
Abstract
Abstract
The goals of the current study were to (1) improve culture conditions and (2) chemical passaging of bovine embryonic stem cell-like (bESC-like) cells. Specifically, the effects of human leukemia inhibitory factor (hLIF), two types of feeders, mouse embryonic fibroblast (MEF) and bovine embryonic fibroblast (BEF), as well as three different enzymatic treatments including Trypsin-EDTA, TrypLE, and Liberase Blendzymes 3 were investigated. The addition of hLIF at 1000 U/mL to the culture medium (41.2 and 36.9%), and the use of either MEF or BEF feeders (40.3 and 38.1%) had no significant effect on the ability of inner cell masses (ICMs) to form primary cell colonies compared to controls. All bESC-like cells were first dissociated mechanically for three passages followed by enzymatic dissociation. The ability to maintain ESC morphology to passage 10 was compared among the three enzymes above. More bESC-like cell lines survived beyond passage 10 when treated with TrypLE compared to Trypson-EDTA (28.8 and 12.6%; p < 0.05), and bESC-like cells differentiated quickly when treated with Liberase Blendzyme 3. The bESC-like cells generated in our study displayed typical stem cell morphology and expressed specific markers such as SSEA-1, AP, OCT-4, and Nanog. When removed from feeders, these bESC-like cells formed embryoid bodies (EBs) in a suspension culture. When EBs were cultured on tissue culture plates, they differentiated into various cell types. In summary, we were able to culture bESC-like cells more than 10 passages by enzymatic dissociation, which is important in gene targeting, maintenance, and banking of bESC lines.
Introduction
The derivation and maintenance of ESC lines from bovine blastocysts is perhaps the most difficult and challenging among all mammals. To date, suitable culture conditions for preventing spontaneous differentiation of bESCs have not been established despite the extensive efforts devoted to the generation of stable bESC or ES-like cell lines (Cibelli et al., 1998; Iwasaki et al., 2000; Mitalipova et al., 2001; Saito et al., 2003; Stice et al., 1996; Strelchenko 1996; Wang et al., 2005). Leukemia inhibitory factor (LIF) is a cytokine that was successfully used in the derivation of mouse ESCs (Evans and Kaufman 1981; Moore and Piedrahita 1997). However, it is dispensable for human ESCs (Xu et al., 2005). Previous reports have documented conflicting results on the effect of human LIF (hLIF) in the derivation and maintenance of bESC-like cells (Mitalipova et al., 2001; Saito et al., 2003; Vejlsted et al., 2005).
Many different cell types, including homologous and heterologous fetal fibroblasts, have been tested for use as feeder cells during the derivation and maintenance of ESCs. The use of homologous primary feeder cells would be expected to be a better choice by producing homologous LIF and other growth factors or cytokines to maintain pluripotency. Homologous fetal fibroblasts have been successfully used as feeder cells to derive ESCs in the mink (Sukoyan et al., 1993), pig (Li et al., 2004), and humans (MaLi et al., 2008). To our knowledge, it is not clear if bovine embryonic fibroblasts (BEF) are more suitable than mouse embryonic fibroblasts (MEF) as feeder cells, for either the establishment or the maintenance of bESC in the undifferentiated pluripotent state.
So far, all reported bESC-like cells were passaged mechanically (Cibelli et al., 1998; Iwasaki et al., 2000; Mitalipova et al., 2001; Saito et al., 2003; Stice et al., 1996; Strelchenko 1996; Wang et al., 2005), which is not only tedious and technically challenging, but may also hamper bESCs' applications such as gene targeting and banking. In the present study, we (1) investigated the effects of hLIF and two types of feeders, MEF and BEF, on bESC-like cells derivation and (2) tested the feasibility of passaging bESC-like cells by different enzymatic treatments including Trypsin-EDTA, TrypLE, and Liberase Blendzymes 3.
Materials and Methods
Experimental design
Experiment 1 was to investigate effects of hLIF on bESC-like cells derivation and maintenance. For derivation, day 7 bovine in vitro fertilized (IVF) blastocysts were randomly divided into two groups, and transferred to four-well dishes with MEF feeder cells with or without hLIF(1000 U/mL). To test the effect of homologous feeder cells, BEF, on bESC-like cells derivation, day 7 bovine IVF blastocysts were randomly divided into two groups, and transferred to four-well dishes with either MEF or BEF as feeder cells, and with all cultured in ES medium with (1000 U/mL) hLIF. Following mechanical dissociation for three passages, bESC-like cells were passaged by Trypsin-EDTA treatment for continued culture.
