
Abstract
Abstract
Previous studies have demonstrated that increased histone acetylation in donor cells or cloned embryos, by applying a histone deacetylase inhibitor (HDACi) such as trichostatin A (TSA), significantly enhances their developmental competence. However, its effect may vary with the type of HDACi and the target species, with some research showing nonsignificant or detrimental effects of TSA on in vitro and in vivo development of embryos. In this study, we show that sodium salt of butyric acid, a short-chain fatty acid produced naturally in the body by bacterial degradation of dietary fibers in the colon and rectum, increases histone acetylation in pig fibroblast and embryos at a concentration of 1.0 and 5.0 mM, respectively. However, treatment of donor cells with NaBu did not affect the rate of blastocyst formation or embryo quality in terms of histone acetylation and total nuclei per blastocyst (p > 0.05). On the contrary, treatment of cloned pig embryos with NaBu for 4 h significantly enhanced (p < 0.01) the rate of blastocyst formation (18.3 ± 2.1 vs. 11.2 ± 3.0%), although the total nuclei number per blastocyst did not differ. More importantly, blastocysts generated from NaBu-treated cloned embryos had increased levels of histone acetylation that was comparable to those of in vitro fertilized (IVF) embryos (36.7 ± 3.6 vs. 45.9 ± 2.5). In conclusion, our data suggest that histone hyperacetylation by NaBu treatment of cloned embryos, but not donor cell, enhances their in vitro development up to blastocyst stage.
Introduction
Acetylation of amino acid residues at the N-termini of the nuclear histone protein tail is an epigenetic regulatory mechanism for controlling gene expression. It may also regulate DNA replication, histone deposition, and the DNA repair mechanism by recruiting proteins having an acetyl lysine binding module, the bromodomain (Rice and Allis, 2001). Acetylation of specific amino acid residues at different tails of various histone proteins form an histone code that is tightly regulated, and show a specific temporal and spatial pattern during embryonic development (Shi and Wu, 2009; VerMilyea et al., 2009). However, cloned embryos produced by SCNT occasionally show epigenetic error in histone acetylation due to inheritance of epigenetic marks from somatic chromatin (Wee et al., 2006). Consequently, chemical modification of histone acetylation by histone deacetylase inhibitors (HDACi) has emerged as a potential solution for improving cloning efficiency.
Several studies have shown that treatment of donor cells with HDACi such as Trichostatin A (TSA) can improve the in vitro development of cloned cattle embryos by partial erasure of preexisting epigenetic marks of donor cells (Ding et al., 2008; Enright et al., 2003; Wee et al., 2007). TSA treatment also altered the expression of imprinted genes in cloned embryos by reducing the expression of HDAC and DNA methyltransferase without affecting the expression of histone acetyltransferase (HAT) in donor cells (Wee et al., 2007). However, the beneficial effect of treating donor cells with HDACi on full-term development of cloned embryos has never been reported. Moreover, the effect of HDACi treatment to donor cells has not been investigated in other species such as the pig.
Studies in the mouse show that, instead of treating donor cell, TSA treatment of cloned embryos after nuclear injection was effective in markedly improving the cloning efficiency (Kishigami et al., 2006; Maalouf et al., 2009; Rybouchkin et al., 2006) both in terms of in vitro development to blastocyst stage and in vivo development to full term. However, variable results have been reported on the effect of TSA in other species, and some groups have even reported its detrimental effects on in vitro and in vivo embryonic development (Iager et al., 2008; Li et al., 2008; Meng et al., 2009; Shi et al., 2008; Srirattana et al., 2008; Svensson et al., 1998; Tsuji et al., 2009; Wu et al., 2008; Zhang et al., 2007; Zhao et al., 2009). More recently, Zhao et al. (2009) reported that scriptaid, an HDACi less toxic than TSA, improved the cloning efficiency of inbred pigs that were difficult to clone by conventional SCNT. Similar results were also reported for scriptaid and other HDACi in the mouse by Van Thuan et al. (2009), who further found that improvement in cloning efficiency by HDACi was likely due to their ability to stimulate de novo synthesis of nascent mRNA in treated embryos. Thus, efficacy of HDACi on nuclear reprogramming of cloned embryos seems to vary with the type of HDACi and the target animal species.
