
Abstract
Abstract
Fusion of terminally differentiated somatic cells with pluripotent embryonic stem cells has been proposed as model for reprogramming the somatic cell genome, and may contribute to our understanding of the underlying mechanisms of this epigenetic process. We established an interspecies cell fusion model using murine embryonic stem cells (ESCs) and porcine fibroblasts. These inter-species fusion experiments yielded much lower conversion efficiency rates than murine intraspecies fusion. Nevertheless, two double-resistant mouse–pig hybrid clones could be generated. Reactivation of the porcine OCT4 gene, an essential pluripotent stem cell marker, and demethylation of the porcine OCT4 promoter in hybrid clone 1, suggested successful reprogramming of porcine chromosomes. A rapid loss of porcine chromosomes was observed during the selection phase. Spectral karyotyping (SKY) analysis showed that fusion-hybrid clone 1 carried a tetraploid mouse chromosome complement with only few pig chromosomes and/or chromosomal fragments. Hybrid clone 2 had a diploid set of murine chromosomes complements and also contained an interspecies chromosome fusion product. Interspecies cell fusion results in hybrid cells that retained the complement of mouse chromosomes and preferentially lose porcine chromosomes during colony expansion. Neither species-specific chromosomal segregation nor reprogrammed diploid porcine cells were observed. These findings indicate that generation of reprogrammed pluripotent diploid cells by cell fusion may require additional supporting provisions.
Introduction
One way to obtain large numbers of patient-specific cells for use in cell therapy is to reprogram differentiated somatic cells from the patient into cells capable of expansion in culture. Resetting the epigenetic control of differentiated cell gene expression is a complex process, but protocols are being developed to achieve this goal. One protocol is based on somatic cell nuclear transfer (SCNT), where the nucleus of an adult somatic cell is reprogrammed by transfer into an enucleated oocyte (Bui et al., 2008; Wakayama et al., 2001). This results in preimplantation embryo “clones” from which stem cells can be cultured. Patient-specific embryonic stem cells derived from SCNT embryos could be used for the establishment of differentiated therapeutic cell lines (French et al., 2008). Unfortunately, therapeutic cloning for human medicine is limited by the requirement of human oocytes. The use in human embryos as a source of stem cells is forbidden for ethical reasons in many countries (Kfoury et al., 2007).
In a recent breakthrough, direct nuclear reprogramming of somatic cells was achieved by transfection with Oct4, Sox2, c-Myc, and Klf4, carried in retroviral vectors. Fully differentiated fibroblasts from both the mouse and humans were reprogrammed to a pluripotent stem cell state. These induced pluripotent stem cells (iPS) are indistinguishable from the embryonic stem cells (ESCs) derived from embryos with regard to morphology and differentiation potential (Takahashi and Yamanaka, 2006; Takahashi et al., 2007; Wernig et al., 2007), and could even produce germ cells (Aoi et al., 2008; Takahashi et al., 2007). This protocol is an attractive alternative for reprogramming of somatic cells as it avoids the ethical constraints associated with the human ESCs (Takahashi et al., 2007), but for cell therapy applications, there is some concern about the viral vectors and the use of oncogenes for reprogramming.
An alternative approach to reprogramming is to use the factors present in pluripotent cells to reprogram a somatic nucleus by cell fusion (Do and Schöler, 2004; Silva et al., 2006). Several groups have successfully demonstrated that this results in hybrid cells in which pluripotency of the somatic nucleus is reactivated (Cowan et al., 2005; Do et al., 2007; Yu et al., 2006). The most reliable marker to confirm reprogramming is the pluripotency-related gene OCT4, which is characteristically expressed in stem cells but is inactive in somatic cells. In contrast to SCNT, where the reprogramming factors are provided by the enucleated ooplasm, reprogramming by fusion with embryonic stem cells critically depends on the presence of the stem cell nucleus (Do and Schöler, 2004). Several pluripotent cell types, including ESCs (Ambrosi et al., 2007; Cowan et al., 2005; Do and Schöler, 2004; Matveeva et al., 2005), embryonic germ cells (Tada et al., 1997), and embryonal carcinoma cells (Do et al., 2007; Flasza et al., 2003; Han et al., 2008) are able to reprogram different somatic cells under appropriate experimental conditions. Some studies have investigated interspecies fusion reprogramming and mouse embryonic stem cells, for example, fused with human lymphocytes reactivated endogenous human pluripotency genes (Pereira et al., 2008).
Mouse embryonal carcinoma cells have the capacity to reprogram human T-lymphoma cells by reactivation of the human OCT4 and SOX2 genes (Flasza et al., 2003). Following the reprogramming events in fused cells, segregation of the tetraploid set of chromosomes is essential to obtain reprogrammed cells with a normal diploid set of chromosomes.
Here, we investigated the feasibility of interspecies reprogramming of porcine fibroblasts by murine ES cells with the goal to produce pluripotent diploid porcine cells that could then be differentiated into therapeutically useful cells. This first study of the fusion reprogramming of porcine somatic cells with murine embryonic stem cells resulted in, at least partial, epigenetic reprogramming of the porcine genome. However, the porcine chromosomes were progressively and preferentially eliminated from the tetraploid mouse–pig hybrid cells, and it does not appear that reprogrammed diploid porcine cells can be obtained by this method.
