
Abstract
Background:
Long noncoding RNAs (lncRNAs), as emerging regulators of a wide variety of biological processes via diverse mechanisms, have been demonstrated to be of increasing importance in biology. Genome-wide association studies of tumor samples have identified several lncRNAs as either oncogenes or tumor suppressors in various types of cancers. In recent years, the importance of lncRNAs, especially in endometrioid cancer (EEC), has become increasingly well understood. The lncRNA Forkhead box P4 antisense RNA 1 (FOXP4-AS1) has been reported to fulfill roles in several types of cancers; however, the main biological function and associated underlying molecular mechanism of FOXP4-AS1 in EEC have yet to be fully elucidated. The present study therefore aimed to investigate how RNA FOXP4-AS1 may participate in the development and progression of endometrioid carcinoma tissues.
Materials and Methods:
In the present study, the expression level of FOXP4-AS1 was investigated in endometrioid carcinoma tissues and matching nearby normal endometrial tissues collected from patients receiving surgery at the hospital. A series of molecular biological assays were performed to investigate the effect of FOXP4-AS1 on cell proliferation, cell migration, and cell invasion.
Results:
An increased concentration of FOXP4-AS1 was identified in endometrioid carcinoma samples and cell lines compared with the corresponding controls, and this lncRNA was found to be positively correlated with advanced FIGO stages in patients with endometrial cancer. Furthermore, knocking down endogenous FOXP4-AS1 led to a significant reduction in the colony formation number and a significant inhibition of cell proliferation, cell migration, and cell invasion in endometrioid carcinoma cells. Moreover, dual-specificity phosphatase 5 (DUSP5), which is lowly expressed in endometrioid carcinoma tissues cells and negatively modulated by FOXP4-AS1, was identified as the downstream target molecule of FOXP4-AS1. Subsequently, the mechanistic experiments confirmed that, through binding to enhancer of zeste homolog 2 (EZH2; one of the catalytic subunits of polycomb repressive complex 2 [PRC2]), FOXP4-AS1 could epigenetically suppress the expression of DUSP5. Finally, the oncogenic function of the FOXP4-AS1/EZH2/DUSP5 axis in endometrioid carcinoma was confirmed via rescue assays.
Conclusions:
The findings of the present study have highlighted how FOXP4-AS1 fulfills an oncogenic role in endometrioid carcinoma, and targeting FOXP4-AS1 and its pathway may provide new biomarkers for patients with endometrioid carcinoma.
Introduction
Endometrial cancer is one of the most common gynecologic malignancies, ranking as the fourth most common cause of cancer in women; endometrioid cancer (EEC) accounts for 80%–90% of all cases. 1 The increasing rates of obesity, high blood pressure, and diabetes throughout the world have resulted in an increased incidence of endometrial cancer. Endometrial cancer is associated with a very high mortality rate at an advanced stage. 2,3 Even though great efforts have been made to reduce the damaging effects of the malignancy, the 5-year overall survival rate remains poor. 4 Consequently, the discovery of novel biomarkers and the drive to better understand the underlying molecular mechanism(s) of endometrial cancer are urgent tasks to be accomplished.
Long noncoding RNAs (lncRNAs) are categorized as a class of noncoding transcripts >200 nucleotides in length. 5 A burgeoning number of studies have shown that the majority of lncRNAs are involved in modulating the tumorigenesis and progression of numerous types of human cancer, 6 –8 including endometrial cancer. 9 –11 In recent years, the importance of lncRNAs in endometrial cancer has come to be understood in greater detail. 12,13 Forkhead box P4 antisense RNA 1 (FOXP4-AS1), an important lncRNA, has been demonstrated to have an oncogenic role in multiple types of solid cancers, including hepatocellular carcinoma, 14 osteosarcoma, 15 prostate cancer, 16 colorectal cancer, 17 esophageal squamous cell carcinoma, 18 and cervical cancer. 19 However, the biological function and related mechanisms of FOXP4-AS1 in EEC remain unknown.
One of the key roles of lncRNAs is to regulate gene transcription by directly interacting with epigenetic regulators. 20 Enhancer of zeste homolog 2 (EZH2) is not merely a catalytic subunit of polycomb repressive complex 2 (PRC2), but it also functions as a methyltransferase for histone H3 lysine 27 trimethylation (H3K27me3), 21 which leads to the promotion of epigenetic gene silencing. 22,23 Dual-specificity phosphatase 5 (DUSP5) has been reported to inhibit tumor progression as a negative regulator of extracellular signal-regulated kinase (ERK). 24,25 For example, ARAP1-AS1, which epigenetically silences DUSP5 expression, has been shown to promote cell proliferation and migration via recruiting EZH2 in cervical cancer. 26
The lncRNA CRNDE was shown to epigenetically suppress the expression of DUSP5 and CDKN1A through binding to EZH2, thereby promoting the development of colorectal cancer. 27 In addition, ROR1-AS1 was shown to inhibit the expression of DUSP5 via binding to EZH2, which influenced the proliferation and apoptosis of colon cancer cells. 28 However, to the best of the authors' knowledge, the expression profile, clinical characteristics, functional role, as well as the underlying targets of FOXP4-AS1 in endometrial cancer have yet to be characterized.