Butyric acid is a short-chain fatty acid produced naturally in the body by bacterial degradation of dietary fibers in the colon and rectum (Bugaut and Bentejac 1993; Russo et al., 1997). Sodium salt of butyric acid (sodium butyrate or NaBu) has a potent HDACi activity, and has been shown to have differentiating and antiproliferative effect on tumor cells by altering the expression of several proto-oncogenes (Abramova et al., 2006; Cuisset et al., 1998; Drottar et al., 2006; Russo et al., 1997). Through inhibition of HDAC, it also induce chromatin rearrangement, release of calcium from intracellular stores, and altered gene expression in several somatic cell lines (Goicoa et al., 2006; Ou et al., 1997). Zakhartchenko and coworkers were the first to show that treatment of donor cells with NaBu markedly improves the in vitro development of cloned embryos in cattle (Shi et al., 2003) and rabbits (Yang et al., 2007). They also found that NaBu was more potent than TSA in improving the cloning efficiency of cloned embryos (Shi et al., 2003). NaBu has also been shown to induce hyperacetylation of histone protein in pig fibroblasts (Mohana Kumar et al., 2007), although utility of NaBu-treated cells for SCNT were not investigated. It is also not known, if postactivation treatment of cloned embryos with NaBu would be more beneficial than treatment of donor cell before the SCNT process.
This study was therefore designed to investigate the in vitro development and histone acetylation of cloned pig embryos produced by SCNT of NaBu-treated donor cells. Because NaBu had been shown to be more potent than TSA (Shi et al., 2003), this study also investigated whether postactivation treatment of cloned embryos with NaBu would be beneficial for improving cloning efficiency.
Materials and Methods
All chemicals were obtained from Sigma–Aldrich Co. (St. Louis, MO, USA) unless otherwise specifically indicated. Each experiment consisted of at least three replicates, and in each of the replication oocytes were from the same collection of abattoir-derived ovaries collected on the same day.
Oocyte retrieval and in vitro maturation (IVM)
Retrieval and IVM of pig oocytes, from abattoir-derived ovaries, were performed essentially as we described earlier (Gupta et al., 2008a). Briefly, cumulus–oophorus complexes were aspirated from medium-sized follicles (3–6-mm diameter) and matured in groups of 50 in 500 μL of Tissue Culture Medium 199 with Earle's salts (TCM-199; Gibco BRL, Grand Island, NY, USA) supplemented with 25 mM NaHCO3, 10% (v/v) pig follicular fluid, 0.57 mM cysteine, 0.22 μg/mL sodium pyruvate, 25 μg/mL gentamicin sulfate, 0.5 μg/mL FSH (Folltropin V; Vetrepharm, Canada), 1 μg/mL estradiol-17β, and 10 ng/mL epidermal growth factor under mineral oil at 39°C in a humidified atmosphere of 5% CO2 in air for 40–42 h.
Somatic cell nuclear transfer (SCNT)
Nuclear transfer was performed essentially as we described earlier (Uhm et al., 2009). Briefly, in vitro matured oocytes were denuded of cumulus cells in TL-HEPES medium supplemented with 0.1% hyaluronidase and enucleated by aspirating the first polar body and adjacent cytoplasm (approximately 30% of ooplasm) using a beveled borosilicate pipette (25 μm internal diameter). Successful enucleation was confirmed by UV-assisted visualization of the fluorescent metaphase plate in the aspirated cytoplasm contained within the enucleation pipette. Enucleated oocytes were subsequently reconstructed by inserting a small-sized (∼15 μm in diameter), smooth-bordered fibroblast cell into the perivitelline space of each enucleated oocyte using the same pipette used for enucleation. Donor cells for SCNT were prepared essentially the same as we described earlier (Gupta et al., 2007c). Membrane fusion of donor cell with cytoplast was induced with single DC pulse of 2.1 kV/cm for 30 μsec delivered by a BTX Electro Cell Manipulator 2001 (BTX, San Diego, CA, USA). The same pulse was also utilized to simultaneously induce activation of reconstructed embryos.