Materials and Methods
Cell cultures
The mouse ES cell lines RHN (Red fluorescent protein expression, Hygromycin resistance, Nanog overexpression) and HM-1 (hypoxanthine phosphoribosyltransferase deficient), and the tetraploid hybrid fused cells were maintained on gelatin-coated (0.1% gelatin; Sigma-Aldrich, Taufkirchen, Germany) dishes under feeder-free (for RHN ES) as well as feeder-including conditions (for HM-1 ES) in ESC medium containing Glasgow MEM (G-MEM) with L-glutamine (Invitrogen, Carlsbad, CA) supplemented with 10% fetal bovine serum (FCS; Invitrogen), 100 U/mL penicillin, 100 μg/mL streptomycin (PAA Laboratories GmbH, Pasching, Austria), 1% nonessential amino acids (PAA Laboratories GmbH), 50 μM 2-mercaptoethanol (Sigma-Aldrich) and 1000 U/mL leukemia inhibitory factor (LIF; ESGRO, Millipore, Billerica, MA). Primary porcine fibroblasts were obtained from a 25-day-old transgenic fetus as described (Kues et al., 2002). This fetus came from a German landrace animal and carried an expression cassette driving the human A20 and the neomycin gene (hA20/neo; Oropeza et al. unpublished). Only fibroblasts with less than four passages were used for fusion experiments.
Cell fusion and hybrid culture
In the first set of experiments, intraspecies mouse hybrids were produced by suspension/coculture fusion of RHN or HM-1 ES cells with murine primary fibroblasts from OG2/Rosa26 transgenic fetuses (day 13.5) carrying a germ line-specific Oct4 promoter-GFP construct (Kues et al., 2005). Hybrid colonies with GFP fluorescence were amplified.
In the second experiments, we adapted the interspecies fusion technology for porcine somatic cells and used HM-1 or RHN ES cells as the pluripotent partner to reprogram porcine somatic cells. The suspension fusion with use of polyethylene glycol (PEG) was performed as described (Do and Schöler, 2004). Coculture cell fusion was carried out as described (Islam et al., 2006) with minor changes. In brief, dispersed RHN or HM-1 cells (1.7 × 106–2 × 106) were plated on gelatin-coated 6-cm dishes containing ES culture medium for 1–2 h to allow attachment. Thereafter, dissociated porcine fibroblasts (1 × 106) were added to the adherent stem cells. Approximately 15 min later the culture medium was replaced by serum-free medium containing 100 μg/mL phytohemagglutinin-P (Sigma-Aldrich). The cells were coincubated at 37°C for 30 min. After washing once with Dulbecco's phosphate-buffered saline solution (PBS; Sigma-Aldrich), and removal of the wash solution, 1 mL of 50% polyethylene glycol 1500 (PEG 1500, Roche Applied Science, Mannheim, Germany) was pipetted onto each 6-cm dish for 2 min. After 2 min, the PEG was aspirated, the cells were gently washed four times with serum-free medium, and then incubated in the ES culture medium for 30 min at 37°C. At this point, the medium was replaced with fresh ES culture medium. Twenty hours after the PEG treatment, the cells were trypsinized and plated on gelatin-coated 10-cm dishes (or for HM-1-derived hybrids on mouse geneticin-resistant feeder cells) in ES culture medium. To initiate the selection process, the medium was supplemented 48 h later with geneticin (400 μg/mL; PAA Laboratories GmbH), and in case of RHN-derived hybrids also with hygromycin (400 μg/mL; PAA Laboratories GmbH). Geneticin eliminated nonfused mouse ES cells and hygromycin eliminated nonfused porcine fibroblasts.
The suspension/coculture fusions with HM-1 cells revealed colonies, which could not be expanded. After coculture fusion with RHN ES cell line and 12–14 days culture in selection medium, double-resistant hybrid cell colonies were subcloned. ES culture medium without antibiotics was used for expansion of any hybrid cells. This procedure was repeated 10 times to obtain two stable lines of fused cells. These primary expanded hybrids cells were designated “one time selected” hybrids. Aliquots of the expanded cell lines were frozen and used in the flow cytometry and spectral karyotyping experiments described below. Both of these subcloned hybrid lines were subjected to reselection using medium containing 400 μg/mL geneticin for an additional 24–26 days to eliminate revertant diploid mouse cells and to enrich for tetraploid hybrids and spontaneously segregated diploid porcine cells. The cells derived from this second selection step were defined as “reselected” hybrids. The “reselected” hybrids were expanded for a further 14 days without antibiotic selection and defined as “hybrids after release from selection” (Fig. 1A).
