB/c nude male mice were acquired from Wuhan Hualianke Biotechnology (Wuhan, China). SW480 cells transfected with sh-lncRNA EBLN3P or sh-NC were subcutaneously injected into the left flanks of the mice. Four weeks later, the mice were anesthetized (with 5 mL/kg of 1% pentobarbital, i.p.) and artificially ventilated using a respirator. Then, tumor tissues were collected for further analyses.
Luciferase reporter assay
Bioinformatics tools predicted the putative binding sequences for miR-519d-3p on lncRNA EBLN3P and ZFP91 3′-UTR. Wild-type (WT) and mutant (Mut) sequences of lncRNA EBLN3P (lncRNA EBLN3P-WT/Mut) or ZFP91 (ZFP91-WT/Mut) were respectively, put into pmirGLO dual-luciferase reporters (Promega, Madison, WI) to generate respective reporter vectors. Then, lncRNA EBLN3P-WT/Mut or ZFP91-WT/Mut were separately transfected with a miR-519d-3p mimic in SW480 and HT-29 cells. After 48 h, the luciferase activity was assessed.
Wound healing assay
Cells were placed onto six-well plates and grown to 100% confluence. Afterward, a confluent monolayer was scratched, followed by culturing in a medium for 48 h. An IX71 Olympus light microscope (magnification × 4; Olympus) was employed to observe the width of wounds at 0 and 48 h after scratching. ImageJ (National Institutes of Health) was applied to analyze the migration.
Western blot analysis
Colon cancer cells were lysed using RIPA lysis buffer, and a BCA protein assay kit (Thermo Fisher Scientific, MA) was applied to determine the concentration. Then, proteins were separated by polyacrylamide gel electrophoresis (PAGE) followed by transfer to a polyvinylidene fluoride (PVDF) membrane (Amersham, USA). After blocking with nonfat milk, primary antibodies were added to the membrane including PCNA (ab29, 1 μg/mL, Abcam, Shanghai, China), Ki67 (ab16667, 1:1,000, Abcam), Bax (ab182734, 1:1,000, Abcam), cleaved-caspase 3 (ab2302, 1:500, Abcam), cleaved-caspase 9 (ab2324, 0.5 μg/mL, Abcam), Bcl-2 (ab182858, 1:2,000, Abcam) MMP2 (ab86607, 1 μg/mL, Abcam), MMP9 (ab283575, 1:1,000, Abcam), ZFP91 (ab30970, 2.5 μg/mL, Abcam) and β-actin (ab8226, 1 μg/mL, Abcam), for one night at 4°C. Subsequently, the membranes were incubated with the secondary antibody (ab6721, 1: 2,000, Abcam) for 1 h at room temperature. After washing, an optical luminometer (GE, USA) was applied for protein band visualization.
EdU assay
EdU assays were performed using an EdU assay kit (Life Technologies, Carlsbad, CA) to measure cell proliferation. Cells were plated in 24-well plates and precoated using fresh laminin (Sigma–Aldrich, USA). Then, EdU (10 μM) was supplemented and incubated for 2 h. After rinsing, a Click-iT EdU kit was used for EdU measurement. Furthermore, Hoechst 33342 was added to stain the cells, and a fluorescence microscope (Olympus, Tokyo, Japan) was used for visualization. Finally, Image-Pro Plus 6.0 software (Media Cybernetics, Rockville, MD) was employed to count the EdU-positive cells.
TUNEL assay
Tumor tissues were cultured with neutral formaldehyde for 48 h, and the TUNEL assay was performed via the Cell Death Detection Kit (Roche, Mannheim, Germany). A fluorescence microscope (200 × magnification, Olympus, Japan) was used to observe the images.
CCK-8 assay
A CCK-8 assay was performed to assess colon cancer cell viability. Cells were grown on a 96-well plate and cultured for 2 h followed by the supplementation with CCK-8 solution at 0 h, 24 h, 48 h, and 72 h. After culturing for 2 h, the absorbance (450 nm) was assessed.
Hematoxylin-eosin staining
Tumor tissues were dehydrated, embedded, and cut into sections, followed by dewaxing in xylene and dehydrating in alcohol. Then, the sections were subjected to hematoxylin and differentiation via hydrochloric acid alcohol. After immersion in ammonia water for 20 s, the sections were subjected to eosin staining and dehydrated. Then, the pathological changes in the tissues were examined via a microscope.
Statistical analysis
Data are displayed as the mean ± standard deviation (SD). SPSS version 18.0 software (SPSS, Chicago, IL) was used for data analyses, and all experiments were performed more than twice. Comparisons between or among groups were made with Student’s t-test or one-way ANOVA. p < 0.05 indicated statistical significance.
Results
LncRNA EBLN3P expression was enhanced in colon cancer
The expression of lncRNA EBLN3P in tissues was evaluated to assess the function of lncRNA EBLN3P in colon cancer. The data revealed that the lncRNA EBLN3P level was elevated in colon cancer tissues relative to normal tissues (Fig. 1A). In addition, lncRNA EBLN3P was also enhanced in SW480, HT-29, LoVo, and HCT116 cells compared with NCM460 cells (Fig. 1B). In conclusion, lncRNA EBLN3P expression was enhanced in colon cancer.
