
Abstract
Abstract
We have recently reported that maturation of the skeletal muscle is delayed in cloned calves during their first year postnatally. This delay could originate from perturbations in fetal myogenesis. The aim of this study was to evaluate the developmental characteristics of muscle in clones versus animals derived from conventional reproduction. We have characterized the anatomical and biochemical properties of the Semitendinosus muscle of clones versus controls at day 60 and day 260. We have analyzed the contractile and metabolic properties of muscle fibers by measuring the abundance of myosin heavy chain (MyHC) isoforms and activities of metabolic enzymes (LDH, PFK, COX, CS, ICDH), respectively. The spatial repartition of some components of the extracellular matrix (collagen types I, IV, VI, chondroitin-6-sulfate, decorin, and tenascin-X) was also studied. At day 60 we found lower numbers and structural organization of fibers, and a delay in the setup of the extracellular matrix. IGF-2 transcript abundance was lower in clones than in their controls. There was no difference in the expression of VEGF (a growth factor regulating vascularization and myogenesis) and its receptor. At day 260 the muscles of fetal clones have not reached the same degree of differentiation than controls as shown by their lower energy metabolisms and their MyHC pattern. These results show for the first time that disturbances in myogenesis occur early in fetal life in cloned cattle.
Introduction
During early pregnancy, monitoring fetal and placental development of clones showed that fetal growth was delayed in the peri-implantation period compared to controls (Chavatte-Palmer et al., 2006). Excessive growth is subsequently observed so that the calves tend to have higher birth weights and longer gestation periods. Some of them display abnormalities such as oversize, together with excess growth of some organs of mesodermic origin such as liver and heart, and cardiovascular abnormalities leading to poor survival rates at birth (Chavatte-Palmer et al., 2004). This pathology, referred to as large offspring syndrome (LOS) (Young et al., 1998) or abnormal offspring syndrome (Farin et al., 2006) most likely originates from fetal nutrition deregulation due to placental abnormalities (defective vascularization and reduced placentome numbers) early in gestation (Constant et al., 2006; Hill et al., 2000). Thus, bovine clones are also candidate animal models to study the long-term effects of fetal perturbations on postnatal cattle development, especially on muscle development.
During myogenesis, mononucleated myoblasts are formed from cells that originate in the somite and fuse to form multinucleated myotubes with subsequent differentiation into slow and fast fiber types. At the same time, the extracellular matrix (ECM) network differentiates. Adult muscle ECM has three separate levels of organization: the epimysium or outer muscle sheet, the perimysium binding muscle fiber bundles together, and the endomysium surrounding each individual muscle fiber. The developmental process of myogenesis can be monitored by using antibodies specific for the different fiber types including myosin heavy chain (MyHC) isoforms (Picard et al., 2002) and some ECM molecules (e.g., collagens and proteoglycans) (Carrino et al., 1999; Listrat et al., 2000), or by assaying the activities of enzymes representative of energy metabolism. After birth, little is known about the muscle development of SCNT clones. In a recent study we have shown that cloned heifers exhibit a delay in muscle maturation until the onset of puberty (Jurie et al., 2009). These differences in contractile and metabolic characteristics observed after birth could be the consequence of a delay in fetal myogenesis.
The aim of the present study was to evaluate muscle developmental characteristics in fetal clones compared to animals derived from conventional reproduction. Two important developmental stages were chosen according to Picard et al. (2002): day 60 of pregnancy, characterized by intensive cell proliferation and muscle fiber formation and day 260 of pregnancy, which corresponds to terminal differentiation of fibers, contractile and on-going metabolic differentiation. Contractile properties were evaluated through the study of MyHC isoforms by immunohistochemistry, electrophoretic separation, and Western blot. Activities of the enzymes reflecting the glycolytic and oxidative metabolic pathways were also measured. The spatial repartition of some components of the ECM (major collagen types, chondroitin-6-sulfate, decorin, and tenascin-X) was examined to characterize the setup of ECM deposition. Moreover, although the regulatory pathways that control skeletal muscle differentiation are not fully understood, data indicate that insulin-like growth factor 2 (IGF2) and vascular endothelial growth factor (VEGF) are involved in this process (Bryan et al., 2008; Hannon et al., 1992). We therefore analyzed the abundance of IGF2 transcript by qRT-PCR and the expression of VEGF and its receptors Flt-1 and Flk-1 by immunohistochemistry.
Material and Methods
Generation of cloned fetuses
Skin biopsies were collected from the ears of four cows of two different breeds [Holstein (Ho), n = 2 and Charolais (CH), n = 2] to generate fibroblast cell lines. These cryopreserved cells were used for nuclear transfer into enucleated oocytes according to Vignon et al. (1998). Briefly, recipient oocytes were matured in vitro for 22 h before enucleation by micromanipulation. Each donor cell was inserted in the perivitelline space of the enucleated oocyte and fused to the cytoplast by two electric pulses of 1.2 kV for 30 μsec. The reconstituted embryos were activated in a medium containing 10 μg/mL cycloheximide and 5 μg/mL cytochalasin B for 5 h. They were then cocultured for 7 days up to the blastocyst stage, in microdrops of B2 medium on Vero cell monolayers. By day 7, grade 1 NT blastocysts were transferred to synchronous recipients of the same breed (Ho embryos into Ho recipients and CH embryos into CHs recipients). Pregnancy was confirmed by repeated ultrasound scanning, and pregnant animals were slaughtered according to European guidelines on animal care at the experimental facilities of INRA, in order to collect the experimental fetuses.