Experiment 2 was to test different enzymatic treatments on passaging of bESC-like cells. Day 7 bovine IVF blastocysts were transferred to four-well dishes with MEF feeder cells. They were all cultured in ES medium with (1000 U/mL) hLIF. Following mechanical dissociation for three passages, bESC-like cell lines were randomly divided into three groups, and passaged by three different enzymatic treatments including Trypsin-EDTA, TrypLE, and Liberase Blendzymes 3 for continued culture.
In vitro fertilization and embryo culture
Frozen semen of a Holstein bull was obtained from Cooperative Resources International (Shawano, WI), and matured oocytes were provided by Evergen Biotechnologies, Inc. (Vernon, CT, USA). In vitro fertilization was conducted following a standard protocol established in our lab. Briefly, after a 10-sec gentle shaking, the semen straw was thawed for 10 sec in a 37°C water bath. Sperm were washed twice by centrifugation (1500 rpm) for 8 min in 10 mL of Brackett and Oliphant (BO) sperm-wash medium (Brackett and Oliphant, 1975) containing 3 mg/mL of bovine serum albumin (BSA, Sigma, St. Louis, MO, USA) supplemented with 10 mM caffeine (Sigma). The washed sperm pellet was then resuspended in the BO sperm-wash medium at a concentration of 1.0 × 106 sperm/mL for subsequent IVF. After maturation in vitro, bovine cumulus–oocyte complexes were washed twice and transferred into a 50-μL drop of BO medium (20–25 oocytes/drop) containing 6 mg/mL of BSA (Sigma) and 10 μg/mL of heparin (Sigma) preequilibrated for 2 h at 39°C in 5% CO2 in humidified air, and 50 μL of sperm suspension was added to each drop subsequently. Oocytes were incubated with sperm for 6 h at 39°C in 5% CO2 in humidified air.
After fertilization, embryos were further cultured in CR1aa medium (Rosenkrans and First, 1994) supplemented with 6 mg/mL of BSA (Sigma) for 48 h in an atmosphere of 5% O2, 5% CO2, and 90% N2. Cleaved embryos were then selected and cultured for an additional 5 days in CR1aa supplemented with 5% (v/v) fetal bovine serum (FBS, Hyclone, Logan, UT, USA) on cumulus cell monolayers in an atmosphere of 5% CO2 in air. The medium was changed every 2 days throughout the culture period.
Preparation of feeder cells
MEFs were obtained from the embryos of CF-1 mice 13–14 days post coitum. BEFs were obtained from a Holstein fetus at day 140 of gestation. Embryonic fibroblasts were isolated from these fetuses by methods described previously (Gong et al., 2004). Briefly, tissue biopsies were obtained from the skin and washed twice with DPBS (Invitrogen, Carlsbad, CA, USA). The tissue was cut into small piece, and cultured in a 25 cm2 tissue culture flask containing 6 mL DMEM (Invitrogen) supplemented with 10% FBS; Hyclone) and incubated at 37°C, 5% CO2 in a humidified atmosphere. When the cells reached confluency after 6∼7 days, they were passaged twice and frozen in DMEM supplemented with 20% FBS and 10% dimethylsulfoxide. For preparation of feeder cells, both MEF and BEF were inactivated by 10 μg/mL mitomycin C (Sigma) for 3.5 h, and washed five times in phosphate-buffered saline (PBS), and then reseeded at a density of 2 × 104 cells/cm2.
bESC-like cells isolation
Day 7 IVF blastocysts were transferred onto MEF or BEF in four-well dishes containing ESC culture medium. ES culture medium consisted of knockout DMEM (Invitrogen), supplemented with 20% Knockout Serum Replacement (KSR, Invitrogen), 0.1 mM 2-mercaptoethanol (Chemicon, Temecula, CA, USA), 2 mM L-Glutamine (Chemicon), 1% nonessential amino acids (Chemicon), 20 ng/mL bovine basic fibroblast growth factor (R&D Systems, Minneapolis, MN, USA) and additionally with or without 1,000 U/mL hLIF (Chemicon). The blastocysts hatched and attached in 1–3 days and ICM-derived colonies from all groups were mechanically picked and placed into new four-well dishes with MEF or BEF feeder cells. For the first three passages, bESC-like colonies were dissociated by mechanical dissecting bESC colonies into a number of small sized clumps using two 21G needles with the small sized clumps placed into new four-well dishes with feeder cells.