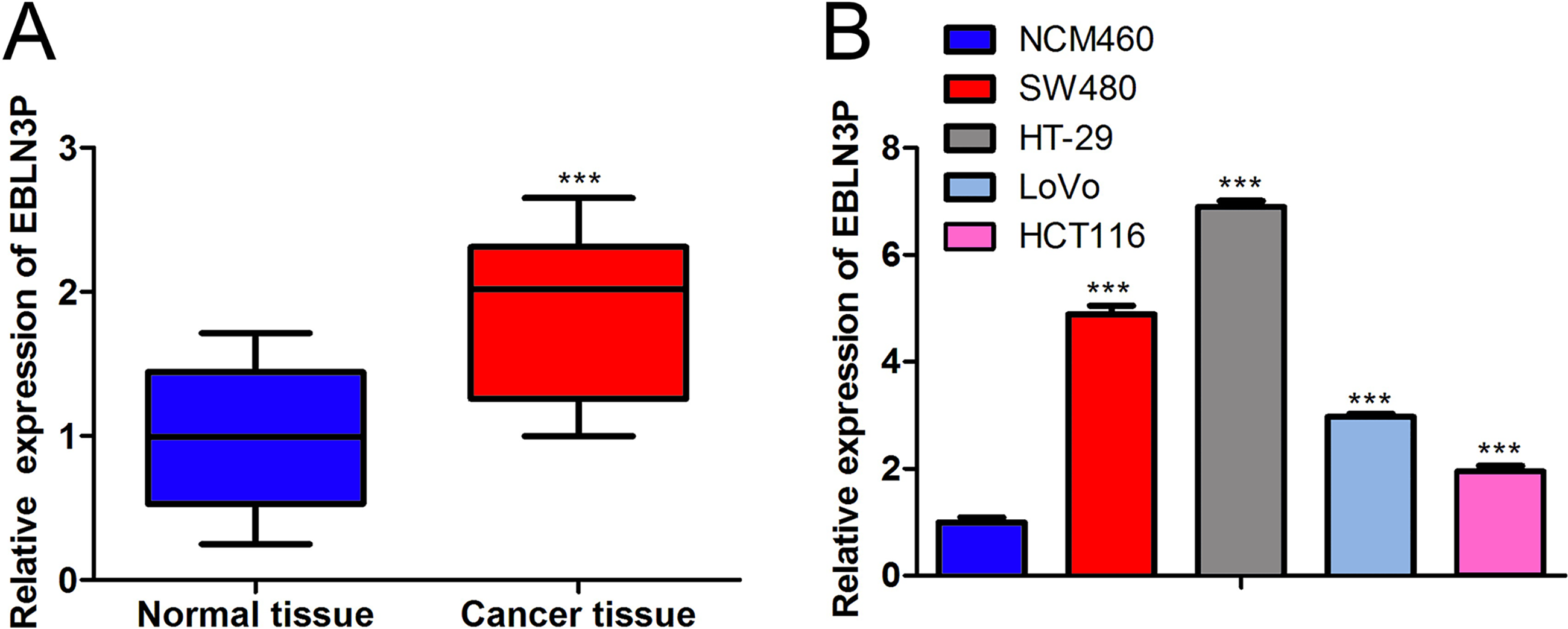
LncRNA EBLN3P expression was boosted in colon cancer.
Silencing of lncRNA EBLN3P retarded cell proliferation and boosted cell apoptosis in colon cancer
Furthermore, the function of lncRNA EBLN3P in colon cancer was assessed. Overall, the transfection efficiency of sh-lncRNA EBLN3P in colon cancer cells was evaluated, revealing that the lncRNA EBLN3P level was evidently decreased due to sh-lncRNA EBLN3P transfection (Fig. 2A). The CCK-8 assay revealed that SW480 and HT-29 cell viability was distinctly reduced as a result of lncRNA EBLN3P downregulation (Fig. 2B). Moreover, cell proliferation was attenuated by lncRNA EBLN3P depletion (Fig. 2C). In addition, the protein levels of proliferation markers (PCNA and Ki67) were strikingly suppressed as a result of lncRNA EBLN3P silencing (Fig. 2D). In addition, cell apoptosis was accelerated by a decrease in lncRNA EBLN3P (Fig. 2E). Similarly, the levels of the cell apoptosis markers Bax, cleaved-caspase 3 and cleaved-caspase 9 were enhanced, while Bcl-2 was declined because of the reduction in lncRNA EBLN3P (Fig. 2F). In summary, silencing lncRNA EBLN3P retarded cell proliferation and boosted cell apoptosis in colon cancer.
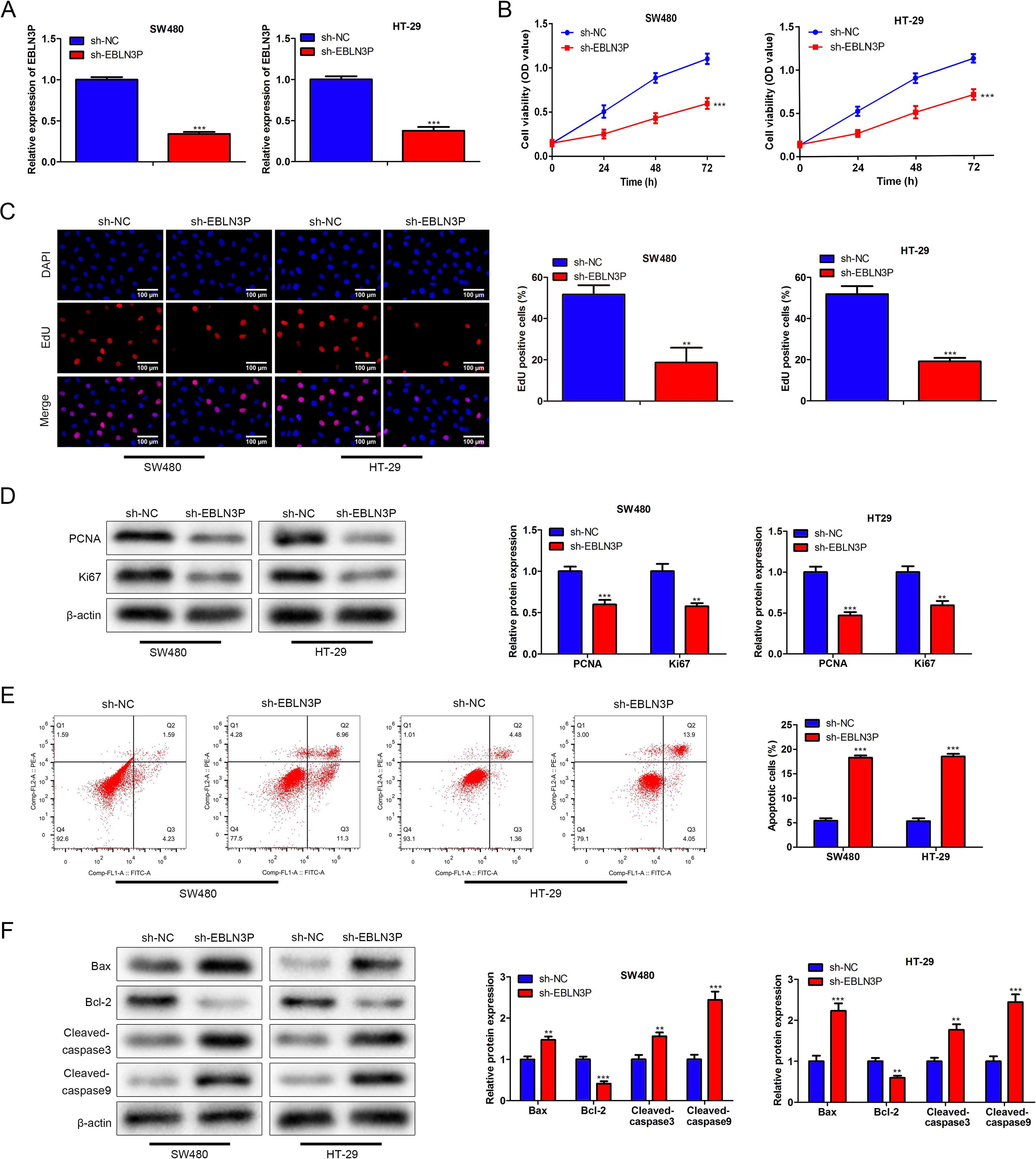
Silencing of lncRNA EBLN3P attenuated cell proliferation and enhanced cell apoptosis in colon cancer.
