
Abstract
Abstract
The present article reviews master stem cell transcription factors, their expression regulation network, and related signaling pathways with the aim of understanding the molecular mechanisms of pluripotent cell fate decisions. Oct4, Sox2, and Nanog are master transcription factors for maintenance of the undifferentiated state and self-renewal of embryonic stem cells (ESCs). In the mouse, they form a regulatory circuitry with coregulators, such as β-catenin, Stat3, Myc, Klfs, Sall4, and Esrrb to control the expression of pluripotency-related genes including themselves. The threshold expression of Oct4, Sox2, and Nanog for sustaining ESC properties depends on the synergistic effects among Stat3, β-catenin, and Smad signaling pathway under the specific conditions of the ESC cytoplasmic microenvironment. Some of the salient differences in human ESC signaling pathways affecting their fate commitment are highlighted.
Introduction
Master Stem Cell Transcription Factors and the Regulation of Their Expression
The undifferentiated state of ESCs is maintained by the action of transcription factors, some of which are ESC specific and common to humans and the mouse. Three transcription factors the octamer binding protein 4 (Oct4) (Nichols et al., 1998; Schöler et al., 1989), the SRY-related HMG-box gene 2 (Sox2) (Yuan et al., 1995), and Nanog (Chambers et al., 2003; Mitsui et al., 2003) are key regulators in the self-renewal of ESCs. Their ectopic expression is sufficient to reprogram somatic cells to the state of pluripotency (Nakagawa et al., 2008; Takahashi and Yamanaka, 2006; Yu et al., 2007). The three master transcription factors locate at the heart of the regulatory network in ESCs. They bind to large numbers of both transcribed and inactive genes, some of which are responsible for maintaining ESCs by controlling self-renewal and pluripotency, and others perform key developmental functions that include differentiation into extraembryonic, endodermal, mesodermal, and ectodermal cell types. Many of the ESC maintenance genes are co-occupied by at least two of the three factors, including the Oct4, Nanog, and Sox2 genes themselves (Boyer et al., 2005; Ivanova et al., 2006; Loh et al., 2006; Zhou et al., 2007).
Oct4 (encoded by Pou5f1, also known as Oct3) is a POU domain-containing transcription factor that binds to an octamer sequence, ATGCAAAT. During mouse preimplantation development, zygotic Oct4 expression is activated at the four-cell stage and is later restricted to the pluripotent cells of the inner cell mass (ICM) and epiblast. In the postimplantation embryo Oct4 expression is downregulated upon epiblast differentiation and its expression is maintained only in the primordial germ cells (PGCs) (Yeom et al, 1996). The activity of Oct4 is essential for the identity of the pluripotent founder cell population in the mammalian embryo and null mutants of Oct4 embryos are not viable and fail to form a functional ICM (Nichols et al., 1998). The precise level of Oct4 is critical for sustaining ESC self-renewal, a twofold increase or decrease of Oct4 levels in ESCs leads to differentiation into primitive endoderm/mesoderm or trophectoderm, respectively (Niwa et al., 2000). Oct4-transfected keratinocytes express other core stem cell regulators, such as Sox2, Nanog, and Rex1 (Grinnell et al., 2007); and compared with RNAi knockdowns of Nanog, Sox2, Tbx3, Esrrb, and Tcl1, Oct4 knockdown cause ESCs show a very dramatic change in gene expression (Ivanova et al., 2006). Moreover, exogenous expression of only Oct4 is sufficient to generate pluripotent stem cells from adult mouse neural stem cells (Kim et al., 2009). These results indicated that Oct4 might be a chief selector for ESC fate decision.
Sox2 is a HMG domain-containing transcription factor essential for pluripotent cell development (Avilion et al., 2003). Sox2 has been identified as target gene of Oct4 (Grinnell et al., 2007), which has an expression pattern similar to that of Oct4 through mouse preimplantation development, as it is expressed in all blastomeres of the four-cell embryo and becomes restricted to the ICM and epiblast of the blastocyst (Avilion et al., 2003). Sox2 forms a complex with Oct4 to stimulate the induction of most pluripotency-associated genes, such as Nanog, Fgf4, Utf1, and Lefty1 (Nakatake et al., 2006; Nishimoto et al., 1999; Rodda et al., 2005; Yuan et al., 1995). The octamer elements within the enhancers of Oct4 target genes are in proximity to Sox2-binding sox elements. Two structures have been identified for a POU/HMG ternary complex bound to composite sox–oct elements: one of these is on an element separated by 3 bp (Remenyi et al., 2003), and the other is on an element separated by 0 bp (Williams et al., 2004). Both revealed that the POU and HMG domains mediate specific protein–protein and DNA–protein interactions. Sox2 and Oct4 can even interact in the absence of DNA, and that the HMG and POU domains are involved in this interaction (Ambrosetti et al., 2000).
The requirement of ESCs for Sox2 appears not to be based solely on the activity of the oct–sox enhancers. Sox2 is indispensable for maintaining ESC pluripotency because Sox2-null ESCs differentiate primarily into trophoectoderm-like cells. The essential function of Sox2 for ESCs is to maintain the requisite level of Oct4 by regulating multiple transcription factors that affect Oct4 expression (Masui et al., 2007).
Nanog is a homeodomain protein preferentially binds the TAAT(G/T)(G/T) motif in mouse (Ralf et al., 2008) and plays a critical role in regulating the cell fate of the pluripotent ICM during embryonic development, maintaining the pluripotent epiblast and preventing differentiation to primitive endoderm (Chambers et al., 2003). In the mouse, Nanog mRNA is first detected in the interior cells of the compacted morula, then confined to the ICM, and disappears in the trophectoderm in the blastocyst stage. In later blastocysts, Nanog expression is further restricted in the epiblast and excluded from the primitive endoderm. After implantation, Nanog expression is downregulated, but can be detected in germ cells of the genital ridges of E11.5 embryos. In vitro, Nanog mRNA is found in pluripotent ESCs, embryonic germ (EG) cells, and embryonic carcinoma (EC) cells, but not in adult tissues. Loss of Nanog is embryonic lethal and induces differentiation of ESCs toward an endodermal-like lineage, whereas forced expression retains the ESC phenotype in the absence of the otherwise essential leukemia-inhibitory factor (LIF ) (Chambers et al., 2003; Mitsui et al., 2003). In humans, Nanog is specifically expressed in the pluripotent cells of the ICM and epiblast, and is rapidly downregulated upon differentiation of ESCs (Hyslop et al., 2005). Inhibition of Nanog expression also causes human ESCs differentiation to extraembryonic cell lineages (Hyslop et al., 2005; Zaehres et al., 2005), whereas overexpression allows ESCs feeder-independent proliferation (Darr et al., 2006). Nanog, therefore, is considered a core element of the pluripotent transcriptional network and stabilizes the pluripotent state of ESC (Pan et al., 2007).