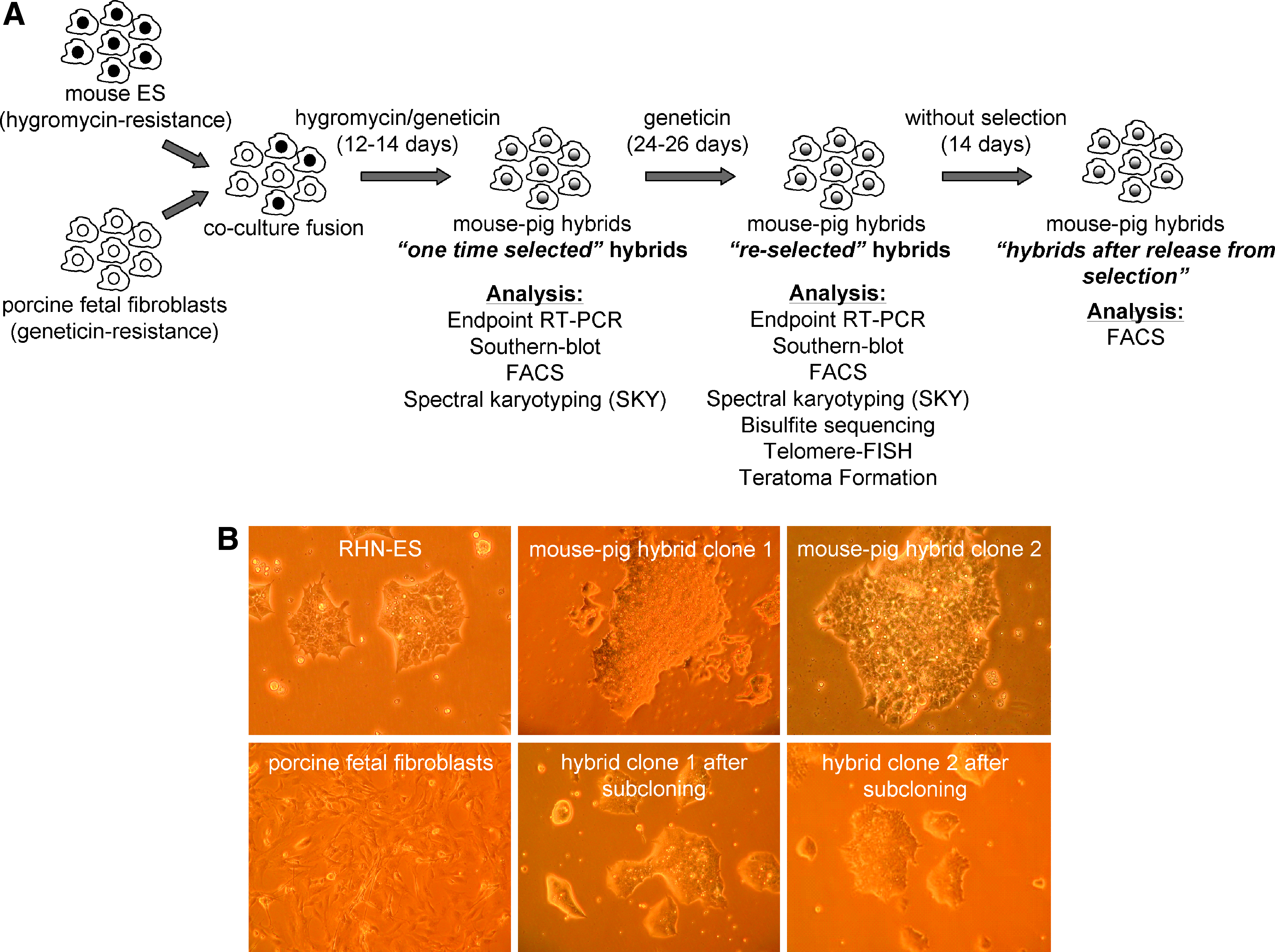
Interspecies cell fusion between mouse ES cells (RHN) and porcine fetal fibroblasts (hA20/neo transgenic). (
Endpoint reverse-transcription polymerase chain reaction (RT-PCR)
Total RNA from fused hybrids, RHN ES cells, and porcine fetal transgenic fibroblasts was extracted using Trizol Reagent (TRIsure; BIOLINE GmbH, Luckenwalde, Germany) and treated with RNase-free DNase (Epicentre Biotechnologies, Madison, WI). Poly(A)+ RNA was extracted from 10 matured porcine oocytes using the Dynabeads mRNA DIRECT Kit (Dynal®) and used immediately for reverse transcription. Negative controls were produced by adding sterile water instead of reverse-transcriptase and RNAse inhibitor. The PCR was performed with 160 ng of cDNA template (two oocyte equivalents) in a total volume of 50 μL using 1 U Platinum Taq DNA Polymerase (Invitrogen), 1 × PCR buffer (50 mM KCl, 20 mM Tris-HCl pH 8.4; Invitrogen), 200 μM of each dNTP (BIOLINE GmbH), 1.5 mM MgCl2 (Invitrogen), and 1.0 μM of each gene-specific primer (Table 1). The PCR program consisted of an initial denaturation step at 97°C for 2 min, followed by 36 cycles of 15 sec at 95°C, 30 sec at 52–60°C, 15 sec at 72°C, and a final elongation at 72°C for 5 min followed by cooling to 4°C. PCR products were separated on 2% agarose gels, visualized on a 312 nm UV-transilluminator, and recorded using CCD camera (Quanix, Photometrics, München, Germany). Amplified products were confirmed by the commercial sequencing service Agowa (Berlin, Germany).
Southern blot hybridization
Southern blotting was performed using DIG High Prime DNA Labeling and Detection Kit II (Roche). In brief, genomic DNA (10 μg) was digested using the restriction enzyme HindIII (New England Biolabs, Frankfurt am Main, Germany) and processed according to standard procedures. A DIG-labeled human A20/neo probe was employed to confirm that the transgene had integrated in the porcine genome.
Bisulfite sequencing analysis of genomic DNA
Genomic DNA (700 ng) was digested with NotI (New England Biolabs) and then denatured by addition of 2 M NaOH. The denatured DNA was embedded in 50 μL of 20 mg/mL dissolved low melting temperature agarose (BMA, Rockland, ME) and aliquots of 10 μL were pipetted into ice-cold mineral oil to form agarose beads. For bisulfite treatment, 2.5 M bisulfite-hydroquinone solution pH 5.0 (Roth Applied Science, Hastings, NE) was added to the mineral oil containing the beads and the reaction proceeded for 4 h at 50°C in a covered waterbath to exclude light. The DNA of each single bead was amplified in a separate PCR reaction. Primer sequences for bisulfite sequencing of both the murine Oct4 promoter and the porcine OCT4 promoter (primer 1) were designed with MethPrimer software (http://www.ucsf.ed/edu/urogene/methprimer/index1.html). For the second porcine primer, Methyl Primer Express v 1.0 software (Applied Biosystems) was used (Table 1). We analyzed the ratio between unconverted and converted cytosine. Only sequences with at least 90% C/T conversion were included. Methylation status was defined by comparing the cytosine and thymidin peaks in CG dinucleotides. We classified individual CpG methylation as low (<33%), medium (33–67%) or highly methylated (67–100%).
