
Abstract
Objective:
Hepatocellular carcinoma (HCC) is a highly lethal cancer with significant mortality, primarily attributed to metastasis. Although Protocadherin Gamma Subfamily A, 9 (PCDHGA9) has been identified as a tumor suppressor gene in cancer metastasis, its role in HCC remains ambiguous. This study clarifies the role of PCDHGA9 in HCC by examining its expression, clinical significance, and molecular activities.
Methods:
Tissue microarray immunofluorescence analysis evaluated the expression of PCDHGA9 and its clinical relevance. In vitro experiments involved manipulating PCDHGA9 levels in SK-HEP-1 cells to assess migration through wound-healing and transwell assays. In vivo, shPCDHGA9 cell injections were utilized to observe effects on tumor growth and metastasis. Protein analysis and Western Blot validated epithelial-mesenchymal transition (EMT)-related proteins. Subsequent to TGF-β treatment, cell proliferation and apoptosis were quantified using cell counting kit-8 and flow cytometry, respectively, followed by investigation of TGF-β effects on PCDHGA9 N6-methyladenosine (m6A) modification via Methylated RNA immunoprecipitation, RT-qPCR, and Western blot analysis.
Results:
Downregulation of PCDHGA9 expression in HCC tissues is correlated with poor prognosis. In vitro experiments demonstrated that modulating PCDHGA9 expression influenced HCC cell migration. In vivo, PCDHGA9 knockdown is correlated with increased metastasis. Furthermore, TGF-β stimulation promoted cell proliferation and inhibited apoptosis. Mechanistically, TGF-β-mediated m6A modification led to PCDHGA9 decay, promoting EMT in HCC cells.
Conclusion:
PCDHGA9 serves as a potential tumor suppressor in HCC by inhibiting EMT. During this process, TGF-β is observed to exert regulatory control over m6A modifications of PCDHGA9.
Introduction
Hepatocellular carcinoma (HCC) is a highly aggressive malignant neoplasm, ranking sixth in incidence and third in cancer-related mortality globally. 1,2 With an insidious development and rapid progression, HCC is often diagnosed in the advanced stages, leading to high recurrence rates, rapid metastasis, poor prognosis, and elevated mortality rates. 3 –5 Therefore, elucidating the molecular mechanisms underlying HCC tumorigenesis and metastasis may provide a better understanding of its pathogenesis and offer potential targets for the treatment of this lethal disease.
The epithelial-mesenchymal transition (EMT) is a critical biological process linked to metastasis in solid tumors, including HCC. 6,7 During EMT, cancer cells transition from epithelial to mesenchymal characteristics, involving the loss of E-cadherin and the acquisition of vimentin. 7,8 Although traditional calcium mucins like E-cadherin and N-cadherin have been extensively studied in relation to tumor EMT, protocadherin (PCDH) has received less attention. Nonetheless, ample evidence suggests that inactivation of PCDH may enhance tumor cell metastasis. 9,10 PCDHs are part of the cadherin superfamily, divided into clustered and non-clustered subgroups. 11 Aberrant expression of PCDHs in various cancers and their potential role in cancer regulation have attracted research interest. Several PCDHs, known as antitumor genes, are often downregulated because of methylation. 12,13 For example, PCDH10 is downregulated through hypermethylation or gene deletion in various malignancies including HCC, colorectal cancer, gastric cancer, and pancreatic cancer, correlating with tumor progression and metastasis. 14 In liver cancer, non-clustered members of PCDHs, such as PCDH20, 15 PCDH8, 12 and PCHD9, 16 are frequently downregulated because of promoter hypermethylation, and exert a tumor suppressor effect. Protocadherin Gamma Subfamily A, 9 (PCDHGA9), a cluster PCDH, plays a crucial role in intercellular adhesion and nuclear signaling. 17 Studies have shown that deletion of PCDHGA9 promotes EMT in gastric cancer, indicating its direct interaction with the TGF-β/Smad2/3 signaling pathway. 18 Reportedly, the TGF-β signaling pathway regulates the development of HCC, yet its regulatory mechanisms remain complex. 19 To our knowledge, the role and functionality of PCDHGA9 in HCC, as well as its relationship with TGF-β, have not been reported.