Restraint of lncRNA EBLN3P repressed cell metastasis in colon cancer
Afterward, the metastatic ability of colon cancer cells by lncRNA EBLN3P inhibition was measured. Wound healing assays manifested that the wound width was decreased after 48 h in the sh-NC group relative to the sh-lncRNA EBLN3P group (Fig. 3A), indicating that lncRNA EBLN3P attenuation reduced colon cancer cell migration. Consistently, cell invasion was also inhibited due to lncRNA EBLN3P downregulation (Fig. 3B). In addition, cell migration- and invasion-associated proteins (MMP2 and MMP9) were attenuated by lncRNA EBLN3P suppression (Fig. 3C). Overall, restraint of lncRNA EBLN3P repressed cell metastasis in colon cancer.
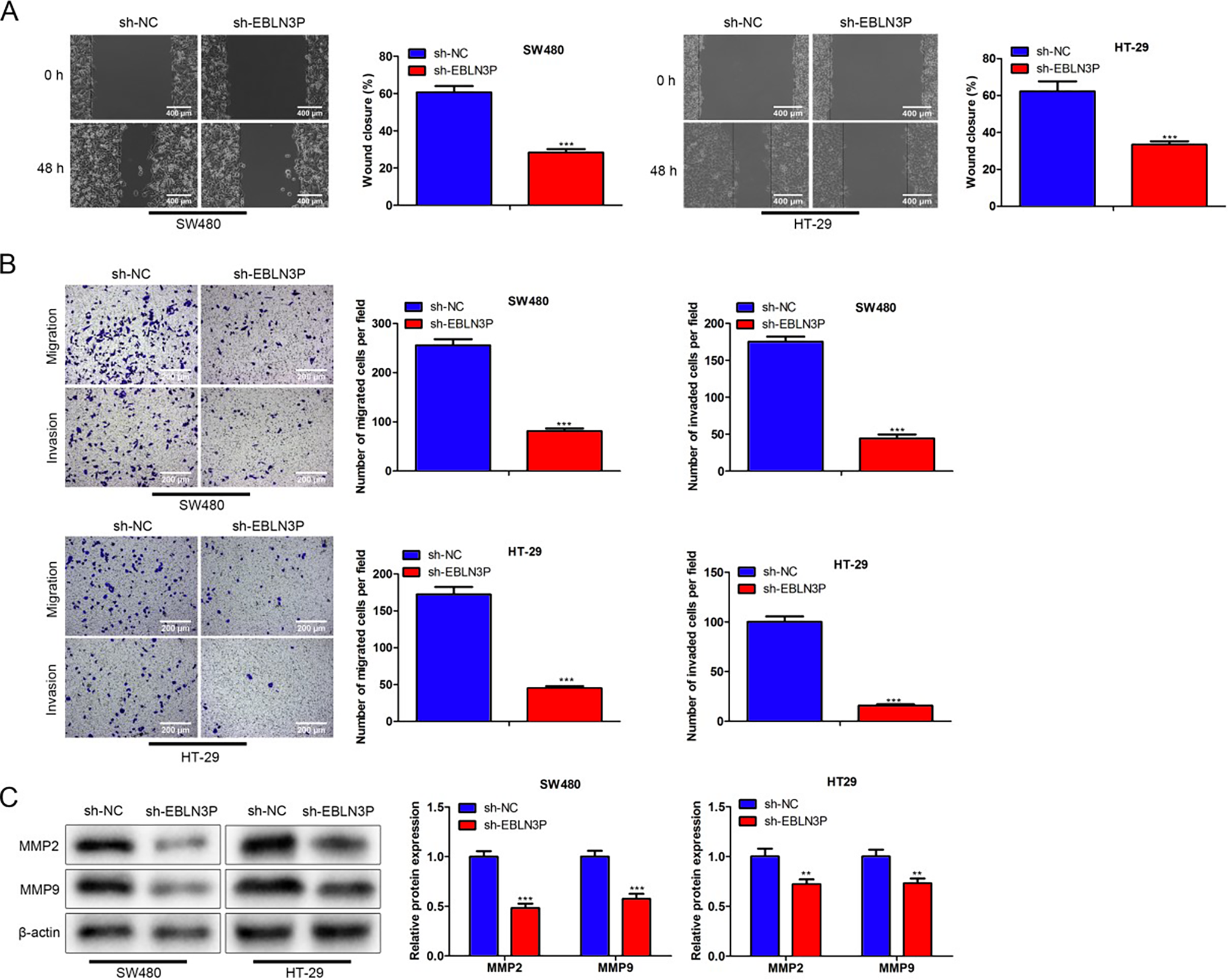
Restraint of lncRNA EBLN3P repressed cell metastasis in colon cancer.
LncRNA EBLN3P sponged miR-519d-3p
Next, the mechanism of lncRNA EBLN3P in colon cancer was investigated. Based on ENCORI, 10 miRNAs with potential binding sites to lncRNA EBLN3P were selected under the screening condition of CLIP-Data ≥ 3 and Pan-Cancer = 1 (Supplementary Fig. S1B). The results of RT-qPCR analysis revealed that only the expression of miR-519d-3p showed a significant increase after lncRNA EBLN3P silencing (Supplementary Fig. S1B). Moreover, according to a previous study, miR-519d-3p was involved in colorectal cancer tumorigenesis. 25 Thus, miR-519d-3p was chosen for further study (Fig. 4A). It was determined that miR-519d-3p enhancement decreased the luciferase reporter activity of lncRNA EBLN3P-WT, but the luciferase reporter activity of lncRNA EBLN3P-Mut disclosed no change in miR-519d-3p mimic and negative control (NC) mimic groups (Fig. 4B), suggesting that lncRNA EBLN3P sponged miR-519d-3p. Moreover, lncRNA EBLN3P attenuation enhanced the miR-519d-3p level (Fig. 4C). In addition, miR-519d-3p expression was lower in colon cancer tissues and cell lines (Fig. 4D–E). Taken together, the results demonstrated that lncRNA EBLN3P sponged miR-519d-3p.