Expression regulation of master stem cell transcription factors
With regard to expression of the three master regulators, the precise regulating mechanism is still not yet clear. What we know is that auto- and reciprocal regulation among themselves exist in ESCs (Chew et al., 2005; Okumura-Nakanishi et al., 2005) (see Fig. 1). Both Oct4 and Sox2 bind directly to the composite sox–oct elements in both Oct4 and Sox2 genes in mouse and human ESCs, thus maintaining Oct4 and Sox2 expression via a positive and potentially self-reinforcing regulatory loop (Chew et al., 2005). The Oct4–Sox2 complex can also activate Nanog expression through binding to the promoter region (−322/+50) of Nanog (Rodda et al., 2005). Nanog, in turn, positively regulates the expression of Oct4, Sox2, and Nanog itself in mouse and human ESCs, by binding to their promoter regions, respectively (Boyer et al., 2005; Chickarmane et al., 2006; Loh et al., 2006). Besides auto- and reciprocal regulation, the expression of the three master factors also needs the regulation of other factors. It has been shown that, Myc, Nobox, GAbpα, SF1, RAR, and Lrh1 are involved in regulating Oct4 expression, whereas Sall4, Klfs, Esrrb, Tcf7, and esBaf are the common regulators of the three factors.
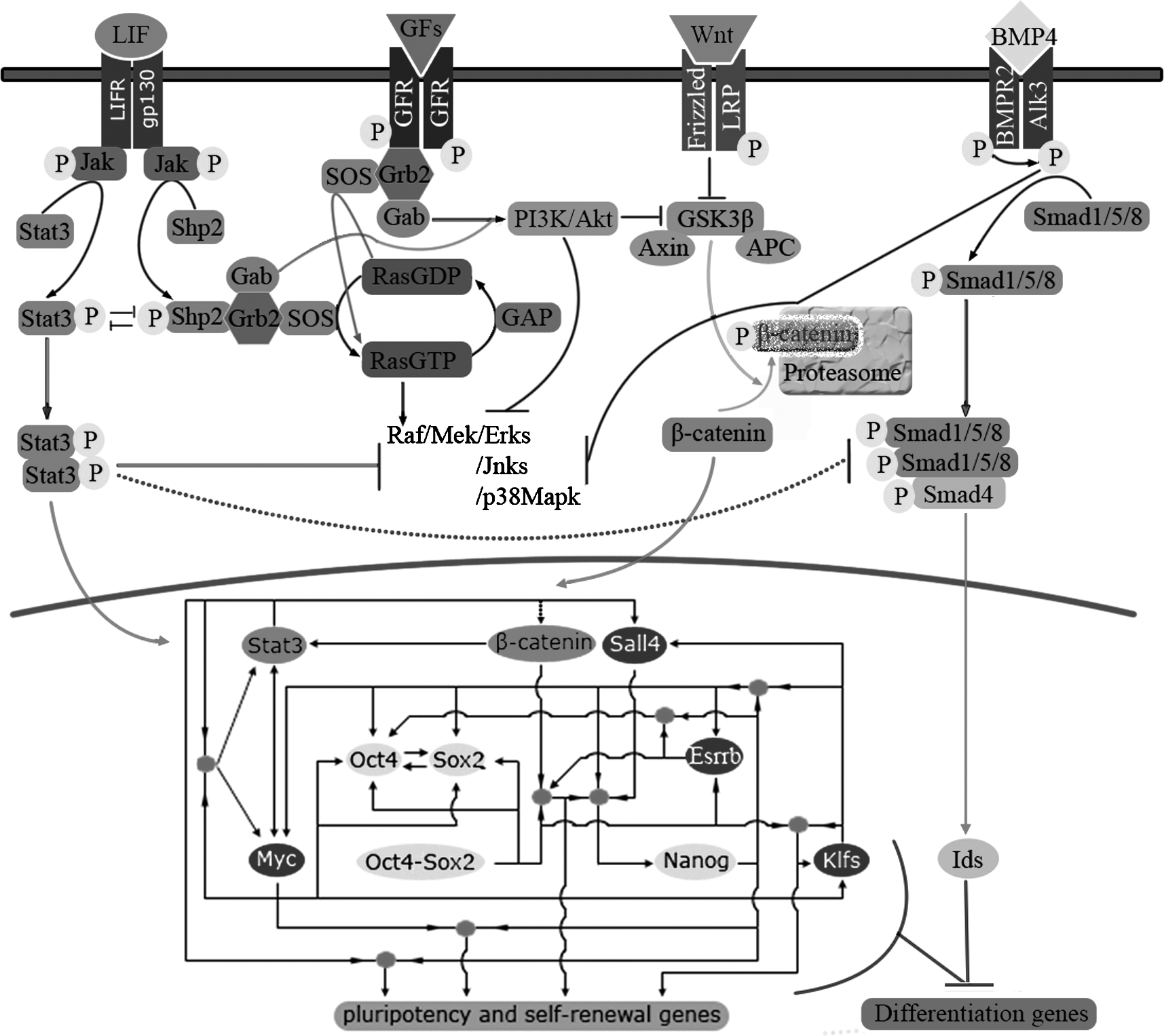
Diagram of signaling interaction and core transcription regulation network in mouse ES. LIF/Stat3 and β-catenin signaling act synergistically to promote self-renewal of mouse ESCs. LIF stimulates the increase of β-catenin through the PI3K/Akt pathway or targeting the Aes1 expression. Aes1 then acts by derepressing Tcf-mediated transactivation. Wnt/β-catenin-mediated Stat3 transcription would enhance Stat3 activation, enabling subthreshold levels of ligand to maintain self-renewal. The synergistic effect of BMP4 and LIF on mouse ES cell self-renewal is attributed to the ability of LIF to activate Jak–Stat pathway and to the ability of BMP4 to antagonise the stimulation of Erks by LIF, to inhibit p38 MAPK and upregulate Id genes. Oct4, Sox2, and Nanog locate at the heart of the regulatory network in ES cell. They bind to large numbers of pluripotency and self-renewal related genes, including themselves. Beta-catenin, Stat3, Myc, Klfs, Sall4, and Esrrb are the principle coregulators of the three master factors. Note: 1. Arrowed factors means be actively regulated by others; –| indicates inhibition. 2. Solid line symbol actural or direct effects; Dash line symbol putative or indirect effects. 3. Solid cycles means coregulating of two or more factors.