N6-methyladenosine (m6A) is a common modification found in mRNA that plays a crucial role in maintaining its stability. 20,21 There is increasing evidence suggesting that RNA m6A modification is involved in the regulation of HCC cells during the process of EMT. 21,22 Downregulation of PCDHGA9 because of promoter hypermethylation has been linked to a poor prognosis in gastric cancer patients. 18 However, the association between PCDHGA9 and mRNA methylation m6A has been scarcely reported. The primary objective of this study was to examine the expression levels of PCDHGA9 in HCC and its potential impact on tumor progression. Specifically, the study involved both overexpression and knockdown of PCDHGA9 in SK-HEP-1 cells to observe changes in cell migration and EMT. Furthermore, PCDHGA9 knockdown was carried out in an in vivo setting to assess its effects on tumor growth and metastasis. Moreover, the study delved into exploring the relationship between PCDHGA9 m6A modification and TGF-β.
Material and Methods
Patients and immunofluorescence on tissue microarrays
Tissue microarrays consisting of 85 liver cancer tissues and their corresponding paraneoplastic tissue specimens were procured from Shanghai Xinchao Biotechnology (Shanghai, China) and approved by the ethics committee (approval number: YBM-05-02). Immunofluorescence experiments were performed on the liver cancer tissue microarray at Servicebio (Wuhan, China) using an anti-PCDHGA9 antibody (MA5-27615) purchased from Thermo Fisher Scientific (Waltham, MA). Apipathwell software was used to analyze the fluorescence signals of the tissue microarray samples to determine the average optical density (OD) and positivity rate of each sample. The PCDHGA9 expression level score was calculated as the product of the mean OD score and the positive rate score. To assign the mean OD score, 170 cancer and paracancerous tissue samples from 85 patients were ranked according to their OD values, with the first 5% scored as 0, 5%–25% as 1, 25%–50% as 2, 50%–75% as 3, and the last 25% as 4. The positive rate score was assigned based on the percentage of positive cells, with a score of 0 for positive rate <5%, 1 for 5%–25%, 2 for 25%–50%, 3 for 50%–75%, and 4 for positive rate >75%. We collected and analyzed tumor volume and survival data in the 85 patients based on their PCDHGA9 expression level score, with a score <6 defined as low PCDHGA9 expression and ≥6 as high PCDHGA9 expression. Another set of 15 pairs of freshly frozen HCC tissue and normal tissue specimens were utilized for the validation of PCDHGA9 methylation. These specimens were obtained from the Henan Provincial Cancer Hospital and approved by the institutional review committee (Approval No. 2016CT054).
Cell culture
The human HCC cell line SK-HEP-1, obtained from the ATCC (Manassas, VA), was maintained in Minimum Essential Medium (MEM, GIBCO BRL, Grand Island, NY). The culture was maintained at 37°C with 5% carbon dioxide (CO2) in a constant temperature and humidity incubator. Only cells in good cellular condition were used for the experiments. Human recombinant TGF-β1 was procured from R&D Systems (Minneapolis, MN). To investigate the impact of TGF-β on PCDHGA9 and tumor malignancy progression, cells were treated with 5 ng/mL of TGF-β1, with phosphate-buffered saline (PBS) serving as the control.
RNA interference and overexpression vector construction and transfection
The PCDHGA9 lentiviral interference vector was constructed and virally packaged at VectorBuilder (Guangzhou, China). The interfering sequence is as follows: NM_018921.3: CCCAAATTCTTGACCGAGAAA, vector VB210901-1375sam. PCDHGA9 overexpression vector information is as follows: pLV[Exp]-EGFP:T2A:Puro-EF1A>hPCDHGA9 [NM_018921.2], control vector information is: pLV[Exp]-EGFP/Puro-EF1A>ORF_stuffer. SK-HEP-1 cells were transfected in six-well plates at a density of 1.65 × 105 cells per well, and the experimental setup was grouped into oePCDHGA9 group, oeNC group, shPCDHGA9 group, and shNC group. The transduced MOI used in the experiment was ∼10. 25 uL of shPCDHGA9 and oePCDHGA9 virus solution (original virus titer of 1 × 108/mL) was added to 2 mL of medium and mixed, and 10 uL of shNC and oeNC virus solution (original virus titer of 2 × 108/mL) was added to 1 mL of medium and mixed, respectively.