Parthenogenetic activation (PA) of oocytes
Presumptive diploid parthenotes were produced essentially as we described earlier (Gupta et al., 2007a). Briefly, denuded oocytes were placed in activation medium (0.3 M mannitol, 0.1 mM MgSO4, and 1.0 mM CaCl2) and electro-activated by a single DC pulse of 2.1 kV/cm for 30 μsec. Following activation, activated oocytes were cultured in North Carolina State University 23 (NCSU23) medium supplemented with 0.4% essentially fatty acid free bovine serum albumin (BSA) and 7.5 μg/mL cytochalasin B for 4 h.
In vitro culture (IVC) of embryos
Reconstructed and activated embryos were cultured in NCSU23 medium supplemented with 0.4% BSA for 5 days and in NCSU23 medium supplemented with 10% (v/v) FBS for subsequent 2 days as we described earlier (Gupta et al., 2008b). The rates of cleavage and blastocyst formation were recorded on day 2 and day 7 of IVC, respectively.
Fluorescent staining of blastocysts for assessment of nuclear number
Nuclear staining for counting the total nuclei number of blastocysts was performed essentially as we described earlier (Uhm et al., 2007). Briefly, day 7 blastocysts were fixed for 5 min in a fixative solution containing 2% formalin and 0.25% gluteraldehyde, mounted on clean glass slides, and stained with a glycerol-based Hoechst 33342 (12.5 μg/mL) staining solution for 10 min. Total number of stained nuclei, which appeared blue when visualized under UV illumination of an epifluorescent microscope (Olympus, Tokyo, Japan) fitted with blue filter (excitation: 330–385 nm; emission: 420 nm; dichromatic: 400 nm), were then counted for individual blastocysts, and digital images (Nikon Coolpix 990; Nikon Corporation, Tokyo, Japan) were taken.
Assays for cell cycle, proliferation, and viability
Analysis for distribution of cells into different cell cycle stages was performed by flow cytometry as described earlier (Shi et al., 2003). Briefly, confluent cultured cells were dissociated into single-cell suspension by trypsinization, fixed in 70% ice-cold ethanol for 1 h, and then incubated with staining solution containing 50 μg/mL propidium iodide, 100 μg/mL ribonuclease A, and 1% Triton X-100 for 20 min at room temperature. Cell cycle distribution was determined by flow cytometry (FacsCalibur, Becton Dickinson, Mountain View, CA, USA). Ten thousand events were recorded for each sample and all analyzed events were gated to remove debris and aggregates. Cell Quest software (Becton Dickinson, Bedford, MA, USA) was used for data analysis. To estimate the rate of cell proliferation, cells were plated in 24-well plates at an initial seeding density of 4 × 104 cells/mL in triplicate, and were counted for total number of cells every 24 h for 5 days using Hemocytometer. Viability of cells was evaluated based on the esterase enzyme activity and plasma membrane integrity using FDA (3′ 6′diacetyl fluorescein) staining as described earlier (Gupta et al., 2007b). Briefly, cells were washed in phosphate-buffered saline (PBS) for 1 min, followed by incubation with 2.5 μg/mL FDA stain for 1 min. Stained cells were then washed in PBS to remove the traces of the dye and observed under UV illumination of an epifluorescent microscope fitted with an FITC filter set (excitation: 460–490 nm; emission; 515–550 nm; dichromatic: 505 nm). Live cells emitted green fluorescence, whereas dead ones were nonfluorescent. Viability was calculated as the number of green cells/total number of cells × 100. All experiments were repeated three times.