Control animals
Ho and CH cows from the same experimental farms were inseminated with the frozen semen of their respective breeds to generate contemporaneous control fetuses. After ultrasound pregnancy confirmation, the control animals were slaughtered to collect fetuses in the same way as the clone recipients.
Recovery of muscle samples
The fetuses were recovered at day 60 (n = 4 clones and n = 4 controls for each breed) and day 260 (n = 3 Ho clones, n = 4 Ho controls). Just after slaughter, the gravid uterus was opened and the fetus carefully isolated and measured. Weights and crown–rump lengths were recorded as not differing between clones and their controls. The Semitendinosus (ST) muscle was immediately dissected. Muscle samples were snap frozen for immunohistological and biochemical experiments and stored at −80°C until analysis.
Contractile differentiation
Contractile properties of muscles depend on MyHC isoforms with specific myofibrillar ATPase activity that determines the speed of contraction of fibers. At day 60, we have thus scored the number of positive cells for the presence of fetal MyHC, after immunocytochemical staining using F1-652 antibody (1/10) (Picard et al., 2002). A combination of electrophoresis/densitometry (Picard et al., 2007) and Western blot analyses (Picard et al., 2006) was used at day 260 to examine the pattern of MyHC isoforms. Two monoclonal antibodies from Alexis Corporation were used: F113 15F4 (1/500) recognizing adult fast MyHCs IIa and IIx, and F1-652 (1/1000) specific of fetal MyHC. The specificity of antibodies on bovine skeletal muscle has been published previously (Picard et al., 2006). For electrophoretic separation of MyHC, the technique of Talmadge and Roy (1993) was used to separate the developmental isoforms, whereas the technique of Picard et al. (2007) was used to separate adult isoforms. Homogenates of ST muscles from two day 260 CH fetuses (CH 0424 and CH 0423) and an adult CH bull (adult) were included as controls in the electrophoretic and Western blot experiments.
Composition of the extracellular matrix
ECM molecules were stained by indirect immunofluorescence on 10 μm-thick sections according to the following procedure. All sections were incubated in 2% bovine serum albumin (BSA) in phosphate-buffered saline (PBS) and then for 1 h in the primary antibody solutions, washed in PBS for 15 min, and then incubated with the second antibody conjugated to dichlorotriazinylaminofluoresceine (Interchim, San Pedro, CA, USA). After washing twice for 15 min in PBS, sections were covered with glass coverslips. All pictures were captured in bright field by Sony DFW-SX900 camera coupled to a Nikon Labophot II microscope. Antibodies were diluted with 2% BSA in PBS (pH 7.2). Polyclonal rabbit antibovine type I and IV collagen antibodies (Novotec, France) were diluted to 1:40. Polyclonal rabbit antihuman type VI (Novotec) was diluted to 1:40. Monoclonal mouse antibovine decorin (DSHB, Iowa City, IA, USA), tenascin-X (generously donated by C. Lethias, IBCP, Lyon, France), chondroitin-6-sulfate (generously donated by B. Caterson, Cardiff University, Cardiff, UK) were diluted to 1:40. Secondary antibodies were diluted to 1:40. Negative controls were performed omitting primary antibodies in the same conditions than previously described. After the last washing bath, nuclei were stained with Hoechst [H33258, Sigma, St. Louis, MO, USA (B2883)] (final concentration: 2 μg/mL).
Metabolic enzyme activities
Maximum activity levels of the enzymes reflecting the glycolytic metabolic pathway [lactate dehydrogenase (LDH), EC 1.1.1.27; phosphofructokinase, (PFK), EC 2.7.1.11], mitochondrial density [isocitrate dehydrogenase (ICDH), EC 1.1.1.42; citrate synthase (CS), EC 4.1.3.7], oxidative phosphorylation cytochrome-c oxidase (COX), EC 1.9.3.1] were measured spectrophotometrically as previously described (Cassar-Malek et al., 2007; Jurie et al., 2006). Briefly, muscle samples were homogenized with a polytron in a 5% (wt/vol) solution with 10 mM Tris (pH 8.0), sucrose 0.25 M and 2 mM EDTA. One aliquot of homogenate was centrifuged at 10,000 × g for 10 min at 4°C for determination of PFK, LDH, and ICDH activities. The rest of the homogenate was freeze thawed and sonicated for determination of CS and COX activities, and thereafter centrifuged for the determination of CS activities (Jurie et al., 2006). Specific activities were expressed as units per g tissue protein, and are presented as percentages of controls.