For METTL3 and YTHDF2 siRNA transfection, the siRNA sequences employed in this investigation were custom-designed and chemically synthesized by RiboBio as follows: for human METTL3, 5ʹ-GCACTTGGATCTACGGAAT-3′ʹ; for human YTHDF2, 5ʹ-CTGCCATGTCAGATTCCTA-3′ʹ; nontargeting scrambled siRNA served as the control. Transfection of SK-HEP-1 cells was executed using Lipofectamine 3000 (Invitrogen), following the manufacturer’s instructions.
Transwell migration assay
The cells were incubated at 37°C with 5% CO2 for 48 h. The liquid was aspirated from the chambers, and the nonmigrating cells from the membrane upper surface were removed using a sterile cotton swab and fixed with 4% paraformaldehyde, stained with 0.1% crystal violet, rinsed with PBS, and photographed under a light microscope. The cells were observed under Olympus CKX53 inverted microscope and photographed.
Wound-healing assay
The cells of each group were spread at a density of 1 × 105/well into a 6-well plate with five parallel lines crossed vertically at the bottom, and after the cells were completely spread across the bottom, the pipette tips were used to score along the bottom of the wells in a straight line. According to the cell proliferation rate, appropriate time points were selected for light microscopic observation and photographed to detect cell migration.
Cell counting kit-8 (CCK-8) assay
Cells were seeded at a density of 5 × 103/well in a 96-well plate and treated with either 5 ng/mL TGF-β1 or PBS as control at 0, 12, 24, 48, and 72 h. Subsequently, 10 μL of 10% CCK-8 solution (Beyotime, Shanghai, China) was added to each well, followed by further incubation at 37°C for 1 h. The OD values of each well were measured at a wavelength of 450 nm using a microplate reader (Thermo Fisher Scientific) to assess cell viability.
Flow cytometric analysis
Cells were seeded in six-well plates, harvested by centrifugation at 1500 rpm for 5 min, washed three times with 1×PBS, and then incubated in the dark at room temperature with 5 μL of FITC-labeled Annexin V and 5 μL of propidium iodide (PI) for 10 min. Flow cytometric analysis was performed using a flow cytometer and analyzed with FlowJo software (BD Biosciences, CA).
Tumor-bearing mice experiment
The animal experiments were conducted in accordance with the guidelines of the Institutional Animal Care and Use Committee of the Guangdong Institute of Applied Biological Resources (20190312). Four-week-old male BALB/c nude mice (SPF-grade, weighing 15–19 g) were procured from Spbeifu (Beijing) Biotechnology Co., Ltd. For the intraperitoneal injection model, SK-HPE-1 cell lines were expanded in culture, and shPCDHGA9 and control shNC cells were collected at a concentration of 2.5 × 107/mL in prechilled PBS. Each mouse was intraperitoneally injected with 100 μL of the respective cells. In the transsplenial injection model, BALB/c nude mice were anesthetized, and a small incision was made in the skin near the spleen. The spleen was gently pulled out using sterile forceps, and a volume of 5 × 107/50 μL tumor cells was inoculated into 1 mL of insulin. The muscle and skin were then sutured, and the mice were kept warm at 37°C until they regained consciousness. After 1 month, euthanasia was performed on all murine subjects, followed by the creation of a transverse abdominal incision, and photographic documentation of intratumoral growth and metastatic manifestations was undertaken.