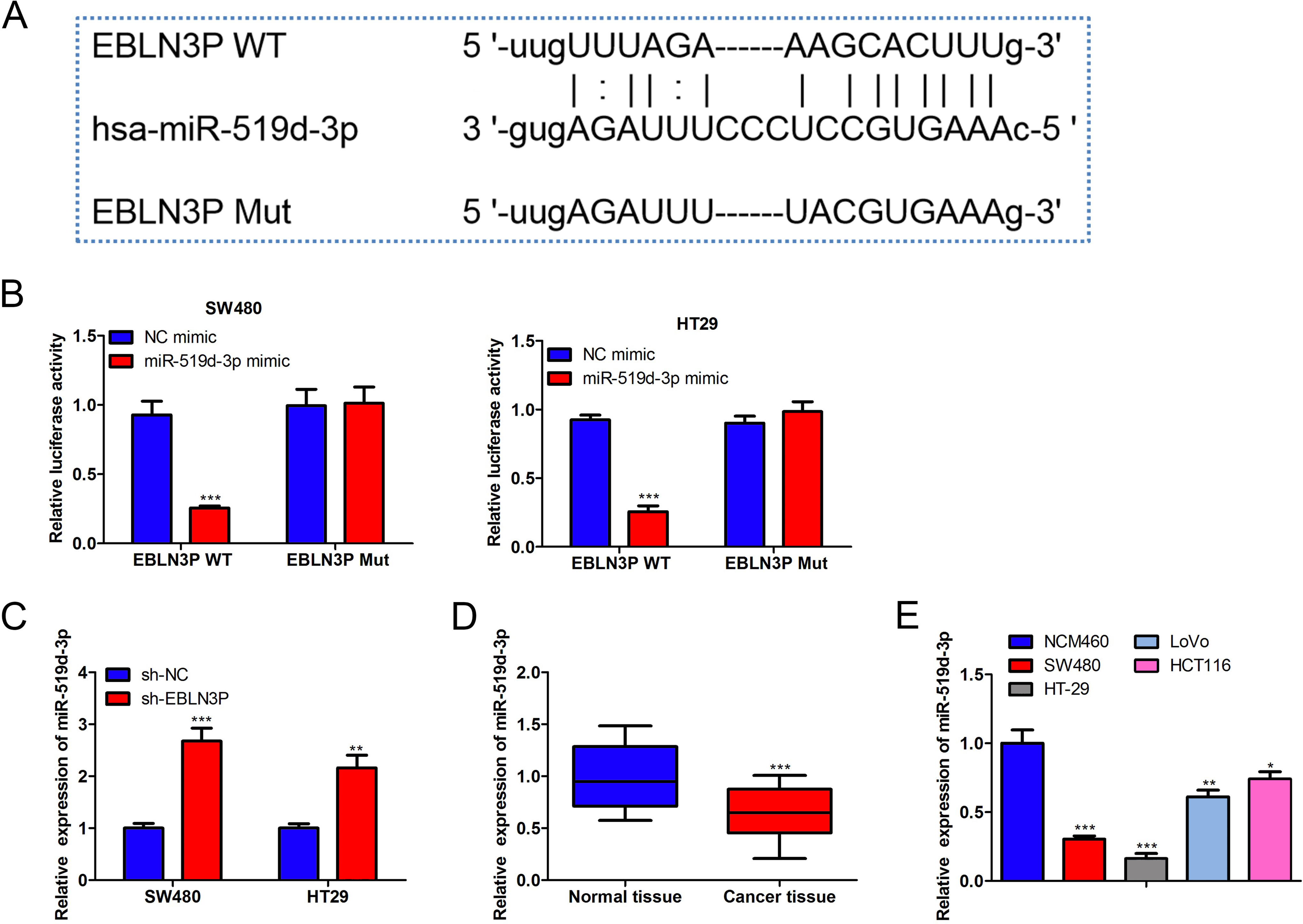
LncRNA EBLN3P sponged miR-519d-3p.
MiR-519d-3p enhancement repressed cell growth and metastasis in colon cancer
Subsequently, the function of miR-519d-3p was investigated. As shown in Figure 5A, miR-519d-3p expression was increased due to miR-519d-3p mimic transfection. MiR-519d-3p mimics markedly restrained cell viability and proliferation in colon cancer (Fig. 5B–C). Additionally, cell apoptosis was elevated due to miR-519d-3p overexpression (Fig. 5D). Moreover, upregulation of miR-519d-3p also attenuated the migration and invasion capacities (Fig. 5E). Overall, miR-519d-3p enhancement repressed cell growth and metastasis in colon cancer.