Elevated Myc activity is required for ESC maintenance. ChIP–chip (Chromatin-immunoprecipitatio–chip) studies revealed that Myc can bind to Oct4 and Sox2 genes and positively regulate their expression. As stated below, Myc is target of both Stat3 and β-catenin. It can also bind and positively regulate other active genes in ESCs, such as Mycn, Rest, Stat3, Mbd3, Jmjd3, Gdf3, Fbxo15, Klf5, Klf7, and Klf9, many of which were cobound by Nanog (Jiang et al., 2008; Kidder et al., 2008). The activation of Myc transcription can support the unusual cell cycle of ESC characterized by a very short G1 period and by suppression of the apoptosis and the senescence pathways. This may account for its function to maintain ESC pluripotency in the absence of LIF (Cartwright et al., 2005; De Felici et al., 2009). Myc binds three out of the four reprogramming factors (Oct4, Sox2, Mycn) and one other another gene, Klf5, which is functionally redundant to the reprogramming factor Klf4 (Jiang et al., 2008). It is noted that Myc is dispensable for inducing pluripotency, although the frequency is lower without it (Nakagawa et al., 2008). A genome-wide survey of Myc promoter occupancy in mouse ESCs leads to the suggestion that Myc may enhance reprogramming by augmenting transcription of pluripotency inducing factors through direct promoter binding (Kidder et al., 2008).
Newborn ovary homeobox (Nobox) is a germ cell-specific homeobox containing transcription factor, which is critical in the early oocyte differentiation. Nobox deficiency disrupts early folliculogenesis and the expression of oocyte-specific genes in the mouse (Choi et al., 2007). The Nobox binding element has been identified in the promoter regions (−426) of the mouse Oct4 gene. Nobox can bind to this site with high affinity and augment transcriptional activity of the luciferase reporter driven by mouse Oct4 promoters, suggesting that Nobox directly regulates the transcription of Oct4 (Choi and Rajkovic, 2006). Thus, it is likely that Nobox response to endogenous expression of Oct4 in oocytes during early folliculogenesis.
GA-repeat binding protein α (GAbpα) is a member of Ets (erythroblast transformation-specific) transcription factors. Ets factors share an evolutionarily conserved DNA binding domain of 85 amino acids that assumes a “winged-helix-turn-helix” configuration, which preferentially binds to purine-rich sequences that contain a GGAA/T core (Rosmarin et al., 2004). In mouse ESCs, as a downstream molecule of the Stat3 pathway, GAbpα regulates Oct4 expression by inhibiting three Oct4 repressors, Cdx2, Coup-tf1 (NR2F1), and GCNF (NR6A1) (Kinoshita et al., 2007). The caudal-related homeobox protein Cdx2 is a transcriptional regulator essential for trophoblast lineage, functioning as early as implantation. Gain of Cdx2 function in ESCs triggers trophoblast-like morphological differentiation, accompanied by loss of pluripotency-associated gene expression (Tolkunova et al., 2006). The nuclear receptor Coup-tf (chicken ovalbumin upstream promoter-transcription factors) proteins are implicated in the regulation of embryonic development and neuronal cell differentiation. Coup-tf1 is an important signaling molecule during vitamin A-mediated very early stage of embryonic development (Zhuang and Gudas, 2008). The function of GCNF (germ cell nuclear factor) is to repress Oct4 expression in somatic cells after gastrulation by binding as a homodimer to a DR0 element (a direct repeat element with zero nucleotides between the reiterated sequence AGGTCA) located in the Oct4 proximal promoter (Hummelke and Cooney, 2001).
Unlike GCNF, another group of nuclear receptors, such as steroidogenic factor-1 (SF1), retinoid acid nuclear receptors (RARs), and liver receptor homologue 1 (Lrh1; NR5A2), support the restricted expression of Oct4 in germ cells. Studies in murine P19 EC cells revealed that interaction of the SF1(s) with the RARs involved in Oct4 expression regulation. There are two SF1-binding sites identified in the Oct4 promoter, the more crucial proximal site located within the RA-responsive element. SF1 and RAR form a complex on the RA-responsive element that synergistically activates the Oct4 promoter (Barnea and Bergman, 2000). SF1 knockout mice lack adrenal glands and gonads, and die due to adrenal insufficiency within the first week of birth, indicating that the regulation of SF1 on Oct4 expression is essential only during late organogenesis (Luo et al., 1994). During early embryonic development and in ESCs, the expression of Oct4 involved the function of Lrh1. Lrh1 is closely related to SF1, particularly in the DNA binding domain, and has the same DNA response element as SF1. Besides functioning in endoderm development as known initially, Lrh1 is also essential for development of early-stage embryos, Lrh-1−/− embryos died at E6.5 to 7.5. The expression pattern of Lrh1 in ESCs and early embryos overlaps with Oct4; Lrh1 can bind directly to response elements in the proximal promoters and proximal enhancer of the Oct4 gene and activate Oct4 reporter gene expression (Gu et al., 2005), suggested that Lrh1 plays an important role in the regulation of Oct4 gene expression in ESCs and early-stage embryos.