Immunocytochemistry for assessment of acetylation at histone protein
Immunocytochemistry for acetylation of nuclear histone protein was performed as described earlier (Rybouchkin et al., 2006; Yeo et al., 2005) with partial modifications. Briefly, embryos were treated with 0.25% pronase enzyme to remove zona pellucida and fixed in 4% Paraformaldehyde for 30 min at 4°C. Fixed embryos were then permeabilized in 0.5% Triton X-100, treated with 1% BSA to block nonspecific binding of antibodies and incubated with rabbit antibody against acetylated Lysine at the ninth position of histone 3 protein (Ac-H3K9) (1:100, Upstate Biotechnology, Lake Placid, NY, USA) for 1 h each. Bound primary antibodies were localized with FITC-conjugated antirabbit IgG second antibody (1:250, Molecular Probes, Leiden, The Netherlands). The embryos were then stained with 300 nM 4′, 6′-diamidino-2-phenylindole (DAPI) and observed using a fluorescent microscopy with ApoTome (Axiovert 200, Carl Zeiss, Germany). Images were captured digitally using standard DAPI and FITC filter sets and processed using software (AxioVision v4.4; Carl Zeiss). Experiments were repeated three times with ∼10–20 embryos in each replication. In case of somatic cells, immunocytochemistry was performed essentially as described earlier except that they were cultured on Poly-L Lysine-coated coverslips placed on 35-mm tissue culture dishes (Falcon BD, Lincoln Park, NJ, USA) before the start of the experiment for this purpose. To compare the signal intensities of Ac-H3K9, the brightness of DAPI signal was first measured as a reference in the same focal plane as other embryonic nuclei. Thereafter, average pixel intensity value for a fixed pixel size (5 × 5) area was measured in five different random regions of cytoplasm, and the total average was subtracted from the same total average measured in randomly selected regions of chromosomes (Green channel). This average chromatin intensity staining was used for comparison of acetylation level. In vitro fertilized (IVF) embryos were used as control for comparison of Ac-H3K9 in cloned embryos and were produced essentially as we described earlier (Gupta et al., 2009).
Statistical analyses
Statistical analyses were performed using SAS software (Statistical Analysis System Inc., Cary, NC, USA) for the chi-square test or analysis of variance (ANOVA) as appropriate. The percentage data were subjected to arc sine transformation before statistical analyses. Data are presented as mean ± SEM. Differences at p ≤ 0.05 were considered significant.
Results
Effect of NaBu on acetylation
In the first set of experiments we evaluated the potency of NaBu in causing histone hyperacetylation. Pig fibroblasts were treated for 24 h and oocytes were treated for 4 h postactivation with different concentrations of NaBu (0.0, 1.0, 2.5, 5.0, and 10.0 mM) and evaluated for histone acetylation at H3K9 (Ac-H3K9) by immnocytochemistry. H3K9 residue was specifically chosen for analysis because this residue is nonacetylated in metaphase nuclei, and hence, Ac-H3K9 of metaphase nuclei upon treatment with NaBu directly indicates the outcome of HDAC inhibition by NaBu (Gupta et al., unpublished data; Yeo et al., 2005). We observed that NaBu could increase histone acetylation in pig fibroblasts when used at a concentration of 1.0 mM or more (Fig. 1A). However, in case of oocytes, it was effective at a concentration of 5.0 mM or higher (Fig. 2), probably due to their larger cell size and smaller surface-to-volume ratio than that of somatic cells or due to differences in their physiochemical properties. Although acetylation of H3K9 changes with cell cycle stages, the difference in concentration of NaBu required for causing histone hyperacetylation in cells and oocytes was not due to differences in the cell cycle stage because the same concentration of NaBu was required for causing histone hyperacetylation in both metaphasic and interphasic somatic nuclei (data not shown). Moreover, as shown in Figure 1B, treatment of pig fibroblasts with NaBu for 24 h had no significant effect on distribution of cells into different cell cycle stages when used at a concentration as high as 5.0 mM. Thus, for all further experiments, NaBu was used at a concentration of 1.0 mM or 5.0 mM for pig fibroblasts and embryos, respectively.