Proteomics mass spectrometry
The shPCDHGA9 and shNC control SK-HEP-1 cell lines were submitted for proteomic mass spectrometry analysis at Lianchuan Biologicals (Zhejiang, China). The protein extraction and reductive alkylation were performed as per standard protocols. Trypsin was added to digest the proteins overnight at 37°C. Tandem Mass Tag labeling, a widely used in vitro peptide labeling technique developed by Thermo Fisher Scientific, was carried out following the manufacturer’s instructions. High-performance liquid chromatography was used to separate the labeled peptides, utilizing a Shimadzu LC20AD chromatograph system and a Waters XBridge Shield C18 RP column. The separated peptides were subjected to liquid tandem mass spectrometry, using a Shimadzu LC20AD chromatograph system, Waters XBridge Shield C18 RP column, and nanoliter UPLC Dionex Ultimate RSLCnano 3000 system. MaxQuant software (v.1.5.2.8, http://www.maxquant.org/) was employed to resolve the mass spectrometry data. The UniProt-GOA database (http://www.ebi.ac.uk/GOA/) was used for Gene Ontology analysis, and the proteins were classified based on their cellular composition, molecular function, or physiological processes. Fisher’s exact test was used to identify differentially expressed proteins. The Gene Ontology enrichment tests were performed, and p-values <0.05 were considered statistically significant.
qRT-PCR analysis
The extraction of total RNA was carried out employing TRIzol reagent (Invitrogen) as per the manufacturer’s guidelines. Subsequently, the reverse transcription of 2 μg of RNA was conducted using the HiScriptII Q RT SuperMix for qPCR (+gDNA wiper) (R223-01) (Vazyme, Nanjing, China) in accordance with the provided instructions. RT-PCR was executed utilizing the ChamQ Universal SYBR qPCR Master Mix (Q711-02/03) (Vazyme). For the qRT-PCR analysis, specific primers were employed: PCDHGA9 (forward: 5′-TCATATAACCCCGTCAACGC-3′, reverse: 5′-CAAATCCAGCAAGAAGTCCC-3′) and GAPDH (forward: 5′-GGGAAGGTGAAGGTCGGAGT-3′, reverse: 5′-GGGGTCATTGATGGCAACA-3′). Each sample underwent triplicate analysis, and the data were normalized to GAPDH. Subsequently, the relative PCDHGA9 mRNA levels were calculated using the 2-ΔΔCt method.
Western blot analysis
Cell lysis for protein extraction was performed utilizing RIPA buffer (Beyotime Biotechnology, Jiangsu, China). Protein concentration quantification was performed following the guidelines provided in the manufacturer’s protocol for the BCA protein assay kit (Beyotime). Equal quantities of protein (30 μg) were resolved on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) matrices and subsequently transferred onto polyvinylidene difluoride (PVDF) membranes (Millipore, Billerica) using established procedures. The membranes underwent successive blocking in 5% skim milk within a tris-buffered saline and tween 20 (TBST) solution for 1.5 h at room temperature, followed by an overnight incubation at 4°C with primary antibodies. Subsequent to incubation with a secondary antibody for 1 h at room temperature, protein visualization was accomplished using ECL reagent (Millipore, Billerica). Primary antibodies specific to anti-GAPDH (1:1000, Santa Cruz, CA), PCDHGA9 (1:500, Abcam, London, UK), E-cadherin (1:800, Cell Signaling Technology), N-cadherin (1:1000, Abcam), ZEB1 (1:1000), TGFB1I1 (1:1000), Mettl3 (1:1000), Mettl14 (1:500), Fto (1:1000), Alkbh5 (1:1000) (all from Proteintech Group, China).
Methylated RNA immunoprecipitation (MeRIP)
Immunoprecipitated magnetic beads of total RNA were routinely prepared and closed in sheep antirabbit IgG suspension prepared at 4°C for 2 h. The RNA was combined with m6A antibody beads in a head-to-tail rotation, and the coprecipitated RNA was separated after elution, i.e., the RNA sample after MeRIP was obtained, and this RNA was subjected to RT-qPCR detection as described in the previous steps.
Actinomycin D experiment
SK-HEP-1 cells transfected with METTL3 siRNA, YTHDF2 siRNA, or control siRNA were subjected to treatment with 2.5 mg/L of actinomycin D for various durations (0, 3, and 6 h). Following treatment, cells were washed with PBS, and total RNA was extracted for the assessment of mRNA levels of the PCDHGA9 gene within the cells using RT-qPCR.