Sall4, Klfs, Esrrb, Tcf7, and esBaf can activate expression of each of the three master factors. Sall4 is a zinc-finger transcriptional regulator, belongs to the spalt family. As a target of Nobox (Choi et al., 2007) and Klfs (Jiang et al., 2008), Sall4 has a critical role in the maintenance of ESC pluripotency. By binding directly to enhancers of Oct4 and Nanog it participates in activating the expression of the two genes (Jiang et al., 2008; Zhang et al., 2006). Krüppel-like factor 2 (Klf2), Klf4 and Klf5 are concurrently required for the self-renewal of mouse ESCs. Klfs and Nanog co-occupy many common binding loci; a large fraction of those genes are required for self-renewal and maintenance of pluripotency of ESCs, including Nanog, Esrrb, Rif1, Pou5f1, Sox2, Tcf3, Mycn, and Foxd3 (Jiang et al., 2008). Klf4 has been identified as a mediating factor that binds to the proximal element and cooperates with Oct4–Sox2 on the distal enhancer in activating a set of Oct4–Sox2 target genes promoter, such as Lefty1 and Klf4 itself (Nakatake et al., 2006). In human ESCs, it has been shown recently that Klf4 and another important transcription factor Pbx1 directly bind to Nanog proximal promoter, and cooperated with Oct4–Sox2, synergistically transactivated the Nanog promoter activity. The location of Klf4 binding motifs is very different from those in mouse Nanog promoter, which is located at the distal enhancer. Pbx1, a homeobox gene, was originally discovered by their involvement in chromosomal translocation in acute pre-B cell leukemia. Pbx1 belongs to the PBC subclass of TALE homeodomain protein, which is important for vertebrate development (Chan et al., 2009).
As a direct target of the master regulators, Estrogen-related receptor beta (Esrrb), a member of the nuclear receptor family, has been showed participate in the regulation of them (Zhou et al., 2007). The binding sites of Esrrb are in the proximal 5′-untranslated region of the mouse Oct4 gene, one of which is in close proximity to a Nanog binding site. Esrrb and Nanog interact through their DNA binding domains; this interaction reciprocally modulates their transcriptional activities and activates Oct4 expression (Zhang et al., 2008). Esrrb has also been demonstrated be recruited near the oct–sox element in the Nanog proximal promoter, interact with Oct4, and positively regulates Nanog expression (van den Berg et al., 2008). Tcf7 is a transcriptional regulator downstream of Wnt signaling. It not only is a direct target of the master regulators but also participates in the regulation of them (Zhou et al., 2007).
Mammalian switching/sucrose nonfermenting [Swi/Snf, also called Baf (Brg/Brahma-associated factors)] ATP-dependent chromatin remodeling complexes are essential for formation of the totipotent and pluripotent cells of the early embryo. Baf complexes in ESCs (esBaf ) include Brg (Brahma-related gene), Baf155 and Baf250A. Ho et al. (2009a, 2009b) reported recently that, Brg and Baf155 extensively bind to each of the ESC regulator genes, including Oct4, Sox2, Nanog, Dppa2, Dppa4, Sall4, and Myc; whereas Brg binds significantly to Klf4 and Rif1. esBaf complexes functionally interact with Oct4, Sox2, and Nanog at their common target genes. Possibly, esBaf is recruited by Oct4–Sox2 heterodimers, and engages chromatin through its multiple histone recognition and DNA-binding domains to remodel its target sites to enable binding of Oct4–Sox2. The requirement of Brg in mouse ESC maintenance might also be necessary because of its colocalization with Smad1 and Stat3. esBaf is likely required for the initial establishment of the pluripotent circuitry, given that Brg is a maternal effect gene found to be essential for zygotic gene activation (Bultman et al., 2006) and somatic nuclear reprogramming (Hansis et al., 2004). Once established, the core circuitry might become less dependent on the activity of esBaf (Ho et al., 2009a, 2009b).
The stability and transcriptional activity of master stem cell factors could also be regulated at the posttranslation level. It has been found that cAMP-responsive phosphorylation at site serine 229 of Oct4 likely causes steric and electrostatic disruption of Oct4 homodimer–DNA configuration, enhancing the transactivation activity to PORE (Palindromic oct factor recognition element)-containing genes. The enhancement is at least partially regulated through the mitogen-activated protein kinases (MAPK) pathway, which is mediated by a synergistic effect between protein kinase A (PKA) and EPAC (an intracellular cAMP receptor) (Saxe et al., 2009).
Signaling in Stem Cell Maintenance
ESCs were initially established and maintained by coculture with murine embryonic fibroblast (MEF ) feeder cells (Evans and Kaufman, 1981). Subsequent studies identified that fibroblasts secrete multiple factors, including LIF, fibroblast growth factors (FGFs), transforming growth factor β (TGFβ), Activin, Wnts, insulin-like growth factor (IGF ), and antagonists of BMP signaling (Beattie et al., 2005; Prowse et al., 2007; Williams et al., 1988). ESCs are normally derived and maintained in media containing these factors in combination. In a neutralized environment, ESCs can be efficiently derived and maintained without a requirement for growth factors or cytokines, suggesting that ESCs have an innate program for self-replication. The extrinsic signaling probably do not instruct self-renewal but acts in unrefined culture conditions to shield the pluripotent state by the elimination of differentiation-inducing (e.g., MAPK) signaling (Ying et al., 2008).