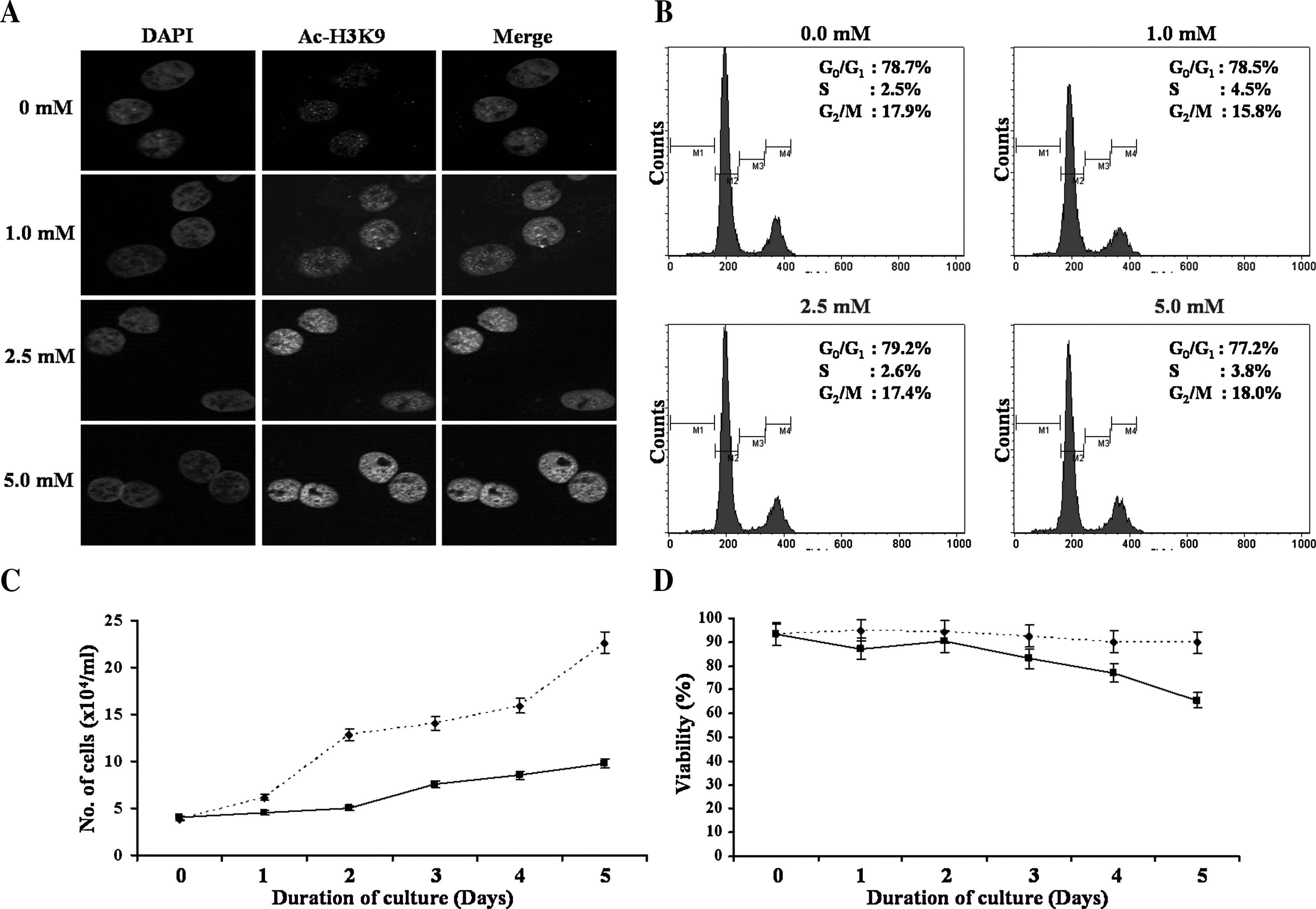
Effect of sodium butyrate (NaBu) on histone acetylation (

Effect of sodium butyrate (NaBu) on acetylation at the ninth Lysine residue of histone 3 nuclear protein (Ac-H3K9) in parthenogenetic pig embryos treated with different concentration of sodium butyrate for 4 h postactivation.