Enzymatic passaging of bESC-like cells
Beyond passages 3, the bESC-like colonies were picked, placed in a 50 μL drop of ESC culture medium, transferred into a 50 μL drop containing one of three enzymes including 0.05% trypsin-EDTA (Invitrogen), TrypLE express (Invitrogen) and 0.15 units/ml Liberase Blendzymes 3 (Roche, Indianapolis, IN, USA), and then pipetted rapidly for approximately 30 sec for dissociation. A 50-μL drop of ESC culture medium plus 10% FBS was then added to the cell suspension for enzyme inactivation. The dispersed cells were transferred into the ESC culture medium in new four-well dishes with MEF or BEF feeder cells.
When there were more than 30 colonies from each blastocyst, enzymes were added directly to the bESC-like cell culture dishes without picking up colonies for passaging. Briefly, ESC culture medium was removed, and 1 mL enzyme solution was applied to bESC-like cells cultured in one well of six-well dish. After approximately 3 min of incubation at 37°C, 500 μL of ESC culture medium with 10% FBS was added for enzyme inactivation, and the solution was pipetted vigorously with a 1000 μL pipette to break up the colonies. The cells were centrifuged and washed with DPBS once, and then transferred into the ESC culture medium in new six-well dishes with feeder cells.
Characterization of bESC-like cells
Bovine ESC-like cells at passages 5–7 from all groups were used to do characterization analysis including immucytochemistry, alkaline phosphatase, karyotype, and reverst transcriptase-polymerase chain reaction (RT-PCR) because more bESC-like colonies can be harvested at these passages. Immunocytochemistry was conducted to detect SSEA-1, Oct-4, and Nanog. After the cells were fixed for 15 min with 4% paraformaldehyde in PBS, they were incubated in blocking solution (15% goat serum) overnight at 4°C. Primary antibodies used including SSEA-1 (mouse anti-SSEA-1, MC-480 from Santa Cruz Biotechnology, Santa Cruz, CA, USA), OCT-4 (mouse anti-OCT-4, Clone 9E3 from Chemicon), and Nanog (Rabbit anti-Nanog, AB9220 from Chemicon) were diluted to 1:100 in blocking solution and incubated with cells for 1.5 h at room temperature. Alexa Fluor 488 goat antimouse IgG, IgM (for SSEA-1 and OCT-4, Invitrogen) or Alexa Fluor 594 goat antirabbit IgG (for Nanog, Invitrogen) were diluted to 1:500 in blocking solution and incubated with cells for 1 h. A total of 100 ng/mL of 4′,6′-diamidino-2-phenylindole hydrochloride (DAPI; Sigma) was incubated with cells for 10 min to visualize DNA. Expression of alkaline phosphatase (AP) was evaluated by alkaline phosphatase substrate Kit I (Vector Lab, Southfield, MI, USA). Karyotyping was conducted following the procedure established in our lab previously (Wang et al., 2005). Briefly, cells prepared for cytogenetic analysis were incubated in growth media supplemented with 0.2 μg/mL of colecemid solution (Gibco, Rockville, MD, USA) for 1 h. Cells were trypsinized, treated with 0.75 M KCl solution for 25 min at 37°C, fixed with methanol: glacial acetic acid (3:1, v/v, Sigma), and drops of cell suspension were spread on clean slides. The chromosomes were stained with 5% Giemsa (Sigma) for 40 min.
Expression of pluripotent factors were also detected by RT-PCR. Total RNA was extracted from bESC colonies picked from feeder cells, and reverse transcribed into cDNA using Cells to cDNA kit (Ambion, Austin, TX, USA) according the manufacture's instruction. The cDNA was then subjected to PCR analysis for gene expression of OCT-4, Nanog, and Gdf-3. Bovine mRNA sequences were obtained from NCBI database and primers were designed by Primer Express 3.0 (Applied Biosystems, Foster City, CA, USA). The following primers were used for PCR: OCT-4, 5′-GGTGGAGGAAGCTGACAACAA-3′ and 5′-CTCGTCCGCTTTCTCTTTCG-3′; Nanog, 5′-CCCCGAAGCATCCAACTCTA-3′ and 5′-GTGTCGAGGGTGTCAGCAGAT-3′; Gdf-3, 5′-ACGGAGCCCCATGTCATAGA-3′, and 5′-GCCACATCCAGCAGGTTGA-3′; 18sRNA, 5′-GCCCGAAGCGTTTACTTTGA-3′ and 5′-CCGCGGTCCTATTCCATTATT-3′. Amplifications yielded products of 87 bp (OCT-4), 90 bp (Nanog), 110 bp (Gdf-3), and 93 bp (18sRNA).