Ploidy analysis by flow cytometry
Fused mouse–pig hybrid clones 1 and 2, porcine fetal fibroblasts, and ES cells were analyzed for DNA content using flow cytometry. Prior to analysis, the cells were fixed overnight in ice-cold 70% ethanol and then washed with PBS containing 3% fetal bovine serum. They were then incubated with RNase A (1 mg/mL; Sigma-Aldrich, in PBS) and their DNA was stained with propidium iodide at a final concentration of 20 μg/mL (Sigma-Aldrich). All samples were prepared in duplicates. Propidium iodide fluorescence was analyzed using a FACScan (BD Bioscience, Heidelberg, Germany) equipped with an argon laser (488 nm, 15 mW).
Chromosome preparation and spectral karyotyping (SKY)
Metaphase chromosomes were prepared from fused mouse–pig hybrid clone 1 (passage 7; “re-selected” with geneticin for 24 days) and clone 2 (passage 8; “reselected” with geneticin for 26 days). Briefly, cells were treated with colcemid at a final concentration of 0.035 μg/mL overnight, incubated in 0.075 M KCl for 20 min at 37°C, and fixed in a freshly prepared mixture of methanol:acetic acid (3:1) at room temperature. Cell suspension was dropped onto glass slides in a climate chamber (Polymer, Kassel, Germany) at 22°C and 48% humidity and stained with Giemsa or used for SKY analysis.
SKY was performed as described previously (Rudolph and Schlegelberger, 2009) and according to the manufacturer's instructions (ASI; Applied Spectral Imaging, Ltd., Migdal HaEmek, Israel). Spectral images were acquired using an epifluorescence microscope equipped with an interferometer (SpectraCube™ ASI), a custom-designed optical filter and the SkyView™ software (ASI).
Telomere FISH
For qualitative analysis of telomeres using fluorescence in situ hybridization (FISH), slides with metaphase chromosomes were pretreated with RNase A and pepsin followed by overnight hybridization with a PNA probe (Applied Biosystems) in a humidified chamber. Images were acquired using an epifluorescence microscope equipped with FishView 2.0 software (ASI).
Teratoma formation assay
Reselected mouse–pig hybrid clone 1 passage 9 (4 × 106 cells pro injection) and clone 2 passage 10 (6 × 106 cells pro injection) and RHN ES cells passage 14 (2 × 106 cells pro injection) were subcutaneously injected in immunodeficient nude mice. After 4 weeks mice with visible tumor growth were sacrificed and the tumor tissues were fixed in 4% formaldehyde/succrose for histological examination.
Results
Production of intraspecies mouse–mouse hybrids
First, intraspecies fusion with murine OG2/Rosa26-transgenic fibroblasts were performed to confirm the reprogramming capacity of two different mouse embryonic stem cell lines, that is, the HM-1 and RHN ES in two different fusion methods (suspension and coculture fusion). Reactivation of Oct4–GFP in mouse–mouse hybrids could be demonstrated (Supplemental Fig. 1A–D). FACS analysis revealed that mouse–mouse hybrids were predominantly polyploid after subcloning and cultivation following termination of selection (Supplemental online Fig. 1E). The coculture system based on RHN fusion proved to be most efficient in preliminary intraspecies fusion. Among the three intraspecies fusion experiments produced with the HM-1 and RHN stem cell lines, the yield of hybrid colonies resulting from each fusion with the RHN ES cells ranged from 11–84 colonies, and the yield with HM-1 ES cells ranged from 1–8 colonies.
Production of interspecies pig–mouse hybrids
We generated interspecies hybrids by polyethylene glycol-mediated fusion employing two different methods (suspension and coculture fusion) using two different transgenic mouse embryonic stem cell lines (HM-1 and RHN ES) as pluripotent partners for fusion with primary porcine fibroblasts.
For interspecies cell fusion, porcine fetal fibroblasts (PFFs) were derived from a transgenic pig carrying the human A20 transgene and neomycin resistance (hA20/neo) to facilitate discrimination between murine and porcine cells. Viable hybrids could not be obtained from suspension/coculture fusion with HM-1 ES cells and suspension fusion with RHN ES cells even though at least 10 attempts were made. In coculture fusion experiments, performed with the RHN ES cells, only two clones survived selection (ten attempts). Both clones displayed typical ES cell morphology, formed ES-like colonies, and showed the high proliferation rate typical for embryonic stem cells (Fig. 1B). After subcutaneous injection of mouse–pig hybrids from clones 1 and 2 in four immunodeficient nude mice, three of them developed visible tumors within 4 weeks.
Interspecies hybrid verification
Species-specific gene expression analysis
To confirm the presence of the transgenic markers in the fused clones, endpoint RT-PCR was performed with primers specific for hA20/neo construct (contributed by the porcine genome) and for the hygromycin construct (contributed by the murine genome). Clones 1 and 2 were found to contain both the hA20/neo and the hygromycin transgenes (Fig. 2A). Confirmation of the porcine genotype in these hybrid cells was achieved by Southern blot analysis with a hA20/neo probe (Fig. 2B).