Statistical analysis
SPSS Statistics 22 and GraphPad Prism 8 were used for analysis, and normally and near-normally distributed measures were expressed as x¯±s, and independent samples t-test was used for comparison of two groups, and ANOVA was used for comparison of multiple groups. Non-normally distributed measures were expressed as medians (quartiles) and compared using the Mann–Whitney U test or Kruskal–Wallis H test. Paired rank sum test was used for cancerous and paraneoplastic tissues, and Kaplan–Meier analysis of the relationship between PCDHGA9 expression and patient prognosis was performed by log-rank test for comparison between groups. The p < 0.05 was considered a statistically significant difference.
Results
PCDHGA9 expression is down-regulated in liver cancer tissues
The statistical results of liver cancer tissue microarray showed that PCDHGA9 expression was significantly downregulated in liver cancer tissues (Fig. 1A), and its expression level was significantly correlated with tumor tissue size (Fig. 1B). In addition, PCDHGA9 was significantly correlated with patient survival in liver cancer patients with survival >2 years (Fig. 1C).

PCDHGA9 downregulation in HCC.
PCDHGA9 inhibits the migration of SK-HEP-1 cells
To investigate the function of PCDHGA9 in HCC development, shPCDHGA9 lentiviral interference vector and overexpression vector were constructed, and SK-HEP-1 cell lines stably interfering or overexpressing PCDHGA9 were screened (Fig. 2). Transwell and wound-healing assays were performed using this cell line, showing that downregulation of PCDHGA9 promoted migration of SK-HEP-1 cells and, conversely, inhibited migration of SK-HEP-1 cells (Fig. 3).
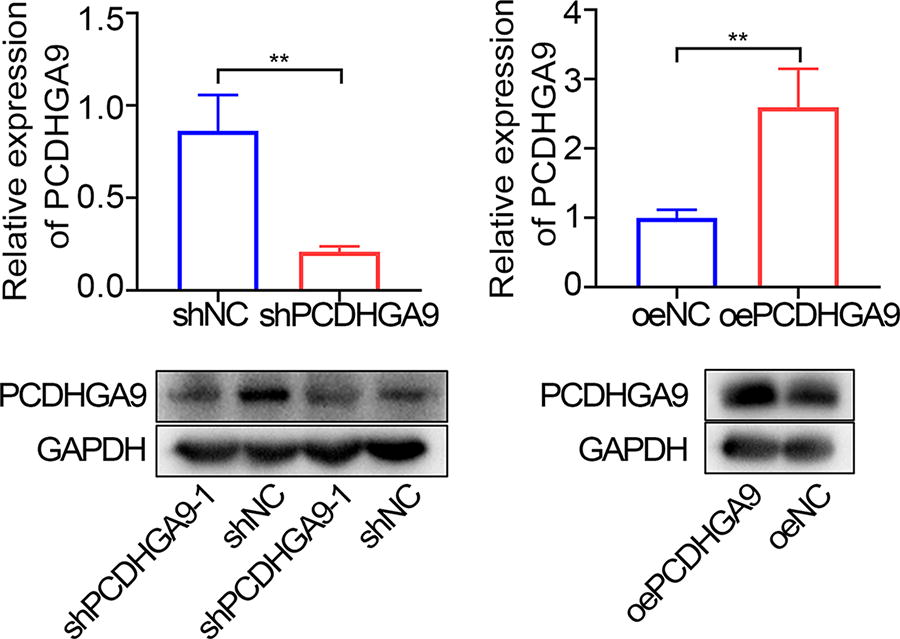
Validation of PCDHGA9 interference or overexpression in SK-HEP-1 cells. Western blotting and RT-qPCR analysis of PCDHGA9 expression levels in SK-HEP-1 cells after knockdown or overexpression of PCDHGA9. **p < 0.01. n = 3. PCDHGA9, protocadherin Gamma Subfamily A, 9.