Endometrioid carcinoma accounts for the vast majority of endometrial cancer. The present study aimed to investigate the expression profile and regulatory mechanism of FOXP4-AS1 in EEC. The experiments performed demonstrated that FOXP4-AS1 was clearly upregulated in EEC, and a high expression level of FOXP4-AS1 predicted poor clinical outcomes. In addition, knocking down endogenous FOXP4-AS1 led to significant inhibition of the proliferative and migratory capabilities of EEC cells. Importantly, it is shown that FOXP4-AS1 was able to epigenetically silence DUSP5 expression, thereby promoting cell proliferation and migration through recruiting the EXH2 subunit of PRC2. Taken together, the results of the present study have shown that the lncRNA FOXP4-AS1 is able to promote the progression of EEC and therefore may serve as a viable target for treating EEC.
Materials and Methods
Patient tissue specimens
In total, 94 pairs of endometrioid carcinoma tissues and matching nearby normal endometrial tissues were collected from patients receiving surgery at the Fourth Affiliated Hospital of Hebei Medical University. None of the EEC patients had received any preoperative treatments before surgery. All tissue samples were confirmed by two independent histopathologists, stored in RNAlater® solution (Ambion; Thermo Fisher Scientific, Inc.) immediately after surgical removal, and kept at −80°C. The present study was approved by the research ethics committee of the Fourth Hospital of Hebei Medical University, and written informed consent was obtained from all the patients (approval No. 2021KY285). The clinical and pathological characteristics of the patients are summarized in Table 1.
The Relationship Between Expression of FOXP4-AS1 Gene and Clinicopathologic Features in Endometrioid Cancer Patients
a p < 0.05; b p < 0.001.
FOXP4-AS1, Forkhead box P4 antisense RNA 1.
Cell culture
The EEC cell lines KLE (No. CL-0133), RL95-2 (No. CL-0197), and Ishikawa (No. CL-0283), purchased from Procell Life Science & Technology Co., together with a normal endometrial cell line, were maintained in the laboratory and cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% Invitrogen® fetal bovine serum (FBS), 100 IU/mL penicillin, and 100 mg/mL streptomycin in a humidified atmosphere containing 5% CO2 at 37°C.
Regarding the culture of endometrial epithelial cells, endometrial samples (n = 10; average age of the patient = 38 years [range, 32–46 years]) were obtained at moment of hysterectomy for endometrial hyperplasia.
RNA isolation and reverse transcription–quantitative polymerase chain reaction
The total RNA used in the study was extracted from 1 cm3 tissue specimens and cancer cells using the TRIzol® method and then reverse-transcribed into complementary DNA (cDNA) using Invitrogen PrimeScript™ RT Master Mix (Thermo Fisher Scientific, Inc.). Reverse transcription–quantitative polymerase chain reaction (RT-qPCR) was subsequently performed following the instructions of the SYBR® Green Master Mix kit (Takara Bio, Inc.). The thermocycling conditions for the PCR were as follows: denaturation at 94°C for 15 s, followed by extension at 55°C for 30 s and then annealing at 72°C for 30 s, for a total of 40 cycles. The expression levels of the genes of interest were normalized against those of GAPDH, and all experiments were performed in triplicate. The fold change of the relative expression level was determined using the 2−ΔΔCq method, 29 and the sequences of the primers used in this study are shown in Supplementary Table S1.
Cell transfection
For cell transient transfection studies, the RL95-2 and Ishikawa cell lines were cultured at 37°C until the cells reached 50%–80% confluence. The transient transfection was conducted with final concentrations of 50 nM of small interfering RNAs (siRNAs) and 2.5 μg of overexpression plasmid using Invitrogen Lipofectamine2000™ (Thermo Fisher Scientific, Inc.) for incubation at 37°C and 5% CO2 for 6 h. Finally, the medium was replaced with RPMI-1640 supplemented with 10% FBS, following the manufacturer's protocol. After 48 h, cell suspension was harvested for RT-qPCR and subsequent cell function assays. The short hairpin RNAs (shRNAs) specific to FOXP4-AS1 (shFOXP4-AS1#1/2), along with the EZH2 overexpression plasmid (pcDNA3.1/FOXP4-AS1), were synchronously constructed by Shanghai GenePharma Co., Ltd. Nonspecific shRNAs and empty pcDNA3.1 vector were employed as negative controls, respectively. The sequences of the shRNAs used in this study are shown in Supplementary Table S2.