In vitro differentiation
Putative ES colonies were picked from the feeder layers and cultured as clumps in bacterial dishes containing ES medium without bovine basic fibroblast growth factor, hLIF, or feeder cells for embryoid body (EB) formation. The cells were fed every other day, and the suspension cultures were monitored daily for the formation of simple EBs. For in vitro differentiation, simple EBs were trypsinized and seeded in a gelatin-coated four-well dish.
Statistical analysis
An unpaired t-test was used to reveal differences in the number of embryos developing to primary colonies. A value of p < 0.05 was considered to be statistically significant.
Results
Most blastocysts (83%) hatched (Fig. 1A) and attached (Fig. 1B–C) in 1–3 days and 51% of them went on to form ICM-derived colonies. ICM-derived colonies from all groups were mechanically picked and placed in new cultures (Fig. 1D). When the primary colonies were separated and reseeded in the culture, new colonies were produced within 5–6 days of subculturing by dissecting bESC colonies into a number of small-sized clumps mechanically using two needles for the first 3 passages (Fig. 2A–B). These colonies grew slowly, requiring an average of 8–10 days between passages. These cells also had a high nucleus cytoplasm ratio, and formed domed colonies that were densely packed with obvious borders (Fig. 1E–F). No morphological difference was found among bESC-like colonies derived from all different groups. As shown in Table 1, most ES colonies differentiated or ceased growth after treatment of trypsin before the fifth passage (Fig. 1G–I). Less than 30% of bESC-like cells can be cultured more than 10 passages when treated with enzymatic dissociation (Tables 1 and 2).

bESC-like cells derivation. A day 7 bovine IVF blastocyst (
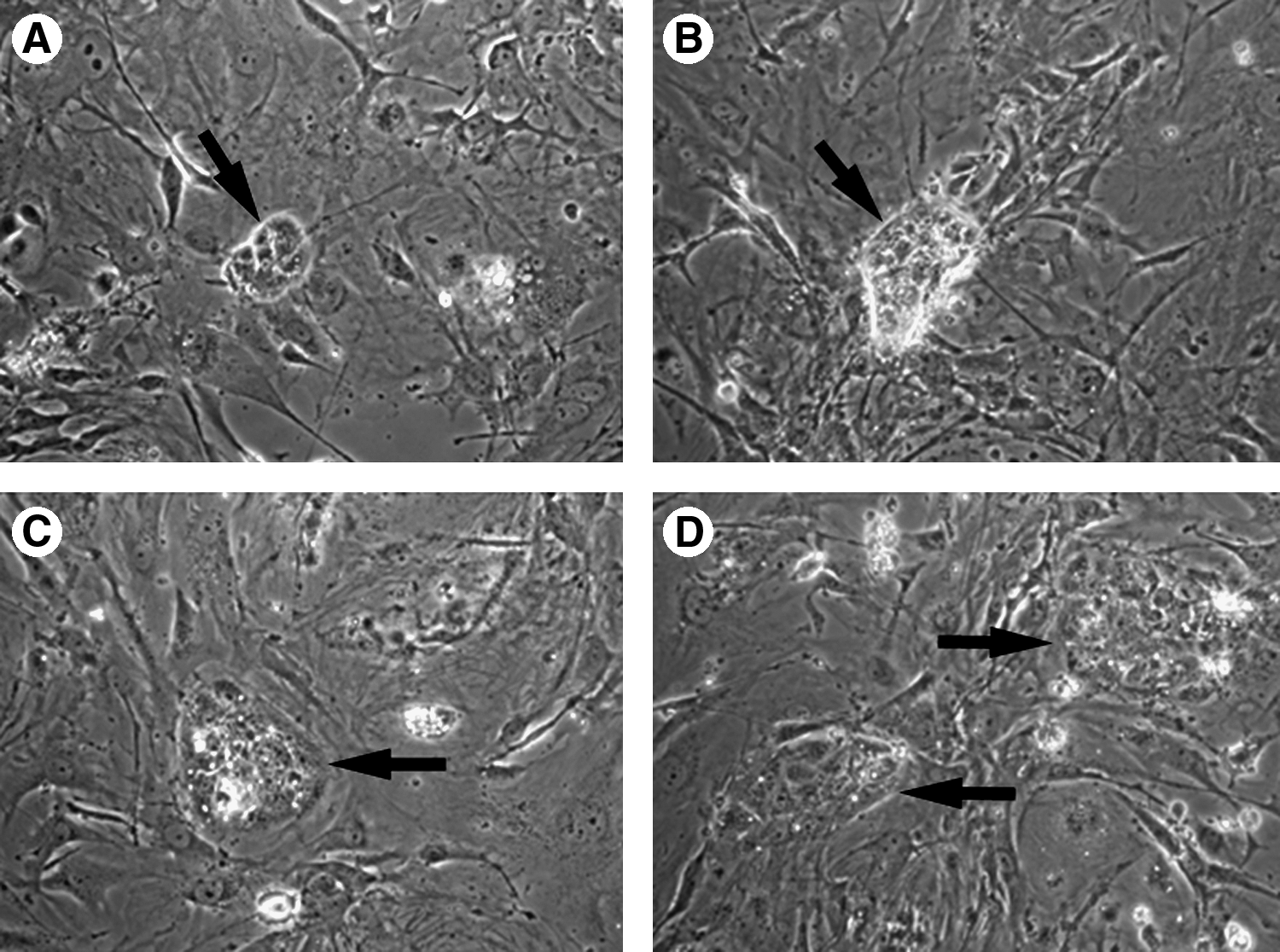
bESC-like colonies (
Based on the numbers of ICMs.