IGF2 transcript level
IGF2 gene expression was analyzed at day 60 by quantitative RT-PCR (qPCR) as described in Bernard et al. (2009) using the primer pair Fw: TTTCTCTCTCCGCTGCTCTC and Rev: TTAGGGAGGACGGGCTGT. Briefly, total RNA was extracted from muscles using Trizol reagent (LifeTechnologies, Carlsbad, CA, USA). The RNA was then purified and treated with DNAse using the RNeasy® Mini kit (Qiagen, France). RNA integrity was checked using Lab Chip Agilent technology (Agilent Technologies, Palo Alto, CA, USA). The qPCR was realized using SYBR Green I dye. The reaction was subjected to melting curve analysis to confirm single amplified products. Real-time PCR experiment was controlled using CDH11 as internal reference gene as described in Bernard et al. (2009) (FW: GGGTCCCTGAGCTCCTTAGA and Rev: AGATTCCTCAGAACGCCAGA).
VEGF and receptor expression
Immediately after dissection, slices of ST muscle were embedded in OCT compound (Tissue-Tek) and snap frozen in liquid N2 vapor. Cryostat sections (10 μm) were mounted onto SuperFrost® microscope slides (Menzel-Gläser, Braunschweig, Germany) and stored at −80°C until used. After vacuum drying, sections were preincubated with 10% goat serum–phosphate buffer for 1 h, then with rabbit anti-VEGF A, anti-Flt-1, or Flk-1 primary antibodies (sc-152, sc-316, and sc-315, respectively, from Santa Cruz Biotechnolgy, Santa Cruz, CA, USA) overnight at 4°C. Primary antibodies preadsorbed with corresponding blocking antigenic peptides (Santa Cruz Biotechnology) were used as negative controls on serial sections. After three washes in phosphate buffer, goat antirabbit IgG-FITC conjugates (Jackson ImmunoResearch, West Grove, PA, USA) were applied for 1 h on the sections. Tissues were counterstained by F-Actin labeling by adding Rhodamin-Phalloïdin (Molecular Probes, Eugene, OR, USA) during the secondary antibody incubation. After three washes, the cell nuclei were stained with Hoechst 33342 (Sigma, St. Louis, MO, USA). Sections were covered with glass coverslips and observed on an epifluorescence microscope (Leica, DMRB, Leica Microsystem, Rueil-Malmaison, France).