Cell proliferation assay
MTS cell viability assays (Promega Corporation) were performed strictly following the manufacturer's instructions. The Ishikawa and RL95-2 cells were placed into 96-well plates in triplicate, and the cells were subsequently incubated for 24, 48, 72, and 96 h with 100 μL RPMI-1640 containing 10% FBS; at the specified times, the cells were treated with 20 μL MTS solution and subsequently incubated for an additional 2 h at 37°C. The absorbance of each sample was then measured at 492 nm using a microplate reader. To investigate the impact of FOXP4-AS1 knockdown on the colony formation of Ishikawa cells and FOXP4-AS1 overexpression on the colony formation of RL95-2 cells, colony formation assays were performed.
First, cells were inoculated into 6-well plates with 2 mL RPMI-1640 culture medium containing 10% FBS, and the plates were subsequently incubated for 4–7 d in an incubator in an atmosphere containing 5% CO2. Subsequently, the cell colonies were fixed with 4% paraformaldehyde and subjected to crystal violet (2%) staining for 10 min at room temperature. Finally, colonies (>50) were counted and recorded manually. Cell clones were captured with a camera (EOS 3000D; Canon, Inc.), and the images were a close focus shot of each hole in a six-hole plate with the camera, without microscopic magnification.
Transwell migration and invasion assay
To investigate the migration and invasion of Ishikawa and RL95-2 cells, Transwell assays were performed according to the manufacturer's instructions. The cells were put into the upper chamber for cell migration assays, after with appropriate medium without 10% FBS added into the lower chamber. For the invasion assay, cells were seeded onto 50 μL 1 × Matrigel® Basement Membrane Matrix-coated (BD Biosciences; 1:3, at 37°C for 2 h) chambers. After cultivation for 24 h, the migrated or invaded cells adhering to the lower chamber were fixed, followed by staining with 0.1% crystal violet and counted under a light microscope (Olympus Corporation) at × 200 magnification, and cell counting was performed manually in five randomly selected visual fields. The assays were repeated in triplicate.
RNA pull-down assay
Pierce Magnetic RNA–Protein Pull-Down kits (Thermo Fisher Scientific, Inc.) were used for the RNA pull-down assays. FOXP4-AS1 and antisense FOXP4-AS1 were in vitro-transcribed using RiboMAX™ Large-Scale RNA Production Systems (Promega Corporation). Each binding reaction was incubated with streptavidin magnetic beads, followed by incubation at room temperature. Finally, the complexes bound to the beads were eluted and detected using standard Western blot analysis.
RNA immunoprecipitation assay
RNA immunoprecipitation (RIP) experiments were performed with Ishikawa and RL95-2 cells using Magna RIP™ RNA-binding protein immunoprecipitation kits, strictly following the manufacturer's instructions (Merck KGaA). Cells up to a density of 80%–90% were harvested, lysed in the RIP lysate buffer, and the lysates were cultivated with magnetic beads that were either bound to EZH2 or a negative control (IgG). Total RNA was extracted with TRIzol, after which RNA levels were determined using RT-qPCR assay, as described previously.
Chromatin immunoprecipitation assay
Chromatin immunoprecipitation (ChIP) assay was performed using the ChIP Assay Kit (Cat. No. P2078; Beyotime Biotechnology) according to the manufacturer's protocol. Ishikawa and RL95-2 cells (a total of 1 × 107 cells) were treated with 1% formaldehyde and incubated for 4 h time to produce DNA-protein cross-links and quenched with glycine for 10 min. The cell lysates were subsequently sheared into fragments (∼200–1000 bp in size) using ultrasound techniques (a 10 s on and 10 s off mode for 12 cycles on the ice) and immunoprecipitated either with anti-EZH2 antibody (6 μg, No. 5246s; Cell Signaling Technology) or with IgG as the negative control. After incubation with 60 μL Protein A + G Agarose (Beyotime Biotechnology) at 4°C for 1 h, DNA cross-linking was reversed by NaCl and protease K to purify DNA. Finally, the purified-DNA was accomplished using qRT-PCR after washing the beads.