ICM, inner cell mass; LIF, leukemia inhibitory factor; MEF, mouse embryonic fibroblast; BEF, bovine embryonic fibroblast.
Different superscripts are significantly different (p < 0.05).
Addition of hLIF (1,000 U/mL) to the culture medium had no significant effect on the ability of ICMs to form primary cell colonies compared to controls (41.2 and 36.9%, respectively; Table 1). Also, no significant difference in primary colony formation was detected between ICMs cultured on MEF or BEF feeders (40.3 and 38.1%, respectively, Table 1). Following mechanical dissociation for three passages, more bESC-like cell lines were passaged to passage 10 when treated with TrypLE compared to being treated with Trypsin-EDTA (28.8 and 12.6%, respectively, p < 0.05, Table 2), and bESC-like cells differentiated quickly when treated with Liberase Blendzyme 3.
Expression of SSEA-1, OCT-4, and Nanog in ES colonies at passages 5–7 derived from all groups was detected by immunocytochemical staining (Fig. 3), and AP was also detected (Fig. 4). Karyotype analysis of bESC-like cells at passages 5–7 from all groups showed the expected number of normal chromosomes (Fig. 5). The RT-PCR results indicated that mRNA for OCT-4, Nanog, and Gdf-3 were detected in bESC-like cells at passages 5–7 (Fig. 6), further confirming expression of ESC transcription factors.
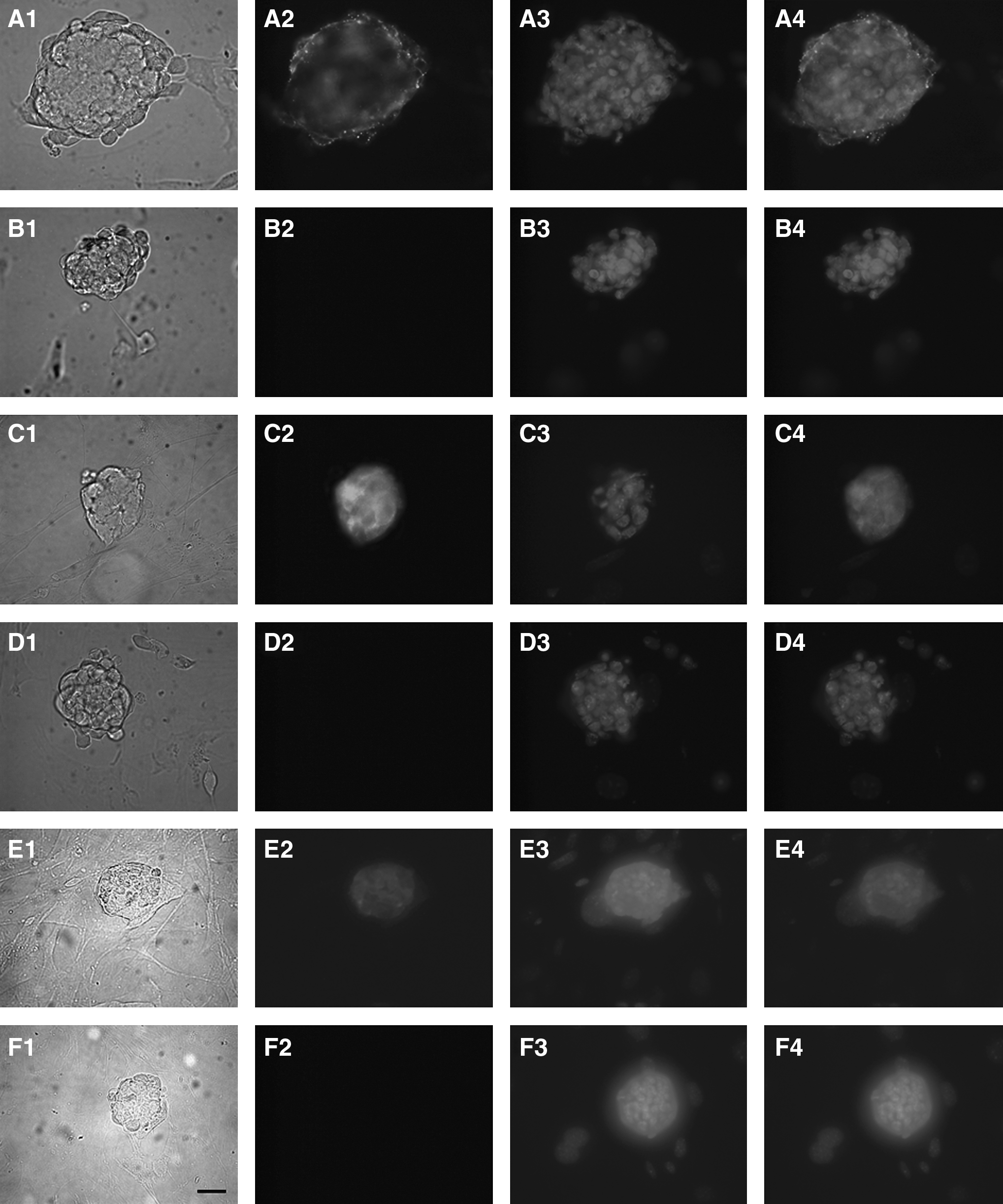
Expression of markers indicative of pluripotency. bESC-like colonies (

Alkaline Phosphatase (AP) staining of bESC-like colony. A representative colony stained red (

A representative metaphase spread of bESC-like cells showing 60 chromosomes.