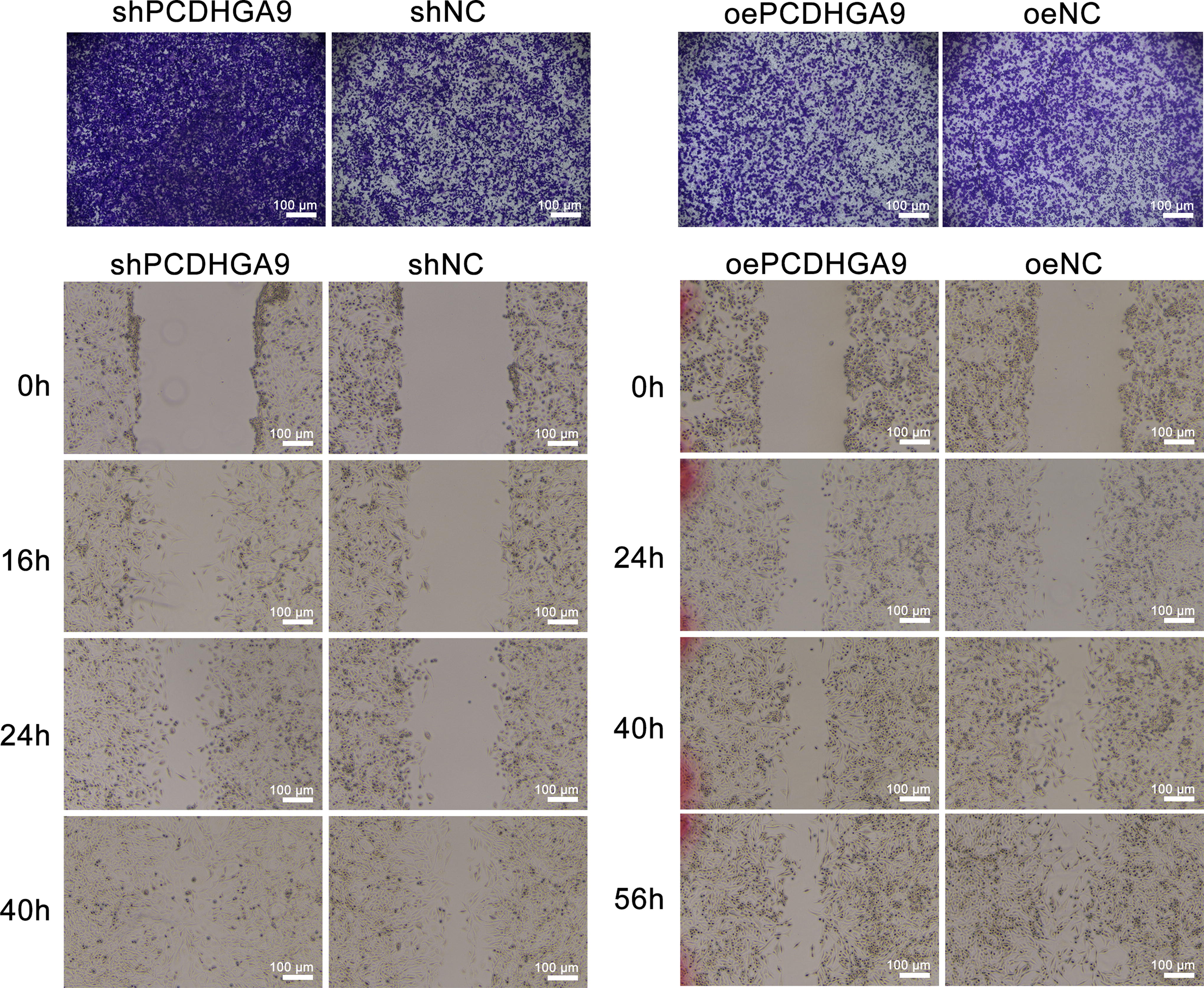
PCDHGA9 promotes migration of HCC cells in vitro. Cell migration ability after PCDHGA9 knockdown or overexpression was assessed using wound healing and Transwell assays. n = 3. HCC, Hepatocellular carcinoma; PCDHGA9, protocadherin Gamma Subfamily A, 9.