Statistical analyses
All data were analyzed using the GLM procedure of SAS software (SAS Institute, Cary, NC, USA). Data from MyHC isoforms and metabolic enzyme activities were analyzed with a model that contained the fixed effect of type (clone, control). Data from IGF2 transcript level were analyzed with a model that contained the type, breed (Ho, CH) and type × breed interaction. CDH11 transcript level was introduced as a covariate in this statistical model. When significant effects were detected, differences between means were further separated by the PDIFF option of SAS.
Results
Muscle characteristics in fetal clones at day 60
At day 60, the MyHC-positive cells expressed several types of MyHC (data not shown), especially the fetal isoform, which is expressed in all myotubes at this stage (Fig. 1) and belonged to the first generation of myotubes. The scored number of myotubes was lower by 29% in the muscle of the clones compared to the control CH fetuses (p = 0.001). Within muscle fascia, the structural myotube organization was less developed in the muscle of clones compared to controls. This indicated a delay in myogenesis in fetal clones, leading to fewer primary myotubes and a lower degree of organization of the first-generation myotubes. The electrophoretic pattern of MyHC was examined in clones and their controls (Figs. 2A and 3A). Two fast and one slow developmental isoforms were detected, with no significant difference in abundance between clones and controls.
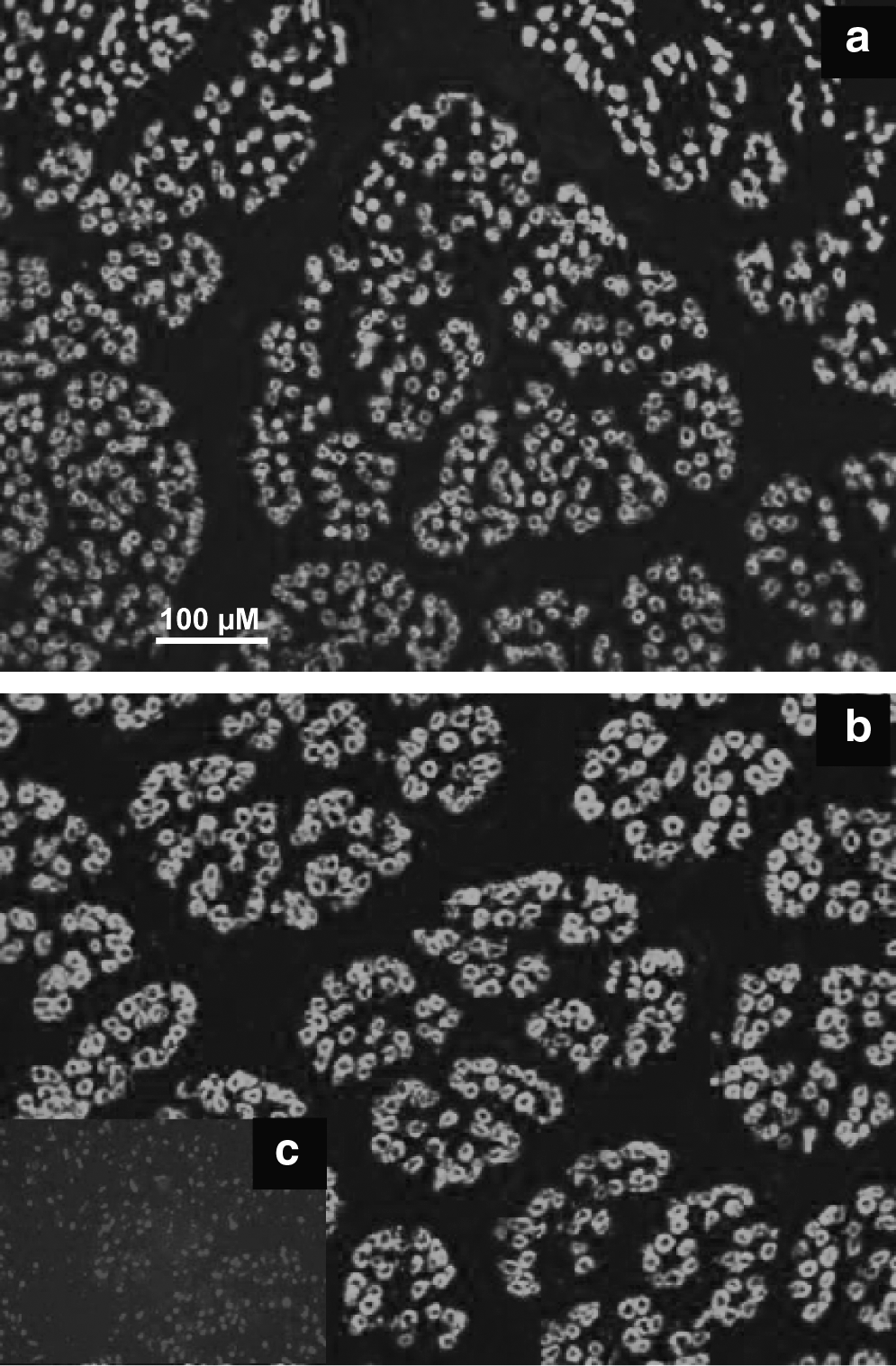
An immunocytochemical analysis of transverse muscle sections was carried out with a monoclonal antibody raised against foetal MyHC in the Semitendinosus muscle at day 60. Scale bar = 100 μm. (