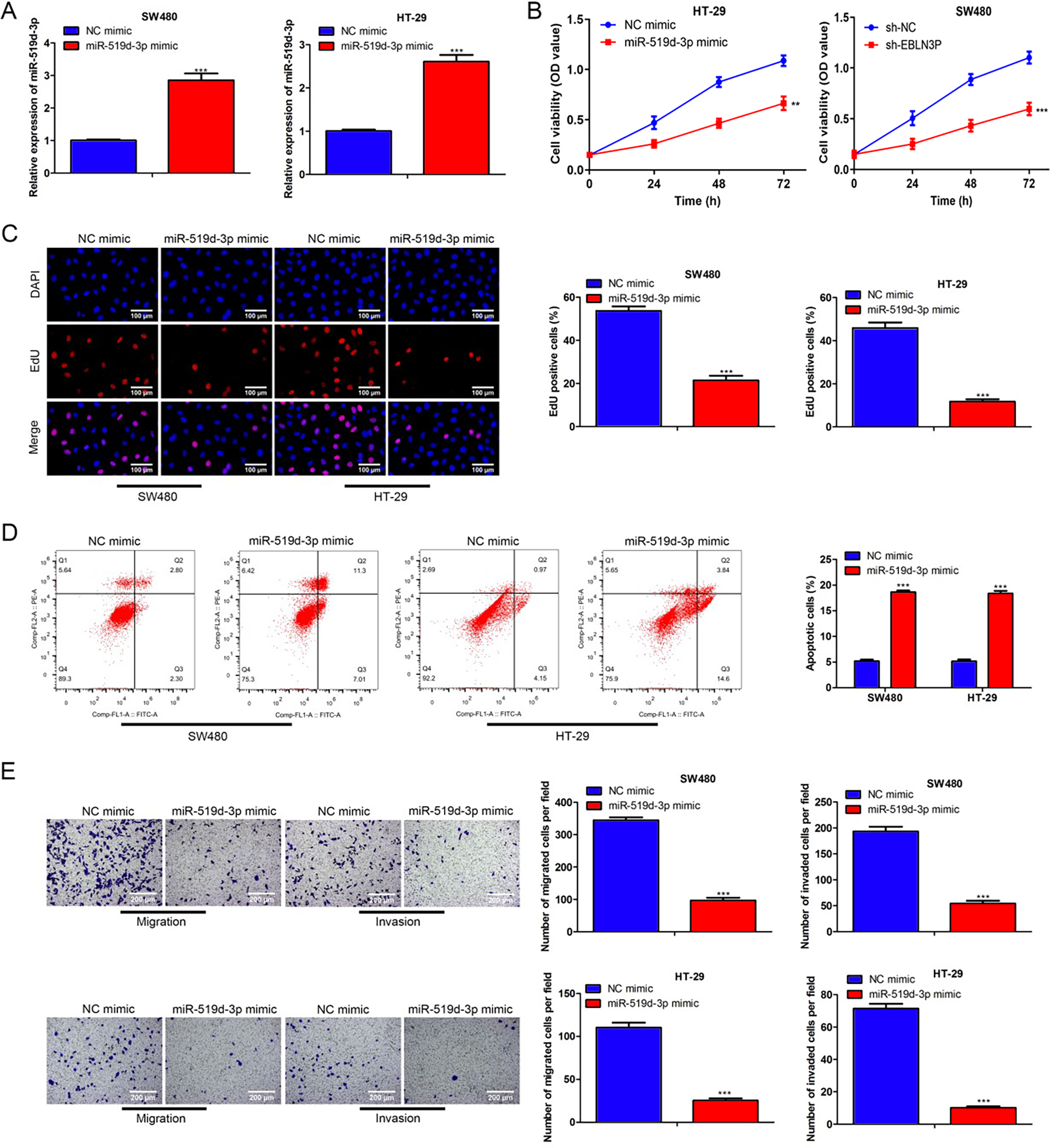
MiR-519d-3p enhancement suppressed cell growth and metastasis in colon cancer.
MiR-519d-3p targeted ZFP91
Then, the target of miR-519d-3p was identified. Bioinformatics analysis was performed using the ENCORI, and mRNAs potentially binding to miR-519d-3p were selected under the condition of CLIP-Data ≥ 5, Pan-Cancer = 5, programNum ≥ 4, program: microT, miRanda, miRmap, PITA, TargetScan, and clipExpNum >15 (Supplementary Fig. S2A). The Venn chart uncovered 22 potential targets of miR-519d-3p, and the results of RT-qPCR analysis revealed that only the expression of ZFP91 was significantly decreased in colon cancer cells after miR-519d-3p overexpression (Supplementary Fig. S2B). Furthermore, based on a previous study, ZFP91 is demonstrated to be involved in colon cancer development. 26 Hence, ZFP91 was chosen, and the potential binding sequences for miR-519d-3p and ZFP91 are displayed in Figure 6A. The luciferase activity of ZFP91-WT was decreased due to miR-519d-3p enhancement, but the luciferase activity of ZFP91-Mut remained the same in the miR-519d-3p mimic and NC mimic groups (Fig. 6B). Besides, the ZFP91 level was increased in colon cancer tissues and cell lines (Fig. 6C–D). The mRNA and protein expression levels of ZFP91 were reduced due to miR-519d-3p overexpression (Fig. 6E–F). In summary, miR-519d-3p targeted ZFP91.
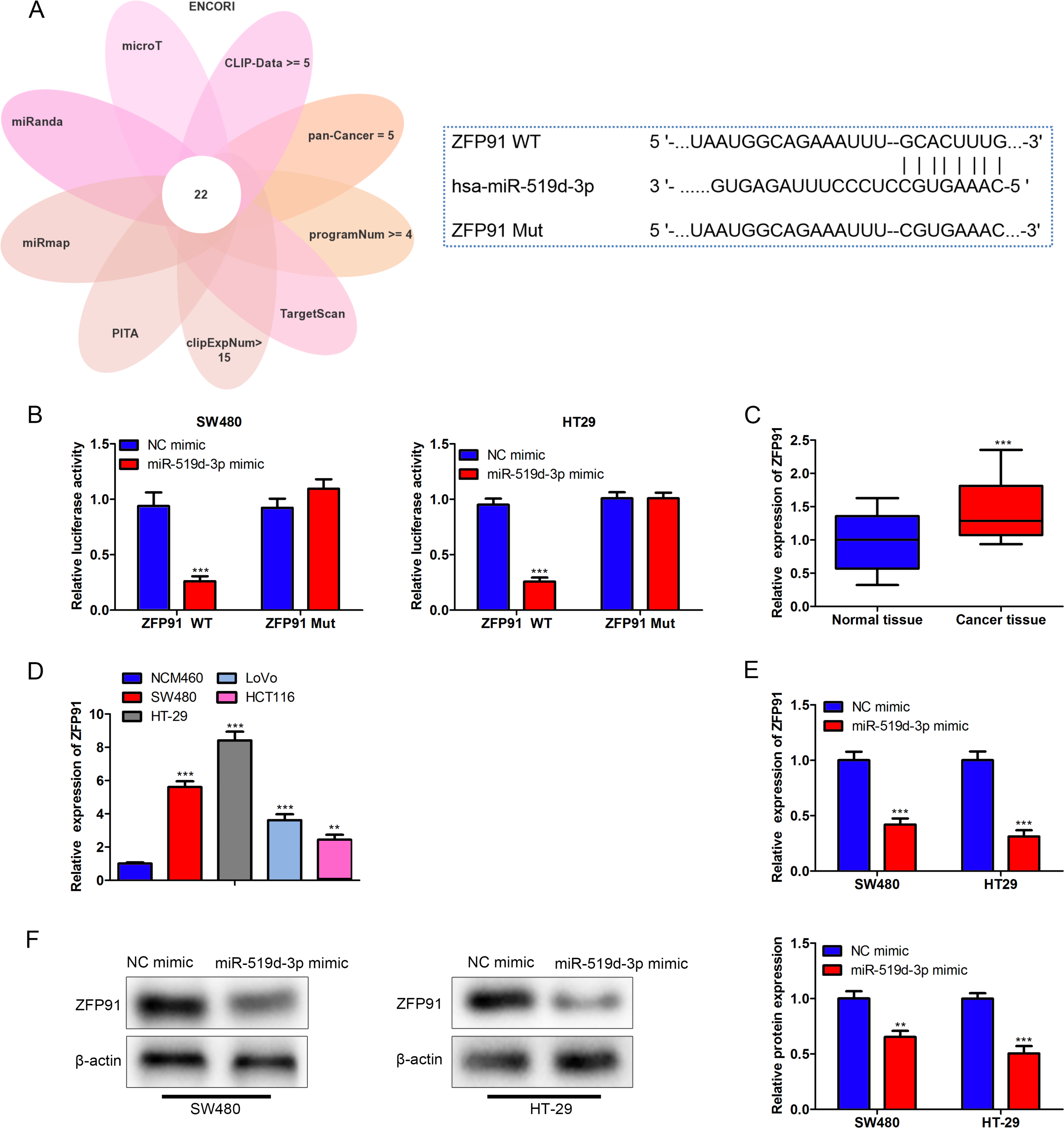
MiR-519d-3p targeted ZFP91.