Subcellular fractionation assay
The nuclear and cytoplasmic fractions of Ishikawa and RL95-2 cells were separated and purified using the Invitrogen PARIS™ Kit Protein and RNA Isolation System (Thermo Fisher Scientific, Inc.) by strictly following the manufacturer's instructions. The relative expression of FOXP4-AS1 in both nuclear and cytoplasmic fractions was then assayed using RT-qPCR.
Western blot analysis
Ishikawa and RL95-2 cells were treated with RIPA solution for total protein extraction, and protein concentration was evaluated using a Pierce BCA protein assay kit (Cat. No. PC0020; Beijing Solarbio Science & Technology Co., Ltd.). Protein samples (40 μg/lane) were then separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE; 10% gels), followed by transfer of the proteins onto a PVDF membrane (Cat. No. IPVH00010; Millipore). The membranes were then blocked with skimmed milk, incubated with primary antibodies overnight at 4°C, and subsequently incubated with anti-rabbit IgG antibody [dilution ratio of 1:1000; Cat. No. 075-1506; Seebio Biotech (Shanghai) Co., Ltd] for 1 h at room temperature.
Following washing with TBST, protein bands were visualized by incubating the membranes with ECL developer (Solarbio Biotechnology) in the dark, followed by detection using chemiluminescence imaging system of Syngene (Chemi-XT4; Syngene Co., Ltd). The immunoblots were probed with the following primary antibodies: anti-EZH2 (Cat. No. 5246; 1:1000; Cell Signaling Technology); anti-DUSP5 (Cat. No. ab232333; 1:2000; Abcam), and anti-GAPDH (ab181602; 1:2000; Abcam). Band intensity was semi-quantified and evaluated using the ImageJ software (1.48v; National Institutes of Health).
Bioinformatics tools
GEO (https://www.ncbi.nlm.nih.gov/geo/tools/profileGraph.cgi?ID=GDS4589:235891_at, GSE17025) and GEPIA (Gene Expression Profiling Interactive Analysis) data sets (http://gepia2.cancer-pku.cn/index.html) were used to explore FOXP4-AS1 expression in endometrial cancer samples. RPISeq data set (http://pridb.gdcb.iastate.edu/RPISeq) was performed to predict that FOXP4-AS1 could bind to EZH2, SUZ12, LSD1, DNMT1, SIRT1, and STAU1. GEPIA data set (http://gepia.cancer-pku.cn/index.html) was used to explore the correlation between FOXP4-AS1 and DUSP5 expression.
Immunohistochemistry staining
For immunohistochemistry (IHC) analysis, the tissue sections underwent deparaffinization and rehydration processes, followed by treatment with heat-induced epitope retrieval, blocked with bovine serum albumin (BSA), and the sections were then incubated overnight with a primary antibody against DUSP5 (1:100, No. 863746; Zenbio). Afterward, a secondary antibody was employed to incubate the sections for 1 h. Finally, DAB (3,3′-diaminobiphenyl) color development was carried out. A microscope (Nikon ECLIPSE, 50i) was utilized to capture images.
Statistical analyses
All statistical analyses were performed using SPSS 22.0 software (IBM, Corp.). Student's t-test or paired Student's t-test were performed to compare two groups, while comparisons among multiple groups were analyzed by using one-way analysis of variance followed by Tukey's multiple comparisons post hoc test. The association between FOXP4-AS1 expression and clinicopathologic features was analyzed using χ 2 test. The influential factors for survival were evaluated using univariate and multivariate Cox regression analyses. Kaplan–Meier analysis was used to assess the overall survival rate of the patients with EEC. All experimental data are expressed in this study as the mean ± standard deviation; p < 0.05 was considered to indicate a statistically significant value.
Results
FOXP4-AS1, upregulated in EEC, functions as an unfavorable prognostic factor in patients with EEC
Based searching the GEO (GSE17025) and GEPIA data sets (http://gepia2.cancer-pku.cn/index.html), the analysis revealed that FOXP4-AS1 was markedly upregulated in endometrial cancer samples (Fig. 1A, B). To verify the expression status of FOXP4-AS1 in EEC, 94 pairs of EEC and adjacent normal tissues were collected, and the expression level of FOXP4-AS1 was subsequently determined in the 2 groups. A noticeably higher expression level of FOXP4-AS1 was identified in the EEC tissues compared with that in adjacent nontumor tissues (Fig. 1C). Subsequently, to analyze the impact of FOXP4-AS1 expression on patients' survival, Kaplan–Meier survival curves were constructed, which revealed that there was a close correlation between high expression levels of FOXP4-AS1 and poor overall survival rates of patients with EEC (Fig. 1D).

FOXP4-AS1, upregulated in EEC, functions as an unfavorable prognostic factor in patients with EEC.