RT-PCR analysis of bESC-like cells. M, molecular weight markers; 1: OCT-4 (87 bp); 2: Nanog (90 bp); 3: Gdf-3 (110 bp); 4: 18S rRNA (93 bp), 5: OCT-4 (reverse transcriptase negative); 6: Nanog (reverse transcriptase negative); 7: Gdf-3 (reverse transcriptase negative); 8: 18S rRNA (reverse transcriptase negative).
To test the differentiation potential of bESC-like cells, putative bESC colonies were cultured in bacterial dishes containing ES medium without bovine basic fibroblast growth factor, hLIF, or feeder cells for EB formation. Simple EBs were formed within a few days (Fig. 7A), and developed to cystic EBs when these cultures were maintained for 1–2 weeks (Fig. 7B). When simple EBs were plated onto tissue culture plates, the cells differentiated into various types of cells (Fig. 7C–H).
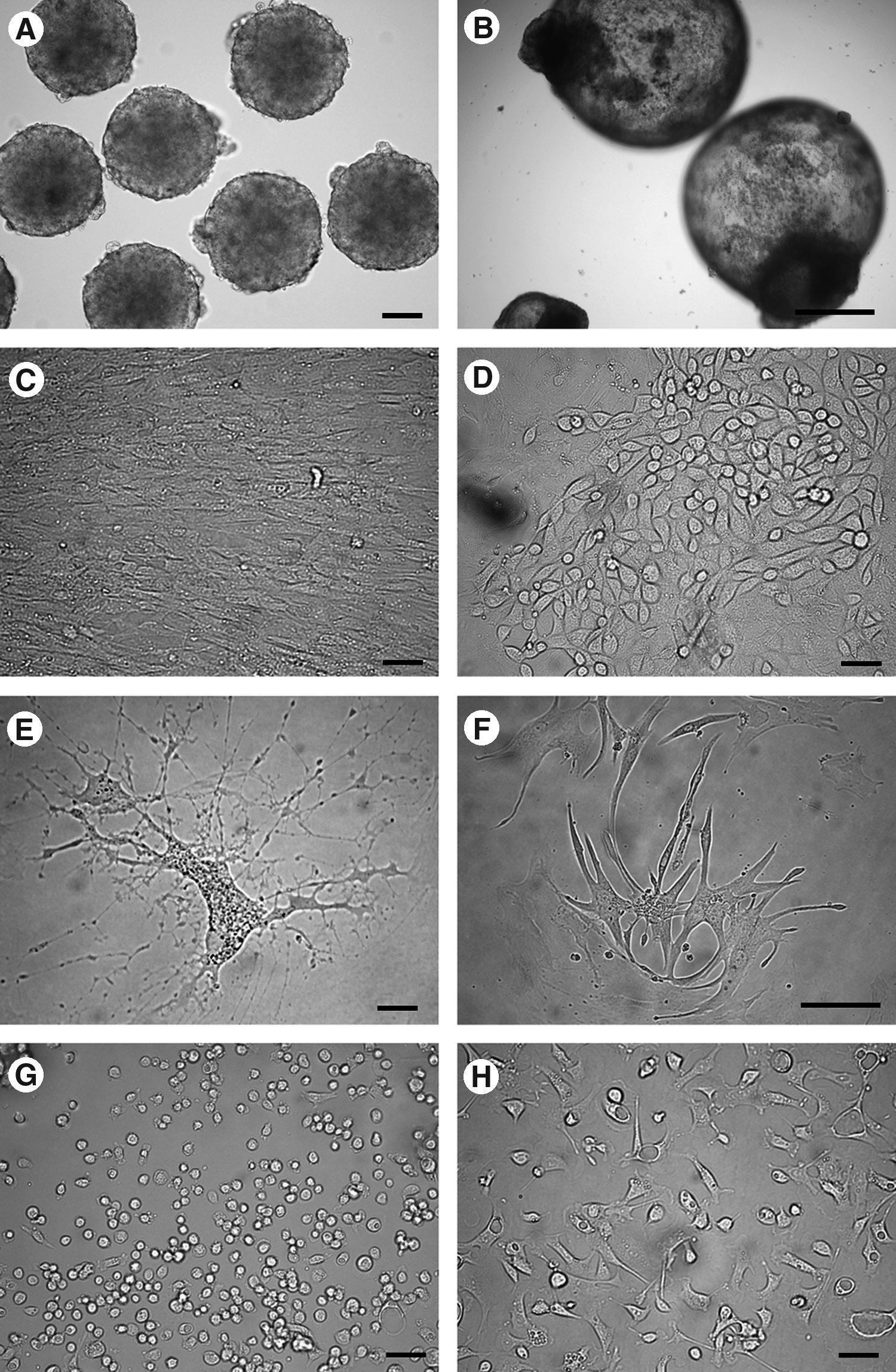
In vitro differentiation of bESC-like cells. (
Discussion
In addition to establishment of animal models for human genetic diseases and transplantation therapies, domestic animal ESCs could also be employed for precise genetic engineering of animals for use as a bioreactor, for disease resistance, and for improving production traits. In this context, the establishment of efficient methods for the derivation and maintenance of ES cell cultures from domestic animals would be of great interest. However, a culture method suitable for deriving and maintaining undifferentiated state of ES cells in domestic mammals is still not available. In the current study, we found that neither the presence of hLIF in culture medium nor the use of BEF as feeders had significant benefits to bESC-like cells derivation or maintenance of pluripotency. The primary colony formation rate in this study was around 40%, which is within the range of reported colony formation (13–67%) (Stice et al., 1996; Strelchenko 1996; Talbot et al., 1995). However, we were able to passage bESC-like cells by enzymatic dissociation, which is important in applications such as gene targeting as well as facilitating maintenance and banking of bESC-like cells following derivation.