Gene expression analysis of interspecies hybrids. (

Gene expression analysis with species-specific primer combinations of stemness- related genes included OCT4, NANOG, SOX2, and STAT3. Expression of porcine OCT4 and STAT3 genes was detected in hybrid clone 1 (Fig. 2A). Surprisingly, in addition to the expected 322 bp OCT4 amplicon, a smaller product of 257 bp was observed in hybrid 1, but not in porcine oocytes. Sequencing results suggest that this smaller amplicon is the result of a cryptic splice site in the porcine OCT4 transcript, resulting in deletion of 65 bp of coding sequence and a frame shift mutation (Fig. 2C). No transcripts of the other two pluripotent markers were detected in either of the two hybrid lines (data not shown, see below).
Methylation status of the porcine and murine OCT4 promoters
To further investigate aspects of pluripotency in the interspecies hybrids, bisulfite analysis was used to determine the methylation status of the OCT4 promoter. The first step was to confirm the presence of four conserved regions (CR1, CR2, CR3, CR4) in the porcine OCT4 promoter (Nordhoff et al., 2001). The porcine CR1 region showed highest homology to the murine sequence (Fig. 3A). The porcine-specific primers designed for bisulfite-treated DNA gave a product that included 21 CpGs spanning CR1 and the adjacent exon 1; the murine-specific primers gave a product that contained 4 CpGs within the CR1 region of murine Oct4. The porcine OCT4 product was almost completely unmethylated in the hybrid cells of both “one time selected” and “reselected” samples from clone 1 (Fig. 3B), whereas the parental porcine fibroblast (controls) exhibited partial methylation of these CpG sites (Fig. 3B). Previous work indicates that there is high heterogeneity among primary fibroblast cultures with respect to OCT4 expression, and that expression levels are altered by culture conditions (Kues et al., 2005). Bisulfite analysis indicated that hybrid clone 1 was demethylated at porcine OCT4, which correlated with the active transcription demonstrated by RT-PCR analysis. No porcine OCT4 sequences could be amplified from hybrid clone 2 and, as discussed below, this may have been due to the loss of porcine chromosome 7 from the tetraploid cells.
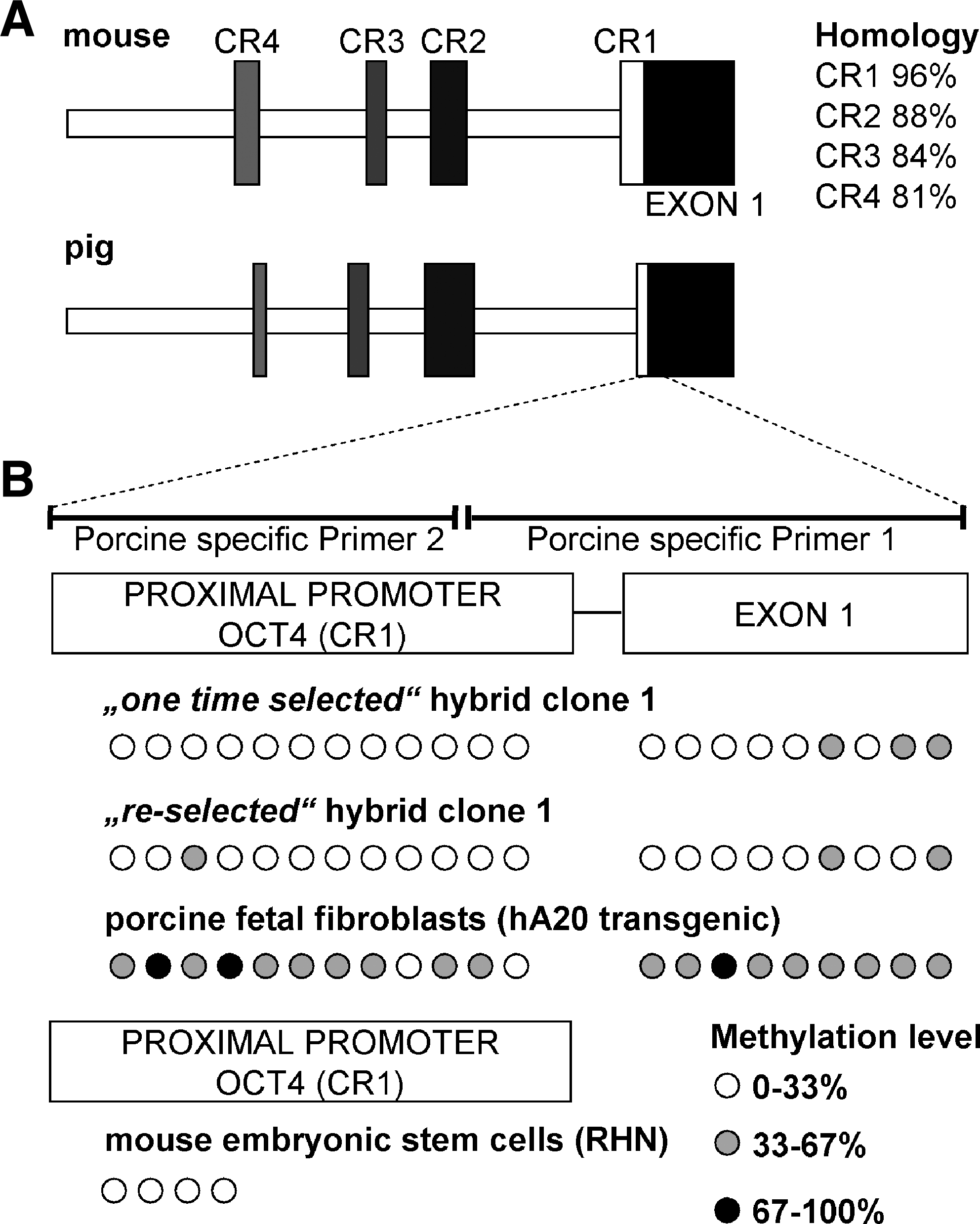
Conservation of murine Oct4 and porcine OCT4 promoter structure and methylation status. (
Ploidy analysis
To study ploidy of the hybrid cells, the DNA content was assayed by flow cytometry after staining with propidium iodide (Fig. 4). Nonfused fibroblasts and ES cells showed two distinct peaks corresponding to cell cycle stages G1 (2n, diploid) and G2/M (4n, tetraploid) (Fig. 4A and B). Cells from the two mouse–pig hybrid clones were analyzed after 2 weeks of antibiotic selection with geneticin and hygromycin (“one time selected hybrids”). At this stage, flow cytometry of hybrid clone 1 indicated the presence of diploid and tetraploid cells and a small population of polyploid cells (Fig. 4C). Hybrid clone 2, however, only gave 2n and 4n peaks, indicating that this hybrid was diploid rather than the expected tetraploid (Fig. 4D).