Effect of NaBu on somatic cells
Next we evaluated the effect of NaBu on proliferation and viability of pig fibroblasts and their subsequent in vitro embryonic development following SCNT. Pig fibroblasts were treated with 1.0 mM NaBu for 5 days, and were evaluated for rate of proliferation and viability. We observed that NaBu reduced both the rate of proliferation (Fig. 1C) and the viability (Fig. 1D) of pig fibroblasts in a time-dependent manner. Moreover, when used at a higher concentration or for a longer period of time, plasma membranes of pig fibroblasts became fragile and were not suitable for electrofusion with enucleated oocytes during SCNT (data not shown). Thus, cells were treated with 1.0 mM NaBu for 24 h to evaluate its effect on embryonic development following SCNT. As shown in Table 1, rates of cleavage (72.5 ± 1.9 vs. 77.4 ± 4.3%) and blastocyst formation (12.4 ± 2.2 vs. 14.0 ± 1.9%) did not differ between control and treatment groups (p > 0.05). There was also no significant effect of NaBu treatment on total nuclei number (37.9 ± 3.4 vs. 33.0 ± 3.5) or Ac-H3K9 level (16.9 ± 4.1 vs. 12.3 ± 4.1) in cloned blastocysts compared to control (Fig. 3). Interestingly, we also observed that ∼10% of cloned blastocysts showed Ac-H3K9 level comparable with those of IVF blastocysts.

Effect of sodium butyrate (NaBu) on acetylation at the ninth Lysine residue of histone 3 nuclear protein (Ac-H3K9) in cloned pig blastocysts. (
Four replicates were performed.
Values in parentheses indicate the number of embryos.
Values with same superscripts within column did not differ significantly (p > 0.05).
Effect of NaBu on embryo
Finally we evaluated the effect of treating cloned embryos with NaBu on their in vitro development and embryo quality. Because previous studies have shown a beneficial effect of HDACi when cloned embryos were treated for 4–24 h in different species (Kishigami et al., 2006; Shi et al., 2008; Yamanaka et al., 2009) and we observed a negative effect of NaBu on the viability of somatic cells, we first evaluated the time-dependent effect of NaBu on embryonic development. For this purpose, PA embryos were preferred over SCNT embryos, partly because of technical ease and partly because, unlike SCNT embryos, PA embryos were not expected to have a beneficial effect due to HDAC inhibition, and hence, formed a suitable model system for evaluating the possible toxicity of NaBu. Presumptive diploid PA embryos were produced essentially by same electroactivation protocol that was used for producing SCNT embryos, and treated with NaBu for 0, 4, 10, or 24 h to evaluate their in vitro development up to blastocyst stage. As shown in Table 2, NaBu did not have a significant adverse effect on rates of cleavage (72.8 ± 1.1 vs. 74.4 ± 0.7%) and blastocyst (29.3 ± 2.0 vs. 32.9 ± 2.5%) or total nuclei number per blastocyst (69.2 ± 11.7 vs. 68.9 ± 5.9) of PA embryos when treated for 4 h. However, NaBu reduced the rate of blastocyst formation and total nuclei number per blastocyst when it was used for 10 h or longer duration (p < 0.05). Thus, a 4-h treatment period was chosen for treating cloned embryos. Karyoplast–cytoplast couplets produced by SCNT of nontreated donor cells were randomly split into control and treatment groups, and were evaluated for their in vitro development and embryo quality after treatment with NaBu for 4 h postactivation. As shown in Table 3, NaBu-treatment of fused couplets had no significant effect on rate of cleavage (p > 0.05). However, NaBu treatment significantly improved the rate of blastocyst formation (11.2 ± 3.0 vs. 18.3 ± 2.1; p < 0.01), although total nuclei number per blastocyst did not differ significantly (p > 0.05). The blastocysts produced from NaBu-treated cloned embryos also had an increased level of Ac-H3K9 (36.7 ± 3.6) that was comparable to those of IVF (45.9 ± 2.5) embryos (Fig. 3).
hpa, hours postactivation. Five replicates were performed.
Values in parentheses indicate the number of embryos.
Values with different superscripts (a,b) within column differ significantly (p < 0.05).
Five replicates were performed.
Values in parentheses indicate the number of embryos.
Values with different superscripts (a,b) within column differ significantly (p < 0.05).