The maintenance of pluripotency is perhaps one of the greatest challenges in bESC-like cells. Primate ES cells do not require LIF or bone morphogenetic protein 4 (BMP4) as mouse ES cells do. Rather, they require nodal/activin and fibroblast growth factor (FGF) (Renard et al., 2007). Our results showed no apparent beneficial effect of addition of hLIF on the derivation of bESC-like cells, which is consistant with some results in cattle (Vejlsted et al., 2005) and pigs (Moore and Piedrahita, 1997). Pant and Keefer (2006, 2009) found that bovine ICM and its primary outgrowths do have the LIF receptor and gp130 signal transducer. It is unclear if these pathways are functional, because LIF did not help in the derivation and maintenance of bESC-like cells. A recent study (Rodriguez et al., 2007) recommended that heterospecific LIF, including human LIF and mouse LIF, should not be used to supplement the culture medium for bovine embryo or bESC-like cells because they found that embryonic development was impaired by hLIF, whereas the number of ICM cells was reduced by mLIF. Thus, it would be desirable to compare the effects between hLIF and homologous bovine LIF on derivation and maintenance of bESC-like cells because a homologous LIF supplement may play an important role in bESC-like cells culture. Indeed, Yamanaka et al. (1999) reported that bovine LIF could improve bovine embryonic development during the whole culture period of in vitro produced embryos. Unfortunately, bovine LIF has not yet been tested, and is not commercially available.