Ploidy status of interspecies hybrids. Flow cytometric histograms of propidium iodide stained nuclei: (
The second round of antibiotic selection with geneticin (“reselected” hybrids), diploid cells were eliminated, and polyploid cells were enriched in hybrid clone 1 (Fig. 4E). This seemed to indicate that the residual diploid cell population consisted either unfused or revertant murine cells. However, after release from geneticin selection, a decrease in the proportion of polyploid cells was detected in the “reselected” hybrid clone 1, suggesting that the fused polyploid cell population was unstable and prone to loosing chromosomes (Fig. 4G). After reselection of hybrid clone 2 and release from geneticin (Fig. 4F and H), ploidy plot was unchanged and characteristic of diploid cells. Together, these data indicate that mouse–pig hybrids could be produced, but that the hybrids are prone to loose chromosomes. This may happen either by segregation into diploid cells containing mouse or pig chromosomes or by random loss of individual chromosomes.
Karyotype analysis and telomere FISH of mouse–pig hybrids
To investigate chromosome composition of the interspecies hybrids, clones 1 and 2 were analyzed by spectral karyotyping
In the first set of SKY experiments with “one time selected” hybrids, they retained the mouse chromosome complements. However, clonal specific chromosomal aberrations were found in hybrid clone 1 including trisomy 8, a balanced translocation T(2;4), and loss of the Y chromosome. Furthermore, only a few unpainted porcine chromosomes or chromosomal fragments [that are described as marker chromosomes (mar)] were discovered in two metaphases of hybrid 1 (Table 2). To eliminate the revertant mouse cells and enrich the tetraploid hybrids as well as diploid pig cells, the fused cells were cultivated over the following 2 weeks under geneticin selection. Surprisingly, in the second sets of experiments with the “reselected” hybrids most of the clone 1 hybrids were nearly tetraploid, and SKY revealed two to four unpainted porcine chromosomes or chromosome fragments in nearly all metaphases analyzed (Fig. 5A). However, hybrid clone 2 did not change chromosome numbers after geneticin reselection, which is consistent with the flow cytometry results. Interestingly, even though this cell line did not exhibit any free porcine chromosomes, a translocation between porcine chromosomal material and mouse chromosome X in the majority of metaphases was observed. The pig chromosomal material was detectable as an unpainted fragment in the painted mouse X chromosome of hybrid clone 2 (Fig. 5B). This could explain their survival of the selection process. Telomere FISH analysis detected the telomere of both murine and porcine chromosomes due to their identical telomere repeat sequence TTAGGG. The derivative X chromosome in hybrid clone 2 showed six telomere signals (Fig. 5C and D). Interestingly, two strong signals could be seen in the centromeric region of the X chromosome. Two equally strong signals were seen in the distal part of the chromosome and two minor signals, perhaps relating to the telomeric instability, which may have permitted the fusion of a porcine fragment into the murine X chromosome.