LncRNA EBLN3P/miR-519d-3p/ZFP91 axis modulates the development of colon cancer
Then, rescue assays were performed to validate whether lncRNA EBLN3P mediated the development of colon cancer through the miR-519d-3p/ZFP91 axis. The data revealed that the miR-519d-3p inhibitor was effectively transfected into SW480 cells to decrease miR-519d-3p (Fig. 7A). Similarly, sh-ZFP91 efficiently downregulated the ZFP91 level in SW480 cells (Fig. 7B). Furthermore, miR-519d-3p suppression reversed the lncRNA EBLN3P decline attenuated cell viability and proliferation, while these effects were further offset by ZFP91 silencing (Fig. 7C–D). In contrast, silencing miR-519d-3p counteracted sh-lncRNA EBLN3P mediated promotion of cell apoptosis, but this effect was inversely changed due to ZFP91 depletion (Fig. 7E). In addition, reduced cell migration and invasion by sh-lncRNA EBLN3P was reversed because of the miR-519d-3p inhibitor, but these effects were neutralized as a result of ZFP91 attenuation (Fig. 7F). In a word, the lncRNA EBLN3P/miR-519d-3p/ZFP91 axis modulates the development of colon cancer.
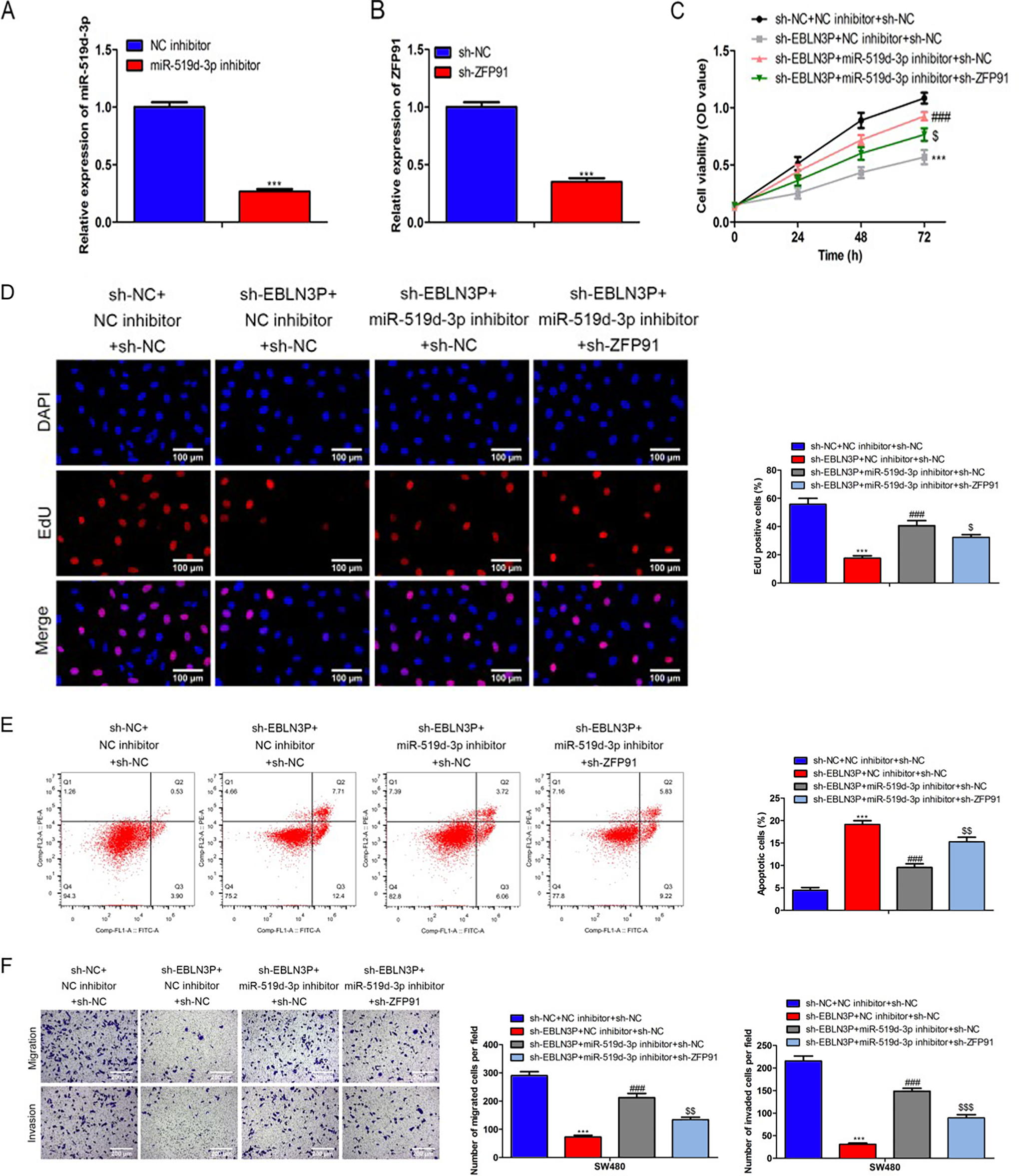
LncRNA EBLN3P/miR-519d-3p/ZFP91 axis modulates the development of colon cancer.