Subsequently, the analysis of the correlation between FOXP4-AS1 expression and the clinicopathologic features of the patients with EEC is shown in Table 1. The mean level of FOXP4-AS1 expression in the tumor tissues was used to allocate the cases into two groups, namely high and low FOXP4-AS1 expression groups. A relatively high level of FOXP4-AS1 was associated with advanced FIGO stage (p < 0.001), lymph node metastasis (p = 0.024), myometrial invasion (p = 0.048), and lymphovascular space invasion (p = 0.036), whereas no significant correlation was identified with age or histological grade (p > 0.05). Furthermore, with the adjustment of clinical parameters in Cox's proportional hazards regression model, both the univariate and multivariate analyses revealed that advanced FIGO stages (III+IV), lymph node metastasis, and a high level of FOXP4-AS1 expression were independent risk factors for the survival of patients with EEC (Table 2). Taken together, these results indicated that an elevated level of FOXP4-AS1 expression may be involved in the progression of EEC.
Univariate and Multivariate Analysis of Clinicopathologic Characteristics Influencing the Prognosis of Endometrioid Cancer Patients
p < 0.001; b p < 0.01; c p < 0.05.
CI, confidence interval; FOXP4-AS1, Forkhead box P4 antisense RNA 1; HR, hazard ratio.
FOXP4-AS1 promotes EEC cell proliferation, migration, and invasion
To observe the relevant functions of FOXP4-AS1 in EEC cells, functional assays were designed in two different EEC cell lines (Ishikawa and RL95-2). As shown in Figure 2A, RL95-2 cells displayed the lowest relative level of FOXP4-AS1 expression, whereas Ishikawa cells expressed FOXP4-AS1 at a higher level. To explore the underlying molecular mechanism and potential downstream targets of FOXP4-AS1 in EEC cells, the authors investigated its distribution in EEC cells, and FOXP4-AS1 was found to be predominantly located in the nuclei of EEC cells (p < 0.05) (Fig. 2B), suggesting the possible participation of FOXP4-AS1 in transcriptional regulation. Hence, the authors chose to overexpress FOXP4-AS1 in RL95-2 cells, whereas it was silenced in Ishikawa cells. Three different FOXP4-AS1 shRNAs and one overexpression plasmid were designed for transfection into the above-mentioned two cell lines of EEC. RT-qPCR analysis was subsequently applied to assess the transfection efficiency.

FOXP4-AS1 promotes EEC cell proliferation, migration, and invasion.
Of the three shRNAs, sh-FOXP4-AS1#1 exhibited the greatest silencing capability in Ishikawa cells. Meanwhile, the expression level of FOXP4-AS1 was increased in RL95-2 cell lines in comparison with the negative control following transfection with pcDNA-FOXP4-AS1 (Fig. 2C). Therefore, sh-FOXP4-AS1#1 and pcDNA-FOXP4-AS1 were selected in all subsequent experiments for silencing or overexpressing FOXP4-AS1, respectively. The impact of FOXP4-AS1 on the proliferation of EEC cells was subsequently evaluated via MTS and colony formation assays. As shown in Figure 2D and E, FOXP4-AS1 knockdown led to significantly attenuated viability of the Ishikawa cells (all p < 0.001), whereas overexpressing FOXP4-AS1 led to an increase in cell viability.
To detect whether FOXP4-AS1 expression could affect the migratory and invasive capabilities of the Ishikawa and RL95-2 cells, Transwell assays were performed. These experiments revealed that knocking down FOXP4-AS1 clearly reduced the numbers of both migratory and invasive cells compared with the shNC group in Ishikawa cells (Fig. 2F, G), whereas the migratory and invasive capabilities of the RL95-2 cells were increased via overexpression of FOXP4-AS1. These experimental findings suggested that FOXP4-AS1 is able to act as an oncogenic factor for EEC carcinogenesis through facilitating cell growth, proliferation, and invasion.
FOXP4-AS1 epigenetically silences the transcription of DUSP5 through interaction with EZH2
First, the probable RNA-binding proteins of FOXP4-AS1 were predicted through searching the RPISeq data set (http://pridb.gdcb.iastate.edu/RPISeq), which revealed that FOXP4-AS1 could bind to EZH2, SUZ12, LSD1, DNMT1, SIRT1, and STAU1 (Fig. 3A). RIP assay was subsequently performed to further explore the possibility of FOXP4-AS1's binding to the above proteins, which demonstrated that FOXP4-AS1 could interact with all these proteins, although the strongest binding occurred to EZH2 (Fig. 3B). The RNA pull-down assays also demonstrated the interaction of FOXP4-AS1 with EZH2 in endometrioid cancer cells (Fig. 3C). Collectively, these findings suggested that FOXP4-AS1 may epigenetically suppress downstream target genes through recruiting EZH2.