LIF/Stat3 signaling
LIF is a pleiotropic cytokine belonging to the interleukin 6 (IL6) cytokine family. It is secreted by various cell types and mediates opposite effects (either proliferative or differentiative, apoptotic or antiapoptotic) depending on the cell lineage and stage of differentiation (De Felici et al., 2009). LIF has been shown to maintain mouse ESCs in vitro and to enable their derivation (Smith and Hooper, 1987), an effect attributed to signal transducer and activator of transcription 3 (Stat3) activation (Niwa et al., 1998; Ying et al., 2003a). LIF signaling in mouse ESC is initiated through dimerization of the LIF receptors (LIFRs) and gp130 following their engagement by the cytokine. This signals the activation of Janus kinases (Jaks), which phosphorylate LIFR and gp130 on the tyrosine residues. Stat3 is then recruited to the receptor complex where it is phosphorylated by Jak at tyrosine 705, resulting in its subsequent dimerization, nuclear translocation, and target gene activation (Hyslop et al., 2005) (see Fig. 1).
It is likely that LIF/Stat3 signaling supports mouse ESC self-renewal and pluripotency via several ways. First, LIF signaling inhibits differentiation signaling pathways. Phosphorylated Stat3 counteracts the programs of differentiation by directly or indirectly inhibiting the receptor-mediated Smad (R-Smad) complex and/or growth factors signaling downstream of extracellular signal-regulated kinase (Erk) (see below). Transcription of LIF-dependent genes activated in differentiation pathways can be inhibited by induction of suppressor of cytokine signaling 3 (Socs3) by LIF itself (De Felici et al., 2009). Second, Stat3 sustains the threshold expression of master stem cell factors requisite for maintaining self-renewal and pluripotency. Mouse ESCs represent a ground state in which they have an innate ability for self-renewal if without signaling factors such as FGF4 (Gotoh, 2009; Ying et al., 2008). But in the absence of LIF stimulation, the endogenous expression of Oct4, Nanog, and inhibitor of differentiation 1–3 (Id 1–3) is insufficient to sustain an ESC state (Ying et al., 2003). It has been shown that Nanog expression is upregulated in mouse ESCs by the binding of Stat3 and T (Brachyury) to an enhancer element in the Nanog gene (Suzuki et al., 2006), and that the levels of Stat3 activation and Oct4 expression correlate in single cell in the presence and absence of LIF (Davey et al., 2006). Third, cooperating with Nanog or Oct4, Stat3 might directly/indirectly transactivate pluripotency genes or silence differentiation-associated genes in ESCs. As shown by ChIP–chip studies utilizing promoter-tiling arrays, Stat3 can bind to many active genes in ESC, including Myc, Sall4, Stat3, Rest, Mbd3, Jmjd3, Tdgf1, and Fbxo15. In the absence of LIF stimulation, some ESC-enriched genes such as Rex1, Utf1, and Bmp4 downregulated (Kidder et al., 2008). Many Stat3 target active genes co-occupy with Nanog and contain histone H3 acetylation modifications (Kidder et al., 2008). They are also bound by Myc such as Mycn, Rest, Stat3, Mbd3, and Jmjd3 (Cartwright et al., 2005; Kidder et al., 2008). Stat3 could also cooperate with Oct4 to maintain silencing of differentiation-associated genes through upregulation of Eed (embryonic ectoderm development) (Ura et al., 2008), and to maintain the self-renewal of ESCs through control the expression of Dax1 (Sun et al., 2008). Eed is a major component of polycomb repressive complex (PRC)-2, which is involved in methylation of histone H3 at Lysine 27 (Ura et al., 2008). Dax1 belongs to the nuclear hormone receptor superfamily and is expressed in undifferentiated ESCs. Dax1 binds with Nanog and shares multiple targets with Oct4 and Nanog, playing an important role in ESC self-renewal (Sun et al., 2008). The Aes1 gene is also directly transactivated by Stat3 in ESCs. Aes1 belongs to the Tle/grg groucho-like family, but lacks the C-terminal region of Tle/grg proteins. Aes1 acts by de-repressing, rather than by inhibiting like the Tle/grg proteins, Tcf-mediated transactivation, and therefore increases the canonical Wnt signaling in ESCs (Sekkaï et al., 2005). The activation of the nonreceptor tyrosine kinases belonging to the Src family (i.e., Hck), are also important for ESC self-renewal and can be downstream of LIF signaling (Ernst et al., 1994).
Unlike in mouse ESCs, LIF is not essential for primate and rabbit ESC self-renewal (Honda et al., 2009; Humphrey et al., 2004). In human ESCs, LIF receptors are expressed, and LIF can stimulate activation of Stat3 under experiment condition, but this pathway is not activated in the undifferentiated state, suggesting that the maintenance of human ESCs is Stat3 independent (Humphrey et al., 2004; Sumi et al., 2004). LIF is also dispensable for maintaining undifferentiated status in rabbit ESCs. Notably, Stat3 is activated LIF-independently in rabbit ESC. But when Stat3 phosphorylation is prevent by Jak-inhibitor treatment, the Oct4 protein level does not decrease (Honda et al., 2009), indicating that the levels of Stat3 activation does not correlate with Oct4 expression. The function of Stat3 in rabbit ESCs needs to be elucidated. LIF signaling might be important during the early phase of establishing rabbit ESCs. As LIF receptor is expressed in rabbit ESCs establishing a rabbit ESC-like line could be improved in the presence of LIF. The withdrawal of LIF during early passages induces differentiation (Catunda et al., 2008).
Beta-catenin signaling
Beta-catenin is the central player in the canonical Wnt signaling pathway. The stability of β-catenin is regulated by the destruction complex, consisting of the tumor suppressors adenomatous polyposis coli (APC), axin, casein kinase 1 (CK1), and glycogen synthase kinase-3β (GSK3β). When Wnt receptors are not engaged, APC and axin bind newly synthesized β-catenin. CK1 and GSK3 then sequentially phosphorylate a set of conserved Ser and Thr residues in the amino terminus of β-catenin and the resulting phosphorylated footprint recruits E3 ubiquitin ligase, which targets β-catenin for proteasomal degradation. Wnt/β-catenin signaling is initiated when Wnt ligands engage their cognate receptor complex, consisting of Frizzled and Lrp5/6. Receptor occupancy inhibits GSK3β activity; as a consequence, β-catenin accumulates and travels into the nucleus where it engages the N-terminus of DNA-binding proteins of the Tcf (T cell factor)/Lef (lymphoid enhancer family) family. In the absence of a Wnt signal Tcf/Lef factors associate with histone deacetylase 1 (Hdac1), bind to Wnt response elements (WREs), TYYCTTTGATSTT, to serve as transcriptional repressors. Beta-catenin can replace Hdac1 to form a complex with Tcf/Lef to elicit sequence-specific transcriptional activation of the target genes (Atcha et al., 2007; Reya and Clevers, 2005). The accumulation of β-catenin may also be a downstream event of growth factors or LIF signaling (Paling et al., 2004). In these signaling transduction pathway, phosphoinositide 3-kinase (PI3K)/Akt mediates GSK3β inhibition, thus resulting in β-catenin stability and nuclear translocation (Fig. 1).