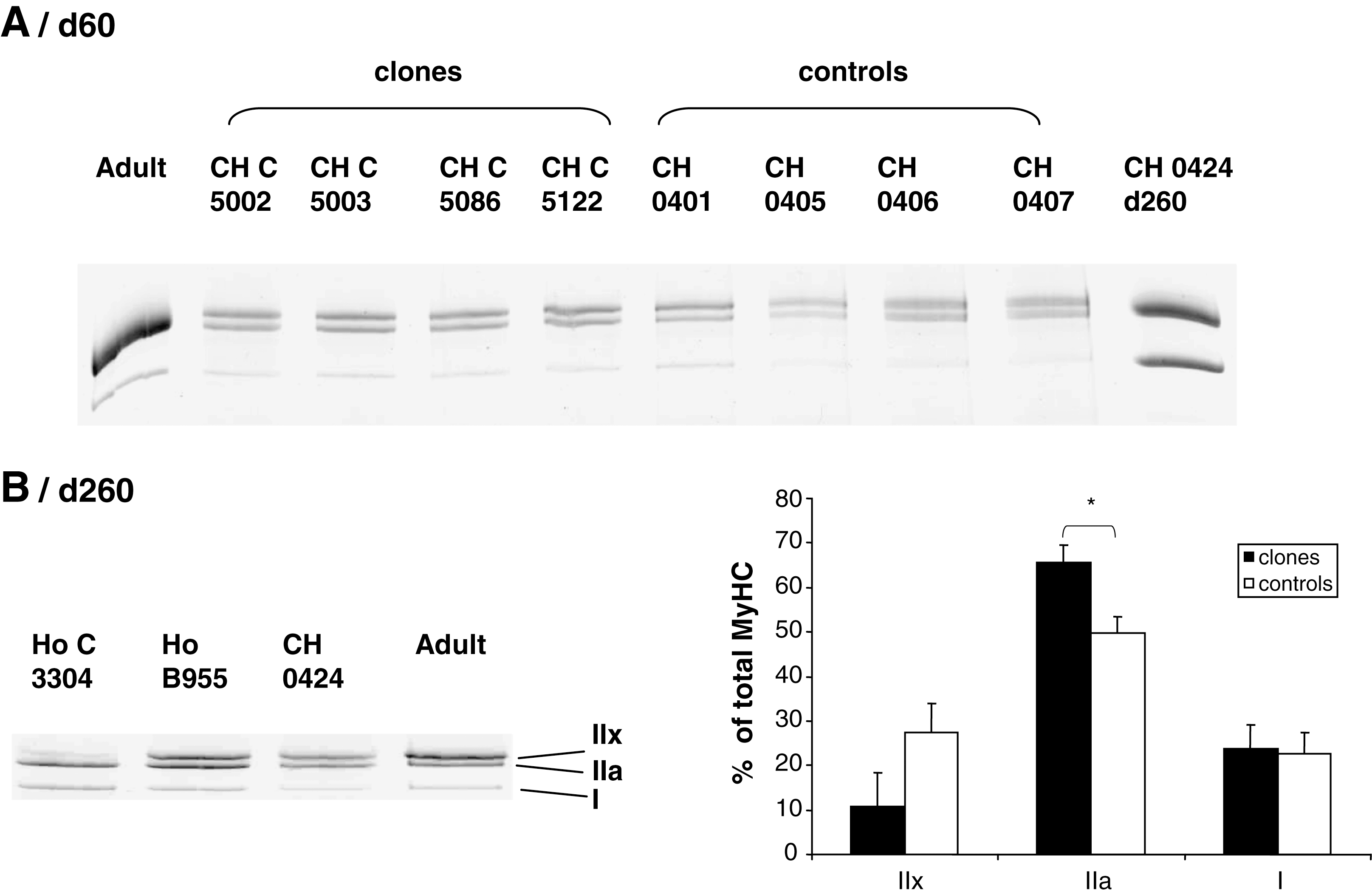
Electrophoretic pattern of MyHC isoforms in the Semitendinosus muscle at day 60 (

Expression of MyHC isoforms at day 60 and day 260 in the Semitendinosus muscle of fetal clones and their controls. The fetal (
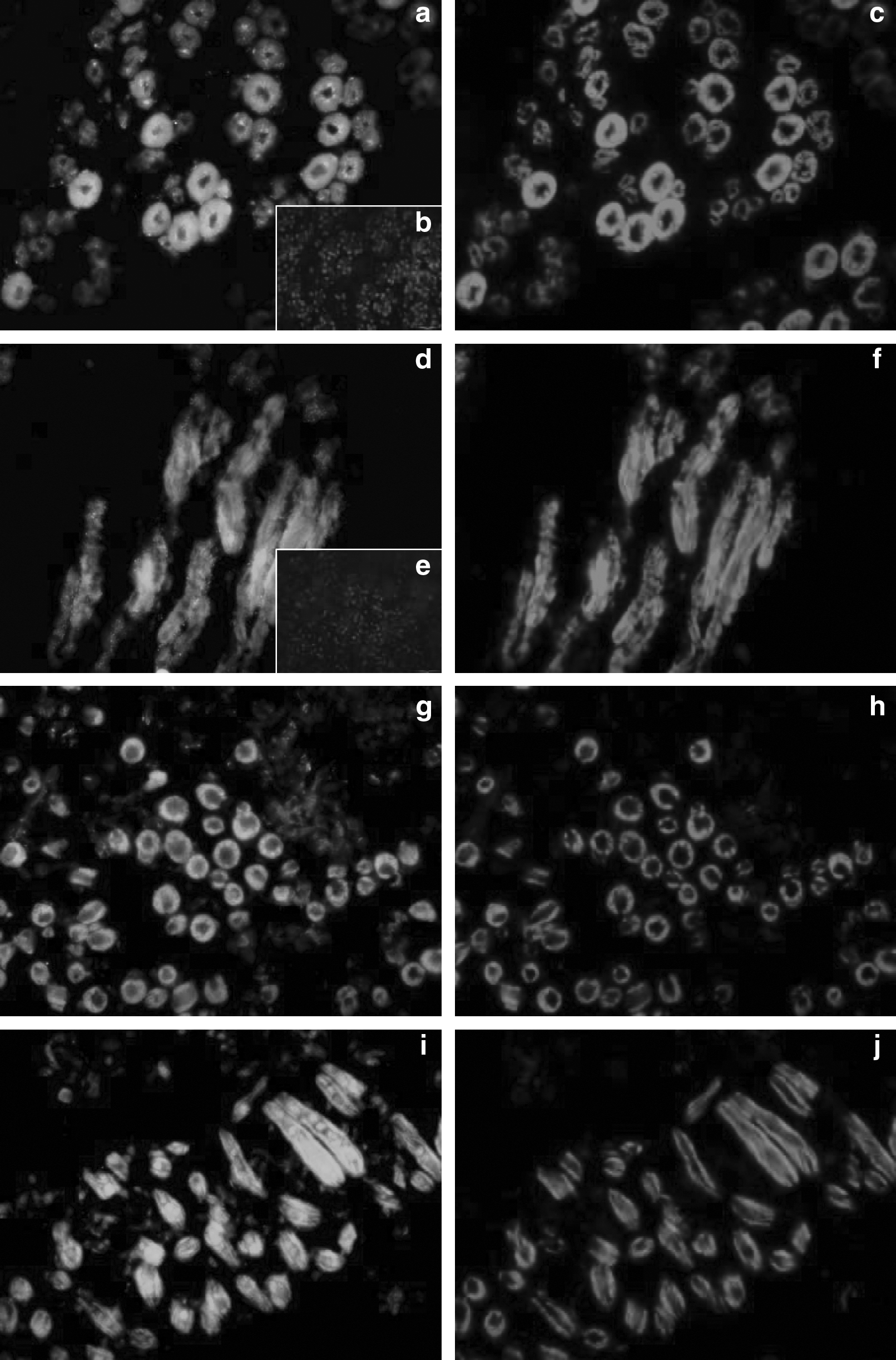
Several components of the ECM, for example, collagens (types I, IV, and VI), tenascin-X (a glycoprotein), chondroitin-6-sulfate (C6S), and decorin (a proteoglycan) were investigated by immunohistochemistry. In fetal clones (Ho or CH) as in control animals, the collagens I, IV, and VI, decorin, and C6S were detected. Type I (Fig. 4a and b) and VI collagens and decorin (Fig. 4e and f) were colocalized both in the main network of connective tissue around the muscle cell bundles (from which originate the future perimysium), and in the secondary network around and inside the muscle fiber bundles. No staining was detected in negative controls. Type I and VI collagens and decorin began to settle around the big myotubes but did not surround them completely. Type IV collagen (Fig. 4c and d) and C6S (Fig. 4g and h) were exclusively localized in the endomysium, the connective network located around the muscle cells. Whatever the staining, but mainly with type IV, we observed the presence of some small myotubes merged with the biggest ones. These myotubes were completely, partly, or even not at all individualized (Fig. 4c and d). There were less individualized small myotubes in clones than in controls. Tenascin-X (Fig. 4i and j) was colocalized with type I and VI collagens and with decorin, both in the connective tissue around the muscle cell bundles, but also within the muscle cell bundles. It was not present in the endomysium. Thus, as illustrated on Figure 4 (b, d, and h) a delay was observed in clones versus controls for the set up of the type I, IV collagens and for C6S.