The feeder cell layer is also an important factor affecting ES cell derivation and maintenance. It serves as an attachment matrix for cells and can secret cytokines, such as LIF, that may stimulate ES cell growth and inhibit their differentiation (Li et al., 2003). MEF and homologous embryonic fibroblasts are currently the most commonly used feeder cells. Although homology exists in proteins derived from different species, the nonhomologous sequences of proteins may have different activities. Based on this consideration, effects of MEF and BEF were compared on derivation and maintenance of bESC-like cells in this study. However, BEF did not have any apparent advantage in either derivation or maintenance of bESC-like cells over MEF. This may suggest that the BEF used in this study do not secret factors whose activity differs significantly from MEF with regard to support of bESC-like cells self-renewal. Van Stekelenbery-Hamers et al. (1995) found that the efficiency of attachment and outgrowth of bovine ICM cells could be remarkably improved when bovine STO cells were used as feeder cells instead of bovine uterous epithelial cells. However, Li et al. (2004) compared three different feeders on porcine ESC cells culture, and found that porcine embryonic fibroblasts (PEF) produced the same results as MEF, and both were superior to STO cells. Another study (Tian et al., 2006) found that goat ESC-like cells could be derived on the support of MEF, but could not be derived on goat embryonic fibroblasts. Due to the long gestation period of domestic animals, systematic studies using homologous feeders at earlier and different stages of gestation should be compared. Further research is needed to compare the components secreted by different feeder cells and to test if it might be possible to use feeder-independent culture conditions for derivation bESC-like cells as was done in the mouse and human (Ludwig et al., 2006; Pease et al., 1990; Xu et al., 2001).
The method of cell passaging is crucial for the maintenance and easy handling of ES cells lines. Trypsin has been successfully used for derivation and maintenance of mouse ES cells (Evans and Kaufman, 1981). However, similar attempts have led to the loss of proliferation and induction of cell differentiation in other species including in cattle (Strelchenko, 1996), pigs (Talbot et al., 1993), monkeys (Suemori et al., 2001), and humans (Thomson et al., 1998). In our hands, after being passaged mechanically three times, bESC-like cells can be passaged 10 more times through Trypsin or TrypLE dissociation without losing pluripotency. It may be that small clumps of epiblasts that were obtained by mechanical dissociation during the first three passages, instead of total dissociation, facilitated cell–cell signaling and aided in the maintenance of pluripotency. Furthermore, when the number of bESC-like cells increased after three mechanical passages, cell–cell communication had progressed so that the single cells obtained by total dissociation through enzymatic dissociation could maintain pluripotency for a period of time. In this context, bESC-like cells like and need each other. Complete dissociation or enzymatic digestion especially in early culture leads to failure, suggesting that cell–cell signaling in bESCs derivation and maintenance is very important for self–renewal and proliferation. Another possible explanation could be that we used, for the first time, KSR instead of FBS in the bESC culture media, which may contribute to the successful passaging of bESC-like cells through enzymatic dissociation. This is supported by a recent report from our group that FBS is detrimental to mESC derivation (Kim et al., 2009).
Bovine ESC-like cells treated by TrypLE tended to survive longer than those treated by trypsin. This may be due to the fact that TrypLE is a recombinant trypsin and is less damaging than native pancreatic trypsin. The immediate differentiation of bESC-like cells once treated by Liberase Blendzyme may indicate that bESC-like cells differentiation could be induced by collagenase and/or protease because Liberase Blendzyme is a mix of collagenase and protease. The successful application of enzymatic dissociation of bESC-like cells after three mechanical passages may contribute to bESCs' application in generation of gene targeted offspring, as transfection techniques in most cases rely on single cell suspensions from which to derive gene targeted cell lines. This improvement will also facilitate the maintenance and banking of bESC lines following derivation.
From our results, it seems that our current culture conditions are adequate only to support the establishment of bESC cultures but are not able to maintain their pluripotency and continued proliferation after several passages. Bovine homologous LIF, specific chemical inhibitors of differentiation signal pathways, and/or other growth factors may need to be supplemented in the culture medium.
Footnotes
Acknowledgments
We thank Evergen Biotechnologies, Inc. for their assistance in bovine IVF and semen supplies.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.