LncRNA EBLN3P promoted colon cancer growth in vivo
Finally, the mouse model of colon cancer was constructed to identify the role of lncRNA EBLN3P in vivo. As manifested in Figure 8A–B, lncRNA EBLN3P silencing decreased the tumor size, tumor volume, and tumor weight in vivo. Furthermore, hematoxylin-eosin (H&E) staining revealed that the tissue damage in the sh-lncRNA EBLN3P group was alleviated relative to that in the sh-NC group. The TUNEL assay also showed that cell apoptosis was aggravated by lncRNA EBLN3P attenuation, while Ki67 and PCNA expression was decreased due to lncRNA EBLN3P suppression (Fig. 8C). In addition, lncRNA EBLN3P and ZFP91 levels were reduced, whereas miR-519d-3p levels were enhanced in the sh-lncRNA EBLN3P group (Fig. 8D). In conclusion, lncRNA EBLN3P promoted colon cancer tumor growth in vivo.
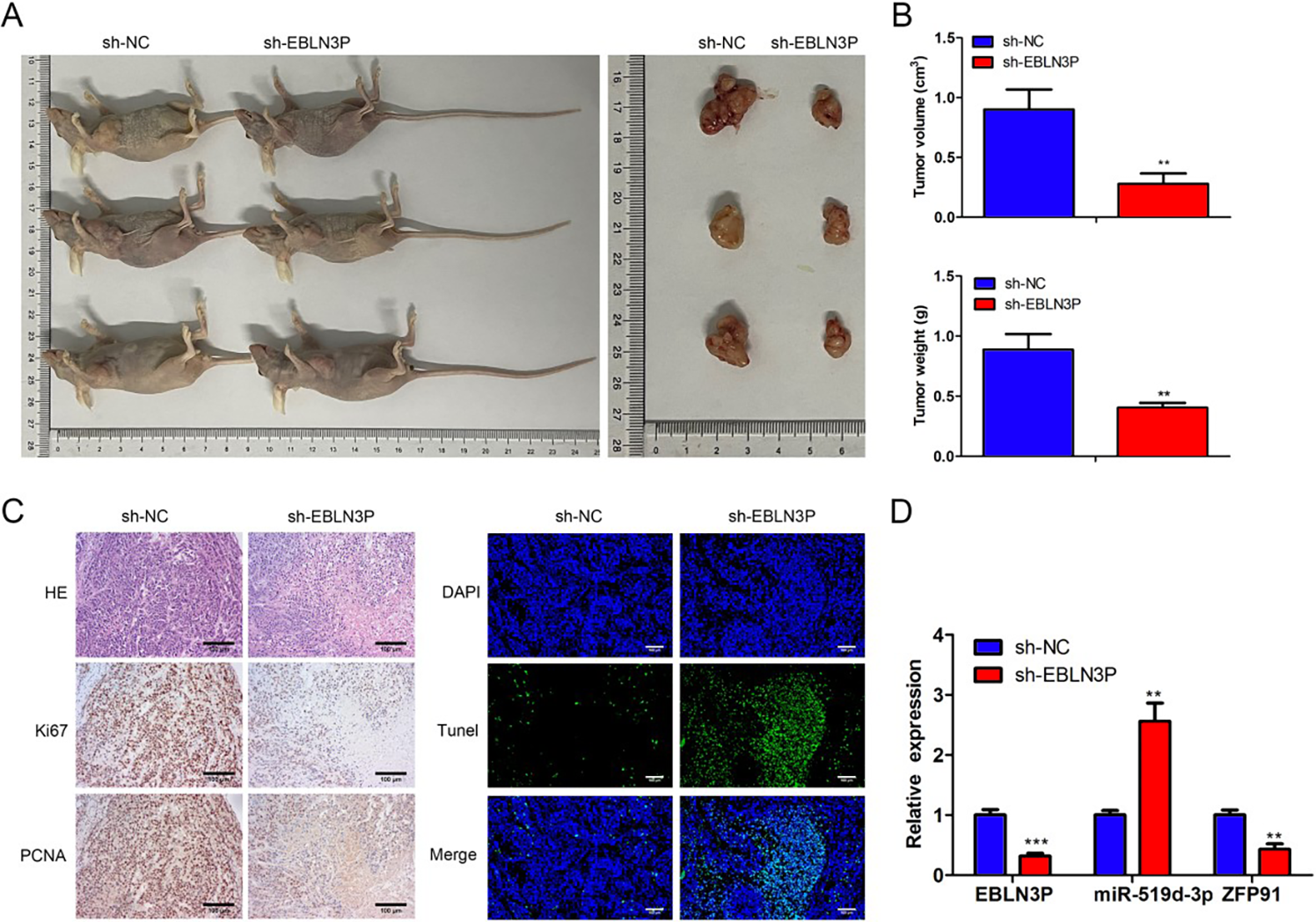
LncRNA EBLN3P depressed tumor growth of colon cancer in vivo.
Discussion
Colon cancer is one of the main causes of cancer-related death in China, with the highest incidence in the 40- to 75-year-old group. Based on a previous study, approximately 2,000,000 new cases and 1,000,000 deaths will occur annually by 2030. 27 With improvements in surgery and adjuvant therapy, the prognosis of colon cancer has improved, but the 5-year survival rate of advanced colon cancer patients is still less than 10%. 28 Finding and clarifying new genes related to colon cancer and their biological functions have important clinical significance for evaluating the prognosis of colon cancer patients and guiding clinical treatment. A comprehensive analysis of the mammalian genome shows that most genome products are transcribed into lncRNAs. A mushrooming number of evidence show that from the integration of chromatin remodeling complexes to the regulation of transcription and posttranscriptional processes, lncRNA expression disorders are related to the occurrence, progression, and poor prognosis of tumors, and may be used as a new biomarker of tumors.29,30
Although a variety of potential biomarkers have been explored, specific biomarkers for colon cancer have not yet been confirmed. Previous studies on the genomic characterization of functional lncRNA loci in human cells also revealed the existence of lncRNA EBLN3P. 31 Previously, Xu et al. first reported that lncRNA EBLN3P was overexpressed in CRC. They performed in vitro experiments, which revealed that forced EBLN3P expression contributed to the promotion of the proliferation, migration, and invasion of SW480 and HCT116 cells via alteration of the miR-323a-3p/U2AF homology motif kinase 1(UHMK1) axis. 22 Our findings were consistent with the oncogenic roles of lncRNA EBLN3P in colorectal carcinoma.