PCDHGA9 inhibits tumor proliferation and metastasis in vivo
After the inhibition of SK-HEP-1 cell migration by PCDHGA9 cells in vitro was clarified, its function was further verified in vivo using BALB/c nude mice subsequently. The results showed that mice injected intraperitoneally with shPCDHGA9 SK-HEP-1 cells had large tumor tissues in the thoracic septal junction, severe abdominal spleen, pancreas, and stomach adhesions, and large tumor volumes; control SK-HEP-1 mice had smaller tumor areas in the thoracic septal junction and smaller abdominal tumors (Fig. 4A). Mice injected with shPCDHGA9 SK-HEP-1 cells via spleen had large-volume tumors in the liver, with severe cirrhosis or kidney involvement in individual mice, whereas control mice showed small-volume single or multiple tumor foci in the liver (Fig. 4B). The results indicated that PCDHGA9 inhibited tumor proliferation on the one hand and tumor metastasis on the other in vivo.

PCDHGA9 promotes migration of hepatocellular cancer cells in vivo. Observation of in vivo tumor profiles in BALB/c nude mice after intraperitoneal
PCDHGA9 inhibits the EMT process
Mechanistically, proteomic analysis using shPCDHGA9 and shNC SK-HEP-1 cells revealed that PCDHGA9 knockdown had significant effects on EMT pathway-related molecules such as TGF1I1, TGFB2, and COL1A1 in HCC cells, while WB experiments also showed that EMT-related molecular markers such as ZEB1, N-cadherin, and TGF1I1, THF1I1 expression were upregulated and E-cadherin expression was downregulated after PCDHGA9 knockdown, while the opposite was true after PCDHGA9 overexpression (Fig. 5).

PCDHGA9 inhibits EMT in HCC.
TGF-β stimulation upregulates the level of m6A modification of PCDHGA9
Since PCDHGA9 regulates the EMT process, it is pertinent to investigate whether the classical cytokine TGF-β, 23 which induces EMT at the cellular level, exerts a regulatory effect on PCDHGA9. Cell proliferation and apoptosis in tumor cells are pivotal indicators of malignant tumor progression. Subsequently, we conducted CCK-8 and flow cytometry assays to assess the impact of TGF-β1 stimulation on cell proliferation and apoptosis. The results of the CCK-8 assay revealed a significant increase in proliferation of SK-HEP-1 cells following treatment with 5 ng/mL TGF-β1 for 12, 24, 48, and 72 h (Fig. 6A). Flow cytometric analysis demonstrated that the percentage of apoptotic cells induced by TGF-β1 stimulation was markedly lower compared to the PBS control group (Fig. 6B). Furthermore, TGF-β stimulation significantly down-regulated PCDHGA9 mRNA levels and protein levels but did not affect the distribution of PCDHGA9 in the cytoplasmic nucleus (Fig. 6C, D). It is hypothesized that PCDHGA9 mRNA underwent m6A modification after TGF-β stimulation, leading to a decrease in its stability and an increase in degradation. As seen from WB, the expression of METTL3 and METTL14 in m6A modification protein complex increased after TGF-β stimulation (Fig. 6E), suggesting that TGF-β stimulation led to an upregulation of the overall m6A modification level in SK-HEP-1 cells. The m6A RIP experiments further verified that the m6A modification level of PCDHGA9 was indeed significantly upregulated after TGF-β stimulation (Fig. 6F). Using actinomycin D, it was demonstrated that the stability of PCDHGA9 significantly increased upon interference with Mettl3 and YTHD2, which have been reported as the m6A modification methyltransferase (writer) and binding protein (reader), respectively (Fig. 6G). 24 Finally, m6A IP was performed on cancer and paraneoplastic tissues from 15 pairs of HCC patients, and 11 patients exhibited an upregulation of PCDHGA9 m6A modification levels in cancer tissues (Fig. 6H). It was shown that TGF-β does have a regulatory effect on PCDHGA9.

TGF-β stimulation upregulates the level of m6A modification of PCDHGA9.
Discussion
The development of HCC is a complex multistep process involving the activation of oncogenes and the silencing of tumor suppressor genes. Previous studies have demonstrated that certain non-clustered PCDHs members can inhibit EMT and cell migration in HCC. 16 Abnormal expression of PCDHGA9 has been observed in various cancers with poor prognoses, such as gastric cancer. 17,18 In this study, we demonstrated that PCDHGA9 expression is significantly lower in HCC tissues than in corresponding normal tissues, and Kaplan–Meier analysis suggests that PCDHGA9 may play a crucial role in the pathogenesis and prognosis of HCC. In addition, proteomic analysis and tumor-bearing mice results indicated that knockdown of PCDHGA9 expression caused changes in EMT-associated proteins and promoted metastasis, which was consistent with the described function of PCDHGA9 in gastric cancer. 17 Although not extensively studied in solid tumors, our findings and other reports suggest that PCDHGA9 and possibly other clustered PCDH family members share a common function of inhibiting tumor metastasis.