FOXP4-AS1 epigenetically silences DUSP5 transcription via recruiting EZH2.
Subsequently, potential targets of EZH2 (p53, CDKN2B, CDKN1A, KLF2, DUSP1, and DUSP5) were selected for further study. Analyzing the GEPIA data set (http://gepia.cancer-pku.cn/index.html) revealed a negative correlation between the expression of FOXP4-AS1 and CDKN1A, DUSP1, and DUSP5 expression (Fig. 3D), and, based on these findings, it was possible to speculate that these genes may be involved in EC development regulated by FOXP4-AS1. RT-qPCR was subsequently utilized to verify the changes in DUSP5, CDKN1A, and DUSP1 expression, and these experiments demonstrated that the expression of DUSP5, considered to be a tumor suppressor gene, was changed most prominently (Fig. 3E). Therefore, DUSP5 was selected as the research object for the following experiments.
Again, RT-qPCR and Western blot analyses were employed to accurately confirm the level of DUSP5 in the cells, and these assays showed that the level of DUSP5 was clearly increased in FOXP4-AS1-silencing Ishikawa cells, whereas it was decreased in FOXP4-AS1-overexpressing RL95-2 cells. Western blot assay was finally applied to further confirm the experimental results (Fig. 3F). Correspondingly, immunohistochemical methods were employed to assess the expression of DUSP5 protein in tissue samples with differential relative expression levels of FOXP4-AS1. As shown in Figure 3G, it demonstrated that the group with high FOXP4-AS1 expression exhibited decreasing DUSP5 protein expression. Taken together, the results above revealed that DUSP5 may be the downstream target molecule of FOXP4-AS1.
LncRNAs are usually involved in tumorigenesis via binding to specific RNA-binding proteins. Considering the evident binding of FOXP4-AS1 to EZH2, as demonstrated in the series of experiments in the present study, increased expression levels of DUSP5 were also found in si-EZH2-treated EC cells (p < 0.05) (Fig. 3H). Subsequently, the ChIP assays suggested that knocking down FOXP4-AS1 resulted in reduced EZH2 binding to the DUSP5 promoter (Fig. 3I). Essentially, these data revealed that FOXP4-AS1 could accelerate EC cell growth, partly through epigenetically silencing DUSP5 via binding to EZH2.
FOXP4-AS1 promotes the proliferation, migration, and invasion of EEC cells through silencing DUSP5
To verify the regulatory pattern of FOXP4-AS1 in EEC, rescue assays were designed and performed. The data from MTS and colony formation assays revealed that the suppression of cell proliferation that was mediated through the inhibitory effect of FOXP4-AS1 was reversed by the downregulation of DUSP5 (Fig. 4A, B). Transwell assays were subsequently conducted, and the results confirmed that downregulated FOXP4-AS1 could suppress cell migration, whereas its inhibitory role was eliminated through the silencing of DUSP5 (Fig. 4C). Taken together, these experiments showed that FOXP4-AS1 could promote the proliferation and migration of EEC cells through silencing DUSP5.