Wnt/β-catenin signaling has been demonstrated to maintain pluripotency in mouse and primate stem cells under MEF feeder cells or MEF-conditioned media (CM) (Dravid et al., 2005; Sato et al., 2004). The precise mode of action of β-catenin signaling in stem cells is not yet clear. From the information available, it is known that β-catenin functions in ESCs at least through three ways. First, β-catenin acts synergistically with Stat3 in mouse ESCs (see below). Second, β-catenin directly or indirectly regulates the expression of master stem cell transcription factors in ESCs. Oct4 expression in ESCs has been shown to be maintained by Wnt3A CM (Sato et al., 2004). The findings that Tcf3 binds to a promoter regulatory region of the Nanog gene and represses its transcription in ESCs through a Groucho interaction domain-dependent process demonstrates a regulatory role of β-catenin on Nanog expression (Pereira et al., 2006). The β-catenin-dependent upregulation of Nanog is indispensable to the Oct4 binding element on the Nanog promoter (Anton et al., 2007; Yukinari et al., 2007). Myc is also a direct target molecule of β-catenin, which is upregulated by β-catenin (Cartwright et al., 2005). Third, Wnt canonical signaling stimulates stem cell survival or proliferation, which is required for either self-renewal or differentiation. In the presence of bFGF and supportive feeder cells, Wnt stimulates the self-renewing proliferation of the undifferentiated human ESC population. In the absence of the feeder cells or CM however, excessive Wnt accelerates ESC proliferation as well as differentiation (Dravid et al., 2005). Wnt/β-catenin signaling is required for neural differentiation of ESCs (Otero et al., 2004). The divergent responses to activation of Wnt/β-catenin signaling in ESCs may attribute to the interaction molecules of β-catenin. It is reported that β-catenin/CBP [cAMP response element binding (CREB)-binding protein]-mediated transcription is critical for cell proliferation without differentiation, whereas a switch to β-catenin/p300 (a homologue of CBP)-mediated transcription is critical to initiate a differentiative program with a more limited proliferative capacity (Miyabayashi et al., 2007).
TGFβ factors signaling
LIF alone is insufficient to maintain mouse ESC self-renewal in serum-free conditions, whereas a combination of bone morphogenetic protein 4 (BMP4) and LIF has the ability (Ying et al., 2003a). In humans, LIF is not essential for ESC self-renewal (Humphrey et al., 2004); several other factors have been identified such as basic FGF (or FGF2) (Levenstein et al., 2006), TGFβ, Activin A/Nodal (Beattie et al., 2005; Vallier et al., 2005), Noggin (Xu et al., 2005), and activators of PI3K signaling such as insulin/IGF (Bendall et al., 2007). Media incorporating different combinations of these factors have been reported to sustain human ESCs in the absence of fibroblasts feeder layers (Amit et al., 2004; Vallier et al., 2005; Xu et al., 2005).
BMPs are members of the TGFβ superfamily. BMP proteins bind to heteromeric receptor complexes that contain type I and type II serine/threonine protein-kinase receptor subunits. In mouse ESCs, upon BMP4 binding to type I receptor (Alk3), formation of a heteromeric complex with type II receptor is induced. Consequently, this latter phosphorylates Alk3 in its kinase domain that, in turn, phosphorylates R-Smad1, 5, or 8. Activated R-Smads form a complex with the common mediator Smad4, and translocated into the nucleus, can bind directly or through transcriptional partners to specific sequences in the promoters of BMP target genes (De Felici et al., 2009) (Fig. 1). The major contribution of BMP4 to mouse ESC self-renewal seems the induction of the expression of the negative helix-loop-helix factors, Ids. Forced expression of Id genes in mouse ESCs is sufficient for maintaining self-renewal in the absence of BMP4 (Ying et al., 2003a). It is likely that such upregulation results from a combined Smad activation and inhibition of MAPKs (see below).
In contrast to mouse ESCs, human ESCs can induce trophoblast differentiation by BMP4 (Xu et al., 2005). The effect may at least partly owing to Smad 1/5/8 activation moderately represses Sox2 (Greber et al., 2008). Activin A, another TGFβ family member, is necessary and sufficient for the maintenance of self-renewal and pluripotency of human ESCs. It can induce the expression of Oct4, Nanog, Nodal, Wnt3, bFGF, and FGF8, and suppresses the BMP signal, support long-term growth of human ESCs on Matrigel-coated flasks without either feeder cells or conditioned medium (Xiao et al., 2006). Recently, TGFβ/Activin-responsive Smads have been demonstrated to directly bind to the Nanog promoter, and enhance Nanog promoter activity, and thus play its essential role in sustaining human ESC self-renewal (Xu et al., 2008). As observed in human ESCs, Activin/Nodal signaling is essential for supporting rabbit ESC self-renewal via activation of Smad2/3, resulting in upregulation of Oct4 and Nanog transcription (Honda et al., 2009). TGFβ factors play their role in collaboration with bFGF in human or rabbit ESCs (Greber et al., 2007; Honda et al., 2009; Xu et al., 2005).