Immunohistochemical analysis with antibodies directed against type I (
IGF2 transcript level was analyzed by qRT-PCR. It was 62.5% lower in the muscles of fetal clones versus their controls, irrespective of the breed (Fig. 5). Expression of VEGF and its receptors was investigated by immunohistochemistry. Both VEGF and the FLK-1 receptor were localized in the F-actin positive (myotube) cells (Fig. 6), but the Flt-1 receptor was not detected in the myotubes. No staining was observed in the negative controls (Fig. 6b and e). No differences between groups were observed, either in the location or in the intensity of staining.
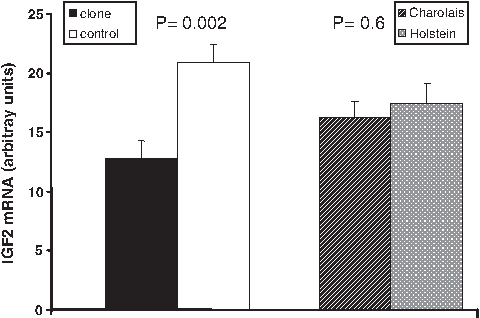
The least squares means ± SEM for IGF2 mRNA levels adjusted for CDH11 levels (the statistical model contained CDH11 levels as a covariate) in the muscle from fetal clones and their controls. Results are expressed in arbitrary units per milligram of total RNA.
Immunolocalization of VEGF (
Characteristics of the muscles of fetal clones at day 260
The activities of enzymes representative of oxidative and glycolytic metabolisms were compared in the muscles of fetal clones versus their controls (Fig. 7). The activities of two glycolytic enzymes (e.g., LDH and PFK) were lower by 26% (p = 0.015) and 49% (p < 0.001), respectively, in fetal clones compared to controls. On the other hand, the activities of two oxidative enzymes (e.g., ICDH and CS) were lower than in controls by 49% (p < 0.05) and 33% (p < 0.01), respectively, while that of COX was not significantly different in both groups. Thus, the muscles of fetal clones near term were both less oxidative and less glycolytic than those of controls. This suggested that their metabolic differentiation was delayed, leading to reduced energy metabolism.

Activities of enzymes representative of the glycolytic (LDH, PFK) and the oxidative (COX, CS, ICDH) energy metabolisms in the Semitendinosus muscle of clones compared to their controls at day 260. N = 3 fetal clones; n = 4 control fetuses. LDH, lactate dehydrogenase; PFK, phosphofructokinase; COX, cytochrome c oxidase; CS, citrate synthase; ICDH, isocitrate dehydrogenase. Results are expressed in a percentage of controls. *p < 0.05, **p < 0.01, ***p < 0.001.
Muscle contractile properties were examined through the pattern of the MyHC isoforms by electrophoresis (Figs. 2B and 3B). Although highly expressed at day 60 without differences between clones and controls (Fig. 3A), the fetal MyHC was not detectable in muscle at day 260. In contrast, MyHC adult isoforms (IIa and IIx) were only detectable at day 260 as shown by Western blot analysis (Fig. 3B). Densitometric analysis (Fig. 2B) revealed a higher abundance of the MyHC IIa in the muscle of fetal clones versus controls (p = 0.03). However, there was not significant difference abundance of MyHC IIx. This was confirmed by the immunoblotting by using an antibody recognizing the fast adult MyHCs IIa and IIx (Fig. 3B). These data indicated a delay in the appearance of the adult MyHC isoforms, clearly illustrating a delay in contractile differentiation.