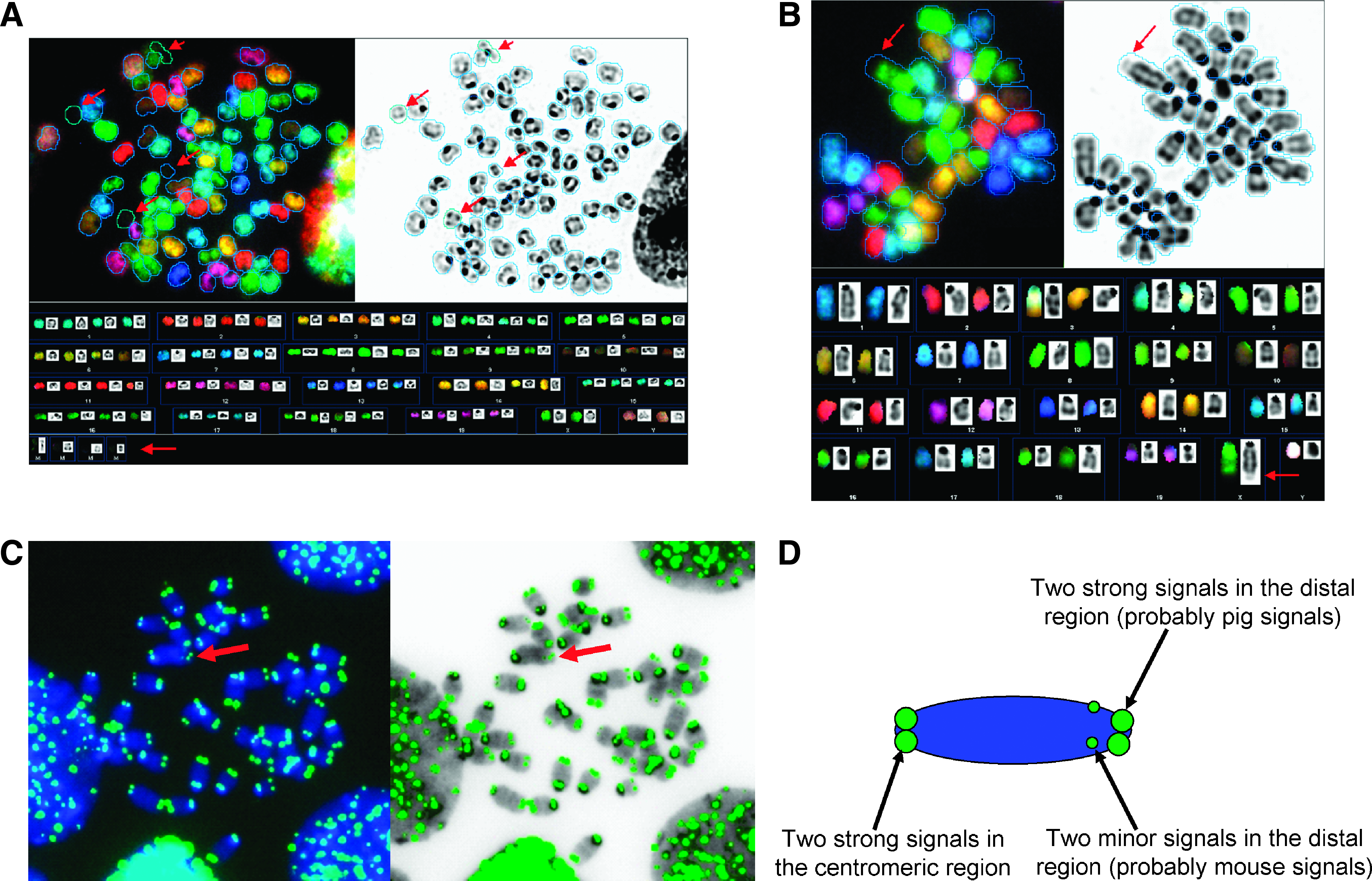
Spectral karyotyping and telomere FISH of interspecies hybrid cell lines. RGB [red–green–blue image (upper left), inverted DAPI image (upper right) and karyograms (below)]. (
Loss of single chromosomes in individual metaphases without clonality; mar, unpainted porcine chromosomal fragments/marker chromosomes.
[n], number of metaphases; XF, fused murine X and porcine chromosomal fragment.
Discussion
Cell fusion with pluripotent cells can reprogram the somatic genome. For therapeutic application, the tetraploid hybrids should eventually resegregate into stable diploid cells with a normal set of chromosomes. This approach could be used to obtain pluripotent cells of a desired genotype or species, and could theoretically be based on interspecies cell fusion. Determination of the chromosome composition in intraspecies hybrid cells is difficult using only morphological criteria for discrimination (Matveeva et al., 2005).
In the current interspecies cell fusion experiments with murine embryonic stem cells and porcine fibroblasts, we were able to monitor aspects of chromosomal stability and reprogramming. Spectral karyotyping with murine specific probes was particularly helpful for discriminating between murine and porcine chromosomes at the single-cell level. Porcine pluripotent cells will be valuable for cell transplantation experiments, as pigs are a favored model organism in preclinical studies, due to their similarity to humans with respect of size, life span, physiology, and immune systems (Kues and Niemann, 2004; Vackova et al., 2007). Unfortunately, at the time of writing, true porcine germline ES cells are not available (Brevini et al., 2007; Chen et al., 1999; Li et al., 2004; Piedrahita et al., 1990; Strojek et al., 1990; Vackova et al., 2007). To address this problem, we undertook an investigation into the feasibility of producing mouse–pig fusion hybrids as a source of reprogrammed stem cells. We looked at the possibility that such fusion hybrids spontaneously resegregate into diploid cells with their original species specific complement of chromosomes.
This is the first study that attempted to reprogram differentiated porcine cells by fusion with murine embryonic stem cells. Unfortunately, the ultimate goal of producing pluripotent porcine stem cells was not realized. Interspecies fusion between mouse and pig cells was so transient that only two hybrid cell lines were obtained. The two clones examined had rapidly lost almost all porcine chromosomal material.
The initial results were encouraging, as the clones did display characteristics of embryonic stem cells including colony morphology and proliferation rate. The hybrids clearly expressed the transgene markers for both parent cell lines, demonstrating that they were the result of interspecies fusion. OCT4 is well known for its importance in reprogramming (Do and Schöler 2004; Han et al., 2008; Ma et al., 2008; Tada et al., 2001), and deletion of OCT4 diminishes the reprogramming capacity of mouse embryonic stem cells (Pereira et al., 2008). For this reason, OCT4 is commonly used for positive identification of toti- and pluripotent cells (Tada et al., 2001). Transcriptional reactivation of Oct4 and other pluripotency markers has been observed after interspecies fusion between murine embryonal carcinoma cells and human T-lymphoma cells (Flasza et al., 2003) and between murine embryonic stem cells and human lymphocytes (Pereira et al., 2008). This indicates that somatic cells can be reprogrammed toward a pluripotent state by the fusion of differentiated human cells with a pluripotent partner from a different species.