Discussion
There is a common agreement in the literature on the effect of HDACi in improving cloning efficacy. However, a discrepancy exists on its use prior to SCNT or after SCNT. Although studies in cattle showed a beneficial effect of HDACi when donor cells were treated prior to SCNT (Ding et al., 2008; Enright et al., 2003; Iager et al., 2008; Wee et al., 2007), studies in mice showed the necessity of HDACi treatment after SCNT (Kishigami et al., 2006; Maalouf et al., 2009; Rybouchkin et al., 2006; Van Thuan et al., 2009). Moreover, the efficacy of HDACi in improving cloning efficiency varied with the species and the type of HDACi used (Van Thuan et al., 2009; Zhao et al., 2009). TSA, the most commonly used HDACi, showed a variable effect on improving cloning efficiency in different species, and some groups even reported its detrimental effect on in vivo and in vitro embryonic development (Iager et al., 2008; Li et al., 2008; Meng et al., 2009; Shi et al., 2008; Svensson et al., 1998; Tsuji et al., 2009; Wu et al., 2008; Zhang et al., 2007; Zhao et al., 2009). We show that NaBu, an HDACi shown to be more potent than TSA in improving cloning efficiency of cattle (Shi et al., 2003), significantly improves the in vitro development of cloned pig embryos when treated for 4 h postactivation, but was inefficient when somatic nuclei were treated with it prior to SCNT.
We observed that, consistent with published reports (Mohana Kumar et al., 2007), NaBu can cause histone hyperacetylation in pig fibroblasts when used at a concentration as low as 1.0 mM (Fig. 1A), but resulted in a gradual reduction of cell proliferation (Fig. 1C) and viability (Fig. 1D) when used for a longer duration. However, contrary to our expectation, there was no beneficial effect of NaBu-treated donor cells on improving subsequent in vitro development of cloned pig embryos (Table 1). This was in sharp contrast to those reported by Shi et al. (2003). Although this might suggest occurrence of species difference (Kang et al., 2001) in responding to an epigenetic modifier or NaBu, it may also be due to the difference in donor cell type. Shi et al. (2003) used an immortalized mammary epithelial cell line (MECL) for SCNT that did not show histone hyperacetylation upon treatment with NaBu at a concentration effective for pig fibroblasts (Mohana Kumar et al., 2007). Thus, the effect of NaBu on improving the cloning efficiency of cloned cattle embryos produced by SCNT of MECL may have been due to effects other than histone hyperacetylation. Nevertheless, the possibility of species difference cannot be completely ruled out because the beneficial effect of NaBu-treated donor cells has also been observed in rabbits (Yang et al., 2007). Moreover, a similar beneficial effect of TSA-treated donor cells has been reported for cattle (Enright et al., 2003; Wee et al., 2007). To rule out the possibility that NaBu-induced changes in distribution of cells into different cell cycle stages (Oback and Wells 2002) might have nullified the potential beneficial effect of NaBu on improving cloning efficiency, we evaluated the cell cycle distribution of NaBu-treated pig fibroblasts. NaBu is known to suppress cell proliferation and induce G1/S and/or G2/M block of the cell cycle in various tumor cell lines (Abramova et al., 2006; Russo et al., 1997) but not in immortalized MECL of cattle (Shi et al., 2003). We found that treatment of pig fibroblast with NaBu for 24 h did not affect the distribution of cells into various cell cycle stages (Fig. 1B). This clearly indicates that lack of a beneficial effect from NaBu treatment of donor cells was not associated with changes in cell cycle distribution.