Discussion
We have compared the anatomical and biological properties of muscles from fetal clones and their controls. The goal of the study was to look for possible differences in myogenesis at key developmental stages, which might explain the delay in muscle maturation during the first year of life (Jurie et al., 2009). Myogenesis takes place during several chronologically distinct phases occurring in fetal life in the bovine species: proliferation of precursor cells (myoblasts), fusion of myoblasts into differentiated multinuclear cells (myotubes), and differentiation of these cells into muscle fibers. Myotube formation occurs in at least two chronologically distinct waves, from embryonic and fetal myoblasts, respectively. They give rise to primary and secondary muscle fibers respectively. At day 60, developing muscle fibers mainly belong to the primary generation (Chaze et al., 2008) from which slow fibers originate (Picard et al., 2002), whereas nonfused myoblasts belong mainly to the secondary generation. Muscle fiber generations are set up at the end of the second trimester of gestation (day 180), a stage when the total number of fibers is fixed. Near term (at day 260), bovine muscles have acquired most of their mature properties (Picard et al., 2002). The final trimester of fetal life is above all marked by a considerable number of changes in the isoforms for contractile and metabolic proteins (adult MyHC, myosin light chains, slow or fast T-troponins, and also alpha and beta enolases) (Chaze et al., 2008). The expression of MyHCs is also modified as fetal and developmental isoforms are progressively replaced by the adult fast isoforms IIa and IIx (Picard et al., 2006). So the lower proportion of fast IIx MyHC in clones versus controls on 260 days indicated clearly a delay in the increase of expression of this isoform in clones' muscle. This is in accordance with the lower glycolytic activities of LDH and PFK, which normally increase in fast fibers during the last trimester (Cassar-Malek et al., 2007; Gagnière et al., 1999). The delay in differentiation of muscle in clones also corresponds to the lower oxidative activities of CS and ICDH. We have demonstrated that fast IIa fibers acquired their oxidative properties during the last trimester of gestation. Histochemical studies have revealed that during this period the proportion of IIa fibers, positive for Succinate dehydrogenase activity (oxidative enzyme usually studied in cellular sections), highly increased (Picard et al., 2006). Accordingly, the lower oxidative activity of the mitochondrial enzymes CS and ICDH observed in clones comparatively to controls could correspond to a lower proportion of IIa oxidative fibers illustrating a delay in the metabolic differentiation.
Muscle fibers are wrapped in ECM. Some components of the ECM, for example, members of the collagen family and the proteoglycan (PG) family, are present very early in myogenesis (Carrino et al., 1999; Listrat et al., 2000). In bovine muscle, the collagens type I, III, IV, V, XII, and XIV are present from day 60, irrespective of breed and genetic type (Listrat, data to be published). In this work we have confirmed the presence of the main ones (e.g., type I, IV, and VI) at day 60 in the muscles of fetal clones and their controls. Decorin, a small PG of the chondroitin sulphate/dermatan sulphate family which is the major PG in the muscular tissue (Nishimura et al., 2002), was detected at day 60 around muscle cells bundles and in the endomysium. This is in agreement with our unpublished data locating this PG in the major perymisium bundles but also in minor ones, and in the endomysium, irrespective of genetic type and breed. C6S was detected exclusively in the endomysium of both clones and their controls. Tenascin-X, a glycoprotein which is expressed in the muscle, skin, and nervous system (Chiquet-Ehrismann and Tucker, 2004) was highlighted in the perimysium of clones and their controls. As far as we know, tenascin-X is known to be present in the skeletal muscle, but its ontogenesis during fetal development has not yet been studied, at least not in the bovine species.
In this study, we found converging evidence that myogenesis is delayed in fetal clones early in development (day 60). Although the morphological structure of muscles appeared to be normal at the two studied stages, the number of myotubes per aera was lower, the organization of myotube fasci and the spatial distribution of some ECM components were obviously less mature in clones than in their controls at day 60. Moreover, the biochemical data indicate that near term (at day 260) the muscles of fetal clones have not reached the same stage of differentiation/maturation than in the controls, as shown by the reduced energy metabolism and the MyHC pattern.
As far as we know, this is the first demonstration that disturbances in both primary and secondary myogenesis occur in fetal life in cloned cattle. Many abnormalities have been reported in clones, including increased gestational length, perinatal mortality, fetal edema, altered body weight, altered growth of organs, hydrallantois, and abnormal placentation (Chavatte-Palmer et al., 2004; Constant et al., 2006). Lee et al. (2004) have investigated the organ development of cloned cattle fetuses at day 100 and day 150. No major malformations were observed in the brain, liver, heart and kidneys, but liver and kidney overgrowth was found. However, muscle development was not examined.