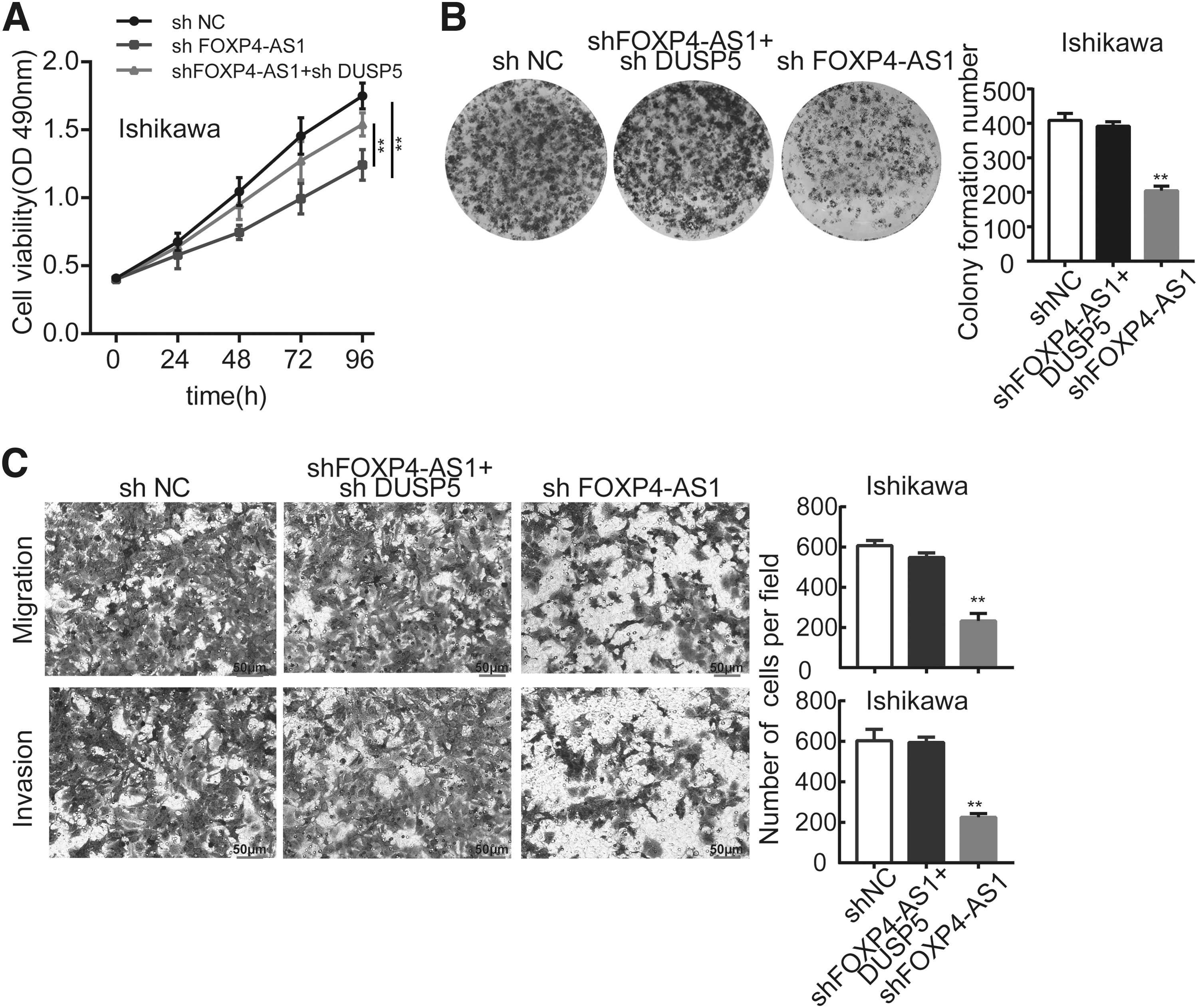
FOXP4-AS1 promotes the proliferation, migration, and invasion of EEC cells through silencing DUSP5.
Discussion
Endometrial cancer ranks first among gynecologic malignancies in terms of prevalence worldwide, endometrioid carcinoma as its main pathological type, 30 particularly demonstrating an upward trend in incidence in younger women all over the world. 31 Therefore, it is urgently required to establish more accurate preoperative staging and preoperative diagnosis of endometrial cancer, and this may be achieved through advances in the pathophysiological understanding of the malignancy and the development of novel clinical tumor markers of endometrial cancer.
In recent years, lncRNAs have shown great potential in the screening of novel tumor biomarkers and have been the focus of numerous studies. 32 Moreover, the number of lncRNAs has been identified as oncogenes or tumor-suppressive genes courtesy of the advancement of diagnostic techniques in endometrial cancer, although the underlying molecular mechanisms remain unclear in a lot of cases.
LncRNA FOXP4-AS1 has been reported to exert cancer-facilitating role in cancers. Emerging studies have demonstrated the multiple biological function of FOXP4-AS1, such as cell proliferation, metastasis, apoptosis, and so on. Moreover, abnormal expression of FOXP4-AS1 in tumor tissues can also serve as an effective prognostic biomarker for multiple solid cancers. 15 –17 In addition, a FOXP4-AS1 centered regulatory network was constructed by Wang et al. in hepatocellular carcinoma. 33 The underlying molecular mechanisms involved in FOXP4-AS1 interactions display diversity and complexity. Hence, exploring the potential mechanism pattern of FOXP4-AS1 in EEC is urgently necessary and would be beneficial to find new molecular targets for treating EEC.
Through searching the GEPIA database, lncRNA FOXP4-AS1 manifested a tendency to be overexpressed, a trend that was positively correlated with a worse prognosis of patients with EEC, indicating the association of FOXP4-AS1 with the progression of EEC. As anticipated, in the present study, an increased expression level of FOXP4-AS1 in EEC tissues was observed, and this was associated with an even worse prognosis of EEC. Functionally, when FOXP4-AS1 was knocked down, cell proliferation, migration, and invasion were inhibited, whereas overexpression of FOXP4-AS1 elicited the opposite effects on these cellular processes. The results of the present study are consistent with those of previous studies. Nevertheless, the present study, to the best of the authors' knowledge for the first time, has demonstrated the carcinogenic properties of FOXP4-AS1 in EEC development and progression.