In the present study, reactivation of the porcine OCT4 gene was evident in hybrid clone 1. Interestingly, a novel, additional, shortened amplicon, representing a nonsense splice variant was detected in hybrid 1. We suppose that this splice variant might be associated with faulty processing of porcine pre-mRNA in the interspecies hybrids. The splicing of 65 bases occurred at a cryptic splice site of the porcine OCT4 transcript, and resulted in the deletion of some coding sequence and a frame shift mutation, which would result in translation of a nonfunctional protein. Splicing errors could be a critical basic problem in cross-species fusion reprogramming.
The methylation status of the proximal OCT4 promoter correlated well with the observed expression pattern of the OCT4 gene, suggesting functional reprogramming of this gene in hybrid clone 1. The difference between the nonmethylated CpG sites in hybrid clone 1 and the distinctly methylated sites in the original porcine fetal fibroblasts is striking. The OCT4 promoter is hypermethylated in adult mouse somatic cell populations including fetal fibroblasts, cumulus cells, and cells from adult liver or spleen (Marikawa et al., 2005). Unfortunately, other pluripotent markers were not detected in the interspecies hybrids, probably due to the rapid exclusion of porcine chromosomes. Interestingly, loss of OCT4 expression during colony expansion was observed in human–mouse hybrids (Flasza et al., 2003).
Two weeks after fusion, small tetraploid cell populations could be identified in hybrid clone 1. Antibiotic reselection of hybrid clone 1 cells resulted in elimination of the residual diploid cell population. This, together with the SKY data, suggests that selective elimination of porcine chromosomes had taken place.
Hybrid clone 2 remained diploid, that is, ploidy was unchanged by reselection. It was interesting to note that most of hybrid clone 2 metaphases carried a translocation, which placed some porcine material most likely with the geneticin resistance gene, on the murine X chromosome. Previous studies have shown that hybrid cells are frequently unstable and rapidly lose supernumerary chromosomes (Matsui et al., 2003; Matveeva et al., 1998; Wang et al., 1998). Moreover, the stability of hybrid karyotypes appears to be dependent on the developmental state of the somatic fusion partner (Matveeva et al., 2005). In some combinations, in vitro culture of fused cells has revealed that one chromosome complement of the fused partner cells often is dominant in the interspecies hybrids (Chinchar et al., 1979; Croce et al., 1976). In other cases, the loss of chromosomes from both fusion partners (mouse and human chromosomes) was reported (Wang et al., 1998). Stable tetraploid hybrids have only been obtained by the fusion of cells from two different embryonic stem cell lines (Matveeva et al., 2005).
SKY analysis clearly indicated preferential loss of porcine chromosomes in the two clones that we examined and retention of mouse chromosomes in culture. In hybrid clone 1, we observed a tetraploid set of mouse chromosomes and four unpainted porcine chromosomes or chromosomal fragments. Hybrid clone 2 did not exhibit any free porcine chromosomes. Translocation between porcine chromosomal fragments and the mouse chromosome X was observed. Mechanisms involved in the preferential loss of chromosomes from one fusion partner or chromosomal translocation in interspecies hybrids have not yet been investigated; differences in the initial rate of cell division of the two fusion lines leading to asynchronies in cell division may account for an early loss of chromosomes. Incompatibilities between the porcine and mouse genome after interspecies cell fusion could cause splicing errors and loss of porcine chromosomes from hybrids.
In conclusion, the data presented here demonstrate that fusion did occur between the porcine murine cell types that the murine genome was more stable in the resultant hybrids, and that reprogramming of the porcine OCT4 gene had taken place as a result of the fusion event. To produce pluripotent porcine cells by fusion, it may be important to select subsets of cells that are more compatible with a particular murine ES cell line. For example, it may be important to select cell lines that have the same rate of cell division or to perform fusion after the cell lines have been synchronized to be in the same stage of the cell cycle. Under the conditions used in this study, it seems clear that there is rapid loss of porcine chromosomes in the interspecies hybrid cells. Determination of the time course of chromosome loss would be interesting, but will require performing a large number of additional experiments. In addition to rapid chromosome loss, the observation of defective posttranscriptional processing of the porcine OCT4 mRNA indicates additional molecular incompatibilities, which may exclude murine ES cells as a practical tool for reprogramming differentiated porcine cells. Future studies will focus on answering some of these important questions.
Footnotes
Acknowledgments
The authors thank Brigitte Barg-Kues, Doris Herrmann, and Andrea Schienke for excellent technical assistance and gratefully acknowledge the supply of hA20/neo transgenic porcine fibroblasts by Marianne Oropeza. Our sincere thanks also go to Dr. Andrea Lucas-Hahn for collaborative work on electrofusion experiments and Thomas Buchholz for providing the teratoma assay with nude mice. We thank Dr. Tobias Cantz (Max Planck Institute, Münster) for providing the HM-1 embryonic stem cell line and advice about cell fusion, and Dr. Tomo Sarič (Institute of Neurophysiology, Medical Center, University of Cologne) for providing RHN ES cells. The OG2/Rosa26 transgenic mouse line was a kind gift of Prof. Hans Schöler. This study was supported by a grant from the Bundesministerium für Bildung und Forschung (BMBF) to H.N and by the Excellence Cluster Rebirth (H.N., B.S., C.R.).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.