What can be the mechanisms underlying delayed myogenesis in fetal clones? Our first hypothesis was that impaired vascularization and VEGF expression and activity could be responsible for impaired myogenesis in clones. In mice, VEGF is coordinately regulated during myogenic differentiation and exerts an autocrine function regulating skeletal myogenesis (Bryan et al., 2008; Germani et al., 2003). However, no abnormal expression of VEGF and its receptor Flk-1 was detected in the muscles of clones compared to their controls. Therefore, VEGF is probably not involved in the delay in myogenesis of fetal clones. In a second hypothesis, perturbations in the ECM deposition probably impact on the time course of myogenesis. The contribution of ECM to myogenesis is well documented. The role of collagens in muscle differentiation has been reported (Gerstenfeld et al., 1984; Nusgens et al., 1986). Decorin, in addition to its interaction with fibrillar collagens and type VI collagen (Wiberg et al., 2001), also modulates the bioavailability of several growth factors that are crucial to the proliferation and differentiation of muscle cells. In particular, it regulates myostatin activity through direct interaction (Miura et al., 2006; Nishimura et al., 2007). Moreover, decorin-null myoblasts when grafted onto chick limb buds have an increased migration and are unable to differentiate (for a review, see Brandan et al., 2008). Thus, we can hypothesize that subtle change in decorin deposition, as in the case of the clones, may be a mechanism contributing to delayed myogenesis. As far as we know, it is not known whether or not tenascin-X and C6S play a role in the regulation of myogenesis. Tenascin-X is known to play a central role in the organization of the ECM of the muscle, via its interaction with various elements of this matrix such as collagens I, III, V, XII, and XIV (Lethias et al., 2006) and decorin (Elefteriou et al., 2001).
Studies have demonstrated that the abnormal development of animal clones is the consequence of epigenetic modifications during the early development of cloned embryos, primarily DNA methylation, and histone modification (Kang et al., 2001; Niemann and Wrenzycki, 2000, Wee et al., 2006), two processes essential for the regulation of gene expression. Inefficient epigenetic reprogramming upon SCNT could result in the inappropriate expression of key genes crucial for developmental processes, such as fetal growth and development (Farin et al., 2006; Lin et al., 2008). Indeed, aberrant expression of several genes has been found in cloned animals (Wrenzycki et al., 2004) including imprinted genes (Yang et al., 2005), X-linked genes (Xue et al., 2002), and apoptosis-related genes (Amarnath et al., 2007). However, scarce information is available on candidate genes directly related to skeletal muscle development in fetuses derived from reproductive biotechnologies. Crosier et al. (2002) reported that in vitro production of bovine embryos resulted in fetuses with altered development of muscle fibers and a decreased expression of mRNA for myostatin was observed in muscle of fetuses in the in vitro group compared to in vivo controls.
Aberrant fetal tissue expression of several components of the IGF system, an important regulator of both placental and fetal growth, has been reported during the gestation (Long and Cai, 2007) and postnatal life (Li et al., 2007; Yang et al., 2005; Zhang et al., 2009) of cloned cattle. In SCNT-derived cloned day 60 whole cattle fetuses, IGF2 is the most abundant transcript together with fetuin (Taniguchi et al., 2001). Expressions of IGF2, H19, and IGF2R are greater in tissues from deceased newborn clones (Yang et al., 2005). Plasma levels of IGF2, however, are lower in live newborn clones compared to controls (Chavatte-Palmer et al., 2002), although this difference does not persist thereafter. Epigenetic abnormalities of the IGF2/H19 imprinting region have been correlated with LOS in sheep (Young et al., 2003) and in cattle derived from SCNT (Curchoe et al., 2009). In the muscles of clones that survived adulthood, expressions of these genes are normal, except for the expression of IGF2, which is highly variable. IGF2 is a fetal mitogen and promotes mesoderm formation (Morali et al., 2000). It is a promyogenic factor (Hannon et al., 1992; Prelle et al., 2000; Stewart et al., 1996) and an autocrine factor for differentiating muscle cells (Stewart and Rotwein, 1996). It is highly expressed in fetal muscle (Lee et al., 1990). In our study, expression of IGF2 gene was lower at day 60 in the muscles of fetal clones, which may account for delayed myogenesis. In cattle fetuses, the muscle expression of the IGF2 gene exhibits a chronological and spatial pattern. The expression of all but one IGF2 transcripts decreases throughout gestation while myogenesis is proceeding (Listrat et al., 1999a). There is a chronological relationship between the presence of IGF2 transcripts, IGF receptors and differentiation in fetal muscles, suggesting that local production of IGF2 could regulate fetal myogenesis (Listrat et al., 1999b). Moreover, IGF2 transcripts, primarily localized in developing fiber bundles, are shifted away from muscle after day 160, and thereafter begin to localize within the connective tissue (Listrat et al., 1994). Alteration of temporal IGF2 expression pattern in the muscles of clones may therefore contribute to delayed myogenesis. However, the aberrant expression of other genes may not be excluded. Indeed, altered development of muscle fibers was previously observed in fetuses produced in vitro (Crosier et al., 2002). In contrast to the data obtained here with clones, the number of fibers and their metabolism were increased in the treated animals, and the authors pointed out myostatin as a candidate gene whose expression may contribute to the observed changes in muscle development. Both proteomic and transcriptomic experiments are in progress to identify molecular pathways that may underlie the delayed myogenesis of fetal clones.
Footnotes
Acknowledgments
The authors thank D. LeBourhis, C Richard, G. Gentes, C. Barboiron, A. Delavaud, D. Chadeyron, and C. Nore for their skilled technical assistance, and the staff of the URH experimental slaughterhouse and the experimental farms at UCEA-Bressonvilliers and Domaine de Gallé-Bourges for animal care. This study was funded by a grant of GenAnimal (ANR-06-GANI-006_MUSCLON).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.