The present study illustrated that lncRNA EBLN3P is dramatically upregulated in colon cancer tissues and cell lines. In vitro, functional assays indicated that the deprivation of EBLN3P inhibited colon cancer cell proliferation, migration, and invasion, and facilitated cell apoptosis. Besides, lncRNA EBLN3P downregulation also attenuated the tumor size, tumor volume, and tumor weight in the xenograft model. The transfection efficiency of lncRNA EBLN3P and the difference in mouse age or weight may contribute to the difference in the tumor weight and volume with the previous studies. Additionally, lncRNA EBLN3P silencing inhibited the expression of Ki67 and PCNA in the xenograft model. In summary, lncRNA EBLN3P aggravated the development of colon cancer in vitro and in vivo, exemplifying the potential of lncRNA EBLN3P as a therapeutic target for colon cancer interventions.
MicroRNAs (miRNAs) are small molecule RNAs that bind to the 3ʹ-UTR of mRNAs to mediate their expression. 32 Recent studies have shown that miRNAs are small noncoding RNA molecules that regulate target protein expression and play a key role as tumor promoters or inhibitors. 33 MiR-519d-3p has been shown to suppress non-small cell lung cancer development by modulating VEGFA and inhibiting the PI3K/AKT pathway. 34 MiR-519d-3p attenuates oral squamous cell carcinoma (OSCC) progression by mediating cell growth and the cell cycle and targeting cyclin D1(CCND1). 35 The miR-519d-3p/B-Cell Lymphoma 6 axis alleviates gastric cancer progression. 36 Nevertheless, whether lncRNA EBLN3P sponges miR-519d-3p to regulate colon cancer is largely unknown.
Bioinformatics analyses revealed that there was a conserved binding site of miR-519d-3p on lncRNA EBLN3P. Therefore, it was inferred that lncRNA EBLN3P could affect miR-519d-3p via a ceRNA mechanism. It was validated that miR-519d-3p expression was negatively correlated with lncRNA EBLN3P expression. Moreover, miR-519d-3p expression was enhanced in both colon tissues and cell lines. Additionally, upregulation of miR-519d-3p also attenuated the proliferation, migration, and invasion, and increased apoptosis of colon cancer cells in vitro. Importantly, the dual-luciferase assay further confirmed that lncRNA EBLN3P directly interacted with miR-519d-3p to reduce its expression, indicating that lncRNA EBLN3P serves as the miRNA sponge that binds to and regulates miR-519d-3p expression in the development of colon cancer. Our findings were consistent with the oncogenic roles of lncRNA EBLN3P in CRC cancer reported by Xu. 22 This result suggests that miR-519d-3p may be a valuable marker for colon cancer.
The miRNAs control gene expression by binding to the 3’ untranslated region’ (-UTR) of the target gene, which causes mRNA cleavage or translational repression. 37 To elucidate the molecular mechanism of miR-519d-3p, we used the ENCORI to predict that ZFP91 may be one of the targets of miR-519d-3p. In addition, Peng et al. reported that ZFP91 is a target protein of miR-188-5p in gastric cancer. 38 Therefore, we chose the reported oncogene ZFP91 for further study. ZFP91 was positively correlated with HIF-lα in human colon cancer. ZFP91 promotes colon cancer cell proliferation and tumorigenesis by activating transcription factors NF-κB/p65 and HIF-lα. 26 In this study, we confirmed that the 3′UTR of ZFP91 binds to miR-519d-3p by a double luciferase assay. In addition, transfection of miR-519d-3p mimics in SW480 and HT29 cells decreased the mRNA and protein levels of ZFP91. In addition, in the tumor transplantation mouse model, we observed that the expression of ZFP91 was downregulated with the increase in miR-519d-3p levels. According to the correlation study, the ZFP91 expression state displayed a negative relationship with miR-519d-3p, but was positively associated with lncRNA EBLN3P in colon cancer specimens. Rescue experiments revealed that the colon cancer progression mediated by silencing lncRNA EBLN3P was reversed due to miR-519d-3p suppression, but these effects were neutralized due to ZFP91 attenuation. This intricate relationship between miR-519d-3p and ZFP91 not only elucidates a key regulatory mechanism in colon cancer progression but also highlights a potential therapeutic target. The negative correlation between miR-519d-3p expression and ZFP91 levels, along with the oncogenic role of ZFP91, suggests that interventions aimed at modulating this axis could offer new avenues for treatment. Specifically, strategies to enhance miR-519d-3p levels or to inhibit ZFP91 expression may disrupt this regulatory circuit, potentially impeding tumor growth and metastasis. Exploring targeted delivery mechanisms and creating inhibitors for ZFP91 may open new therapeutic pathways, enhancing the prospects for more effective colon cancer treatments.
This study acknowledges certain limitations, primarily the use of in vitro assays and a xenograft model that may not fully represent the human tumor microenvironment’s complexity. These models, while valuable for preliminary investigations, do not entirely capture the intricate cellular interactions and microenvironmental conditions present in human cancers. Consequently, the findings’ direct applicability to human clinical scenarios might be constrained. Future research could benefit from incorporating more physiologically relevant models, such as patient-derived organoids or advanced in vivo models, to enhance the translational potential of these findings.
In conclusion, this study identified that lncRNA EBLN3P accelerated cell proliferation and invasion of colon cancer by modulating the miR-519d-3p/ZFP91 axis. This discovery may provide useful information and therapeutic applications in colon cancer.
Authors’ Contributions
X.W.: Conceived and designed the experiments; Performed the experiments; Analyzed and interpreted the data; Wrote the article. Y.Y.: Analyzed and interpreted the data; Contributed reagents, materials, analysis tools or data; Wrote the article. J.T., F.K., B.S., and J.X.: Performed the experiments; Analyzed and interpreted the data; analysis tools or data. S.Y.: Analyzed and interpreted the data; Contributed reagents, materials, analysis tools, or data; Wrote the article.
Footnotes
Funding Information
This work was supported by the Science Foundation of Bayinguoleng Menggu Zizhizhou People’s Hospital (BHS201913), Special Scientific Research Project of Health Young Medical Science and Technology talents in Xinjiang Uygur Autonomous region (WJWY-202108), Tianshan Youth Program (2020Q144), the fifth batch of Gusu health personnel training project in Suzhou (GSWS2020085), Natural Science Foundation of Nanjing University of Chinese Medicine (XZR2020038), Suzhou Science and Technology Development Plan (SYSD2019213), and Science and Technology Innovation Project of Suzhou Medical and Health Care (SKJY2021136, SKY2023216).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation, to any qualified researcher.
Additional Information
No additional information is available for this article.
Disclosure Statement
The authors declare no conflict of interest.
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.