Several members of the PCDHs (PCDH10, 17, and 20) have been reported to be inactivated by promoter methylation in human cancers. The data of Zhang et al. 12 demonstrate that PCDH8 plays an important role in the carcinogenesis of liver cancer and is often inactivated by promoter methylation. Lv et al. 15 showed that deletion and/or aberrant epigenetic modulation of PCDH20 gene in HCC tumor specimens was responsible for its downregulation. These results suggest that epigenetic silencing of PCDHs may be a factor in the carcinogenesis of HCC. Weng et al. 17 showed that PCDHGA9 was down-regulated in GC tissues and GC cell lines and that this down-regulation was accompanied by hypermethylation of the PCDHGA9 promoter. In the present study, mRNA methylation m6A was promoted as the main regulator of PCDHGA9 level, after excluding transcriptional level regulation.
Furthermore, one of the acknowledged functions of TGF-β is to trigger EMT, 25 which plays a pivotal role in the onset and progression of HCC. 19 Previous studies have reported that the TGF-β signaling pathway can regulate m6A RNA methylation. 26 TGF-β2 induces differential m6A and RNA levels in cells, and m6A methylation may regulate the expression of EMT markers. 27 Similarly, m6A RNA methylation plays a crucial role in TGF-β-induced EMT in lung cancer cells. 28 It is well-established that dysregulated m6A methylation is a factor in the tumorigenesis and metastasis of various cancers, partially attributed to the upregulation or downregulation of three types of regulatory factors: “writers,” “erasers,” or “readers.” 21,28 Previous research has demonstrated that m6A writer (METTL3) and reader (YTHD2) participate in many pathological processes, including cancer. 21,28 Consistent with this, destabilization of PCDHGA9 significantly increases upon interference with Mettl3 and YTHD2, suggesting that m6A-mediated regulation of PCDHGA9 can be attributed to the distinct functions of these m6A writer and reader proteins. This is the first time that TGF-β stimulation upregulated PCDHGA9 m6A modification was disclosed, which further promoted the decay of PCDHGA9.
Conclusion
In conclusion, this study provides novel evidence that PCDHGA9 is a promising tumor suppressor gene that inhibits EMT in HCC. The significant upregulation of the m6A modification level of PCDHGA9 after TGF-β stimulation indicates that this epigenetic modification may play a critical role in hepatocarcinogenesis. Our findings shed new light on the role of PCDHs in HCC and suggest that PCDHGA9 may serve as a potential therapeutic target for HCC.
Footnotes
Ethics Statement
The studies involving human participants were reviewed and approved by the Ethics Committee of Henan Cancer Hospital (no.2016CT054). The patients provided their written informed consent to participate in this study. The animal management and utilization plan has been approved by the ethics committee of Guangdong Provincial Institute of Biological Resources (20190312) and strictly adheres to the Guangdong Experimental Animal Management Regulations.
Authors’ Contributions
All authors contributed to the study conception and design. Material preparation, data collection, and analysis were performed by W.G., X.Z., Y.W., Y.O., Y.Z., Z.Z., and H.Y. The first draft of the article was written by H.W. and all authors commented on previous versions of the article. All authors read and approved the final article.
Data Availability
The data supporting the findings of this study are available from the corresponding author upon reasonable requests.
Authors Disclosure Statement
The author reports no conflicts of interest in this work.
Funding Information
The present study was supported by the National Natural Science Foundation of China (81902891), Guangdong Basic and Applied Basic Research Foundation, Natural Science Foundation of Guangdong Province (2019A1515011456), GDAS Special Project of Science and Technology Development (2020GDASYL-20200202001, 2022GDASZH-2022020402-01), 2021TQ060920, and Guangdong Province Overseas Master Project (2021).