Further investigating the precise biological function of FOXP4-AS1 in EEC should prove to be beneficial in terms of investigating novel promising clinical biomarkers and potential therapeutic targets. Considering that the role of FOXP4-AS1 as an oncogene in EEC has been validated, the goal of the present study was partly to shed light on the underlying molecular mechanism.
Several previous studies have shown that the vital role of lncRNAs in cancer is likely to be associated with their ability to affect cellular functions such as interacting with histone modification enzymes and mediating epigenetic alteration, also including the regulation of target gene expression. 34,35 For example, the repression of P21 and KLF2 expression transcription was shown to be mediated by the lncRNA HOXA-AS2 through its binding to EZH2 and LSD1 in colorectal cancer. 36 Linc00673 functions as an oncogene to accelerate cell proliferation and invasion, and the suppression of KLF4 expression was shown to be mediated through its interacting with EZH2 and DNMT1. 37 The lncRNA ROR1-AS1 facilitates colon cancer cell proliferation through inhibiting the expression of DUSP5/CDKN1A. 38 It is well known that lncRNAs usually bind to specific RNA-binding proteins that participate in tumorigenesis.
In the present study, RIP assay clearly demonstrated that FOXP4-AS1 was able to directly bind to EZH2. EZH2 is known to be one of the main effectors involved in epigenetic regulation; therefore, the assumption was that FOXP4-AS1 could regulate its target gene by interacting with EZH2.
DUSP5, which acts as a tumor-suppressor gene, is known to exercise roles in certain types of cancer; 39 –41 however, its role in endometrial cancer has yet to be fully elucidated. In the present study, it has been demonstrated that DUSP5 is the most obvious molecular target of EZH2. Initially, the expression level of DUSP5 was investigated in EEC cells, which showed that the expression of DUSP5 in EEC cells was low. Moreover, FOXP4-AS1 was found to be mainly localized in the nucleus of EEC cells. These findings encouraged us to further explore the association between FOXP4-AS1 and DUSP5.
According to the findings, it is assumed that FOXP4-AS1 may participate in epigenetic regulation, as has been confirmed by numerous studies on different cancer types. For example, the lncRNA CRNDE accelerates colorectal cancer cell proliferation through epigenetically suppressing DUSP5/CDKN1A expression. 42 lncRNA ARAP1-AS1 promotes cervical cancer cell proliferation and migration via epigenetic regulation of DUSP5. 40 Additional mechanistic experiments were conducted to validate the hypothesis. In the present study, it was found that DUSP5 acts as a downstream target of FOXP4-AS1 in EEC cells. Furthermore, it was possible to confirm that FOXP4-AS1 could recruit EZH2 to the promoter of DUSP5 and then modulate the transcriptional expression of DUSP5. Finally, rescue assays were performed to confirm that FOXP4-AS1 was able to aggravate the cellular progression of EEC through epigenetically silencing DUSP5.
In conclusion, the present study has shown that the expression level of FOXP4-AS1 was increased in EEC tissues and that this was associated with poor prognosis of EEC patients. Furthermore, the mechanism experiment has demonstrated that FOXP4-AS1 may enhance the progression of EEC through binding to EZH2 and inhibiting the expression of DUSP5, which is considered a tumor suppressor.
Ethics Approval and Consent to Participate
This study was approved by the ethics committee of the Fourth Affiliated Hospital of Heibei Medical University (approval No.2021KY285) and complied with the Declaration of Helsinki. All patients provided written informed consent.
Patient Consent for Publication
Not applicable.
Availability of Data and Materials
The data sets used or analyzed during the present study are listed in the article.
Footnotes
Acknowledgments
The authors acknowledge the doctors at the Department of Obstetrics and Gynecology, Hebei Medical University, Fourth Hospital, for their assistance in recruiting study subjects.
Authors' Contributions
L.L., Z.Z., and L.S. designed and revised the article. J.J. and J.Z. recruited the patients and collected the data. H.G., J.M., and L.L. performed the experiments, data collection and analysis, statistical analysis, and article drafting. All authors have read and approved the final article.
Disclosure Statement
The authors declare no potential conflicts of interest.
Funding Information
This study was supported by Grants from the Cancer Foundation of China (Grant No. LC2016W11) and the Scientific Research Fund of Hebei Provincial Health Commission (Grant No. 20200227).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.