Next, we evaluated the effect of treating cloned pig embryos with NaBu after SCNT. The effective concentration of NaBu for causing histone hyperacetylation (Fig. 2) without possible toxicity (Table 2) was determined in control PA embryos. We found that treatment of cloned pig embryos with NaBu significantly (p < 0.01) increased their in vitro development up to the blastocyst stage (Table 3). However, similar to that reported by others, the effect of HDACi on improving the in vitro development of cloned pig embryos (22–35 vs. 17%, Yamanaka et al., 2009; 21 vs. 9%, Zhao et al., 2009) was not as pronounced as has been reported for the mouse (∼80% vs. 20%) (Kishigami et al., 2006; Ryobuskin et al., 2006; Van Thuan et al., 2009). Some groups have even reported little or no significant effect of treating cloned embryos with HDACi on rate of blastocyst formation (Ding et al., 2008; Iager et al., 2008; Meng et al., 2009; Shi et al., 2008; Wu et al., 2008). Interestingly, however, Li et al. (2008) found a dramatic increase in the rate of blastocyst formation (∼80 vs. ∼54%) following TSA treatment of cloned pig embryos produced by hand-made cloning (HMC). The TSA-treated embryos also lead to live births following embryo transfer, although none survived beyond 2 months. To rule out the possibility that differences in our results with those of Li et al. (2008) were not due to the type of HDACi, in a separate experiment we evaluated the in vitro development of cloned pig embryos treated with TSA similar to published reports (Li et al., 2008; Yamanaka et al., 2009; Zhang et al., 2007). We found that TSA indeed increased (Supplementary Table 1; Supplementary Fig. 1) the in vitro development of cloned pig embryos by more than twofold, as reported by others (Li et al., 2008; Yamanaka et al., 2009; Zhang et al., 2007), but again, the effect was not as profound as has been reported for the mouse (Kishigami et al., 2006; Rybouchkin et al., 2006; Van Thuan et al., 2009). Thus, apparently, TSA is more effective than NaBu in improving the in vitro development of cloned embryos but the effect of both HDACi is less profound in pig embryos than that reported in the mouse. The lower efficacy of NaBu in improving the in vitro development of cloned pig embryos may be because maintaining histone hyperacetylation for 4 h by NaBu treatment was not as adequate as longer periods of treatment that have been used for other HDACi such as TSA or scriptaid (3–6 h for rabbit, Shi et al., 2008; 10 h for the mouse, Kishigami et al., 2006, Ryoboskin et al., 2006, Van Thuan et al., 2009; 24 h for pigs and cattle, Iager et al., 2009; Yamanaka et al., 2009; Zhang et al., 2007; Zhao et al., 2009). Yamanaka et al. (2009) found that TSA was not effective in improving the in vitro development of minipig embryos when they were treated for 10 h or less. However, it was not possible to test the effect of NaBu for 24 h, as it resulted in a decreased rate of in vitro development in PA embryos (Table 2), possibly due to toxicity (Mohana Kumar et al., 2007; Shi et al., 2003).
We also found that blastocysts generated from NaBu-treated cloned embryos had increased histone acetylation at a level similar to that observed with IVF blastocysts (Fig. 3). Previous studies have suggested that the level of histone acetylation during early embryonic development affects their subsequent development (Iager et al., 2008; Li et al., 2008; Van Thuan et al., 2009; Yamanaka et al., 2009; Zhao et al., 2009), and hence, histone hyperacetylated cloned embryos may have higher development competence than that of nontreated embryos (Bourc'his et al., 2001; Iager et al., 2008). However, further experiment is required to test if NaBu-treated embryos have a higher development to full term, as Rybouchkin et al. (2006) found that the increased rate of blastocyst formation in TSA-treated cloned mouse embryos was not associated with an increased full-term development. Similarly, Parnpai and coworkers also found that TSA treatment of cloned embryos had no effect on improving pregnancy rate in cattle and gaur (Bos gaurus) (Srirattana et al., 2008). Nevertheless, increased histone acetylation at a level similar to IVF embryos might allow production of multiple recloned generations without a reduction in cloning efficiency (Wakayama et al., 2008). It is also expected that HDACi treatment of cloned embryo might produce more of a healthy offspring with less phenotypic abnormalities (Kishigami et al., 2006; Li et al., 2008; Van Thuan et al., 2009; Zhang et al., 2007; Zhao et al., 2009), although long-term evaluation of HDACi treatment has not yet been reported in most animal species.
In conclusion, we show that treatment of donor cells with NaBu do not have a significant effect on subsequent in vitro development or embryo quality of cloned pig embryos. However, treatment of cloned embryos with NaBu improves preimplantation embryonic development. Our study also support previous reports that the effect of HDACi may vary with the type of HDACi and the target species. Further study should determine the effect of NaBu on pregnancy outcome following embryo transfer.
Footnotes
Acknowledgments
This work was supported by grants from BioGreen 12 (Code #20070401034017 and #2007040134031) program of RDA, Korea Science and Engineering Foundation (KOSEF) grant funded by the Korea Government (MEST) (No. 2009-0083571), and from Institute of Biomedical Science and Technology (IBST-2007-03-04) of Konkuk University, Republic of Korea.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.