FGF signaling
FGF signaling is dispensable for mouse ESC self-renewal, and it is thought to be normally inactivated in such cells. Even more, it appears that the FGF-dependent extracellular signal-regulated Erk1/2 and perhaps PI3K pathways are crucial for mouse ESCs to differentiate into both neural and nonneural lineages (Chen et al., 2000), and that bFGF stimulate proliferation of differentiating mouse ESCs via phospholipase gamma-protein kinase C (PLCγ–PKC) pathway (Dvorák et al., 1998). It is to be mentioned here that in contrast to mouse ESCs, FGF signaling appears to be central for human and rabbit ESC self-renewal (Honda et al., 2009; Vallier et al., 2005). Consistent with this, human iPS cells could be expanded in the presence of FGF but not in the presence of LIF, whereas mouse iPS cells require LIF (Gotoh, 2009).
Basic FGF may promote human ESC self-renewal indirectly by stimulating paracrine and autocrine effects. It has been reported that in vitro culture of human ESCs contains two cell types—pluripotent ESCs and fibroblast-like cells—that may be spontaneously derived from the ESCs. The fibroblast-like cells have FGF receptors and respond to bFGF, leading to the production of IGF. IGF then stimulates human ESCs to maintain their self-renewal capability in a paracrine manner. It is therefore proposed that IGF and bFGF cooperatively function to establish the regulatory stem cell niche (Bendall et al., 2007). Another bFGF paracrine effect is to stimulate the secretion of beneficial factors (i.e., TGFβ1) in MEFs, and suppress the release of differentiation-inducing activity such as BMP4 (Greber et al., 2007). bFGF may also modulate human ESC expressing TGFβ ligands, which in turn, act on human ESCs in a concerted and autocrine manner (Greber et al., 2007). Human ESCs express bFGF and its receptors (FGFRs). But the presence or absence of exogenous bFGF does not significantly affect the proliferation of the cells. However, the inhibition of FGFR suppresses activation of downstream protein kinases and causes rapid cell differentiation (Dvorak et al., 2005). bFGF knockdown induced human ESC differentiation, and expression of endogenous bFGF decreased during human ESC differentiation (Eiselleova et al., 2009). These results suggest that an involvement of autocrine bFGF signals in the maintenance of proliferating human ESCs in the undifferentiated state (Dvorak et al., 2005).
Another mechanism of bFGF promoting self-renewal in human ESCs is to antagonize the BMP pathway and to stimulate Mek (mitogen activated, Erk activating kinase)/Erk signaling. In contrast to mouse ESCs, high basal Mek/Erk activity is required for maintaining human ESCs in an undifferentiated state. In human ESCs, exogenous and endogenous bFGF synergistically activate Mek/Erk and PI3K/Akt signaling. Mek/Erk signaling cooperating with PI3K/Akt signaling stimulates the expression of stem cell genes while suppressing cell death and apoptosis genes (Eiselleova et al., 2009; Kang et al., 2005; Li et al., 2007).
Synergistic Effects in ESC Ccells
Wnt/β-catenin and the LIF/Stat3 synergize to maintain pluripotency and self-renewal of mouse ESCs
In the absence of LIF, β-catenin is not sufficient to keep pluripotency of mouse ESCs (Dravid et al., 2005; Yukinari et al., 2007), although low levels of LIF that are otherwise insufficient to maintain self-renewal can do so in the presence of Wnt (Davey et al., 2007), suggesting that Wnt/β-catenin and the LIF/Stat3 act synergistically to promote self-renewal (Ogawa et al., 2006) (Fig. 1).
The pathway from LIF stimulation to the inactivation of GSK3β through the PI3K/Akt pathway has been described in mouse ESCs (Paling et al., 2004). LIF stimulation can induce phosphorylation of the effector protein Shp2. Shp2 is a cytoplasmic tyrosine phosphatase with two Src-homology 2 (SH2) domains at the NH2-terminus that modulates signal strength downstream of cytokine/growth factor receptors. Shp2 binds through SH2 domains to Y757 of the intracellular domain of gp130 and Y969 of LIFR. When phosphorylated by Jak, Shp2 can bind Grb2-associated binder (Gab), activating the PI3K kinase pathway, subsequent GSK3β inhibition results in β-catenin stability and nuclear translocation (Paling et al., 2004) (Fig. 1). Stat3 may also directly target the Aes1 expression in ESCs; Aes1 then acts by de-repressing Tcf-mediated transactivation and increasing the canonical Wnt signaling (Sekkaï et al., 2005).
However, Wnt/β-catenin can upregulate Stat3. In the presence of Wnt or constitutively active β-catenin, a two- to theefold increase in Stat3 expression and protein has been observed, but the resultant Stat3 proteins were incapable of inhibiting ESC differentiation unless serum or LIF was added to the culture. The Wnt/β-catenin pathway may act through convergence on the LIF/Jak–Stat pathway at the level of Stat3; the former increases Stat3 mRNA, whereas the latter promotes the phosphorylation of Stat3 proteins (Hao et al., 2006). Interestingly, low levels of LIF that are otherwise insufficient to maintain self-renewal can do so in the presence of Wnt or constitutively active β-catenin. A positive feedback loop in Jak–Stat pathway provides insight into the role of Wnt signaling in self-renewal. In ESC, Stat3 activation controls the expression of Stat3, gp130, and LIFR. Therefore, in the presence of Wnt, β-catenin-mediated Stat3 transcription would enhance Stat3 activation, enabling subthreshold levels of ligand to maintain self-renewal (Davey et al., 2007).
BMP4 and LIF function synergistically in mouse ESC
The maintenance of mouse ESCs depend on the balance between LIF signaling and BMP4 signaling (Qi et al., 2004; Ying et al., 2003a). In one way, BMP4 signaling inhibits LIF-dependent differentiation. LIF phosphorylated Shp2 can bind the adaptor protein growth factor receptor-bound protein 2 (Grb2) and activate the Ras/Raf/Mek/MAPK signaling pathway (Burdon et al., 1999). The MAPK family comprises four groups of proteins: Erk 1 and 2, Erk5, c-Jun aminoterminal kinases (Jnks) 1, 2, and 3, and p38MAPKs (De Felici et al., 2009). LIF-dependent MAPK activation appears to play a dual opposite role in ESCs by contributing to the undifferentiated status through stimulation of Stat3 phosphorylation (Ernst et al., 1994) and by favoring their differentiation (Chen et al., 2000). In ESC culture, the differentiation MAPK pathways are normally inhibited either by the BMP4 signaling or by PI3K (Paling et al., 2004) (Fig. 1). For instance, it has been reported that activated BMP4 receptor can cause the inactivation (dephosphorylation) of Erks and MAPK, likely by MAPK phosphatase HePTP (hematopoietic protein tyrosine phosphatase) (Qi et al., 2004). Inhibition of PI3K has been found to augment LIF-induced phosphorylation of Erks, leading to a reduction in the ability of LIF to maintain self-renewal (Paling et al., 2004).
Alternatively, LIF signaling blocks BMP4 induced differentiation events. In the absence of LIF, BMP4 alone promotes differentiation of mouse ESCs into nonneural fates (Ying et al., 2003b). Phosphorylated Stat3 counteracts the programs of mesodermal/endodermal differentiation by directly or indirectly inhibiting the R-Smad-complex (De Felici et al., 2009). In mouse ESCs, the roles of LIF is also including to eliminate differentiation-inducing signaling from Erk stimulated by serum or feeder cell-derived growth factors such as FGF, which can promote commitment of mouse ESCs to multiple lineages (Chen et al., 2000; Ying et al., 2008). In brief, the synergistic effect of BMP4 and LIF on mouse ESC self-renewal is likely attributed to the ability of LIF to activate Jak–Stat pathway and to the ability of BMP4 to antagonize the stimulation of Erks by LIF, to inhibit p38 MAPK and upregulate Id genes (De Felici et al., 2009) (Fig. 1).
Master ESC transcription factors co-occupy target genes with other stemness regulators
It can be concluded from different parts of above statements that Oct4–Sox2 interacts with β-catenin, Stat3, and Klfs, and that Nanog interacts with Klfs, Stat3, and Myc, to play their role in ESCs (see Fig. 1). These features showed that stemness regulators form a highly interconnected transcriptional regulatory network: one example is the extensive cotargeting of the two regulatory loci of Nanog by transcription factors important to maintain ESCs. By coaction with β-catenin, Oct4–Sox2 recruiting Esrrb occupy the Nanog proximal promoter, whereas Sall4, Nanog itself, Klf2, Klf4, and Klf5 directly bind to the Nanog enhancer (Jiang et al., 2008; van den Berg et al., 2008; Yukinari et al., 2007) (Fig. 1).
Dose-Dependent Controls in ESC Cells
Stem cells must decide between contrasting cell fates: proliferation versus quiescence, self-renewal versus differentiation, survival versus apoptosis. These fates are generally considered to be all-or-nothing (yes/no) responses. These switch-like mechanisms may arise at the level of cell signaling pathways or in downstream cell-fate machinery (transcription factor networks), each with unique implications on the nature of stem cell fate control.
A computational model of the dynamics of Nanog, Oct4, and Sox2 core network has revealed that it functions as a bistable switch, which is in the “on” state. It corresponds to these transcription factors being expressed and the downstream differentiation target genes being shut off (Chickarmane et al., 2006). In the case of Myc, the finely tuned expression level is very important for ESC maintenance, because sustained activation of Myc in human ESCs induces apoptosis and differentiation with reduced expression of the pluripotent markers Oct4 and Nanog (Sumi et al., 2007).
A specific threshold of β-catenin signaling might be required for pluripotency maintenance, because somatic cell reprogramming is triggered by a dose-dependent accumulation of active β-catenin when fused with ESC (Lluis et al., 2008). A switch-like (on/off ) signaling in response to LIF is also found in mouse ESCs. ESCs express functional levels of gp130 activating ligands that act to temporarily support Stat3 activation and self-renewal in the absence of exogenous LIF (Davey et al., 2006). However, the autocrine signaling and low concentrations of exogenous LIF cause ESCs to adopt the weakly responsive (“off”) state of differentiated cells. Endogenous Stat3 activity decreases with time in the absence of LIF, and ESCs gradually lose ligand responsiveness reversibly preceding the loss of the ESC transcription factors Oct4 and Nanog. ESC responsiveness to gp130 ligands is in a ligand concentration-dependent manner. At sufficient concentrations of exogenous LIF, a positive feedback loop, whereby Stat3 activation controls the expression of Stat3, gp130, and LIFR, maintains robust pathway responsiveness (“on”) (Davey et al., 2007).
In conclusion, the undifferentiated state maintenance of ESCs is regulated mainly by the action of three key transcription factors, Oct4, Sox2, and Nanog. Beta-catenin, Stat3, Myc, and Klfs are the principle coregulators of the master factors. The threshold expressions of Oct4, Sox2, and Nanog for pluripotency maintenance and self-renewal depend on the regulation of synergistic effects among β-catenin, Stat3, and Smads signaling pathway. Beta-catenin, Stat3, or Smads signalings indeed correlate to the expression of the three master factors, but the correlated regulation may be indispensible for specific condition of ESC cytoplasmic microenvironment. Therefore, if we apply these signaling mechanisms when establishing an in vitro somatic cell reprogramming approach without exogenous gene insertion, the establishment of β-catenin and Stat3 synergistic effects in somatic cells, and the creation of specific intracellular environments for β-catenin and Stat3 to initiate the transcription of key pluripotent factors, will prove to be difficult problems scientists must overcome first. Interestingly, some reports have revealed that the cell culture confluency effect the activity of β-catenin and Stat3 signaling (Steinman et al., 2003; Su et al., 2007; Vultur et al., 2004). Confluence-dependent signaling suggests a link between cell confluence and stem cell factor expression. Hepatocyte growth factor (HGF ) could stimulate β-catenin activation and nucleus translocation as its receptor remains active even after confluence, thus preventing cell apoptosis (Ishibe et al., 2006). It may be worthwhile testing the possibility of driving the reprogramming process by HGF and confluence-dependent signaling.
Footnotes
Acknowledgments
This work was supported by Shandong province reward fund to young scientist, and Weihai marine institute fund.
Author Disclosure Statement
The author declares that no conflicting financial interests exist.