
Abstract
Abstract
The concerns over xenogeneic pathogens and immunogenic molecules derived from mouse embryonic fibroblasts (MEFs) trigger the development of human-derived feeder layers for human embryonic stem cell (hESC) maintenance. It is essential to evaluate the capability of these human feeder layers to retain the stemness and pluripotency of hESCs. In the present study, two Chinese hESC lines, SDU-hESCm-1 and SDU-hESCm-2, were continuously cultured on human adult uterine endometrial cells (hUEC), human fetal trophonema matrix cells (hFTMC), and MEFs for at least two month (up to 10 passages). A side-by-side comparison of the abilities to support: (1) self-renewal of the hESCs, (2) expression of undifferentiated markers, and (3) neural differentiation, was made between the human and mouse feeder layers. We demonstrated that the hESCs maintained on hUEC and hFTMC exhibited significantly higher growth rates and generated higher levels of DNA content than those on MEFs. Under neural differentiation-promoting conditions, greater neural differentiation was found in the hESCs maintained on human than on mouse feeder layers. These results suggest that human feeder layers derived from hUECs and hFTMCs are more efficient in supporting a long-term growth and neural differentiation of hESCs than MEFs.
Introduction
Although the above-mentioned feeder cell types showed supportive potential, there is a lack of close comparison between human and mouse feeders. Besides the elimination of the murine elements, it is not clear if there are other advantages to using human feeder layers to maintain hESCs. In the present study, a side-by-side comparison of the abilities to support proliferation and pluripotency of hESCs was carried out between human feeder layer-derived hUECs and hFTMCs and mouse feeder layers (MEFs). We found that, like MEFs, the human feeder layers maintain major embryonic stem cell features, such as self-renewal, stemness, normal karyotype, and pluripotency after prolonged culture of at least 10 passages. We also demonstrated that human feeder cells derived from hUECs and hFTMCs support a long-term growth of hESCs through accelerating growth rate. Furthermore, under neural differentiation-promoting conditions, the hESCs cultured on hUEC or hFTMC feeder layers generated significantly more neural progenitors than those on MEFs.
Methods and Materials
Establishment and maintenance of two hESC lines (SDU-hESCm-1, SDU-hESCm-2) on MEF
Nonpronucleus zygotes were donated to this study by couples undergoing in vitro fertilization (IVF). The protocols used for IVF and for hES cell derivation were in accordance with the stem cell guidelines issued by the Ministry of Science and Technology and the Ministry of Health, and were approved by the Ethical Committee of Shandong Provincial Hospital. Since the 10 passages, hES cells began to be used for experiments.
Establishment of adult and fetal human tissue cells (hFMCs and hUECs)
Human uterine endometrium was obtained by biopsy from patients who had an explored uterine cavity and the patients had not taken exogenous hormones for 3 months before surgery and were normal by pathology test. Human trophonema matrix (6–9 weeks of gestation) was obtained individually from the chorionic villi of aborted fetus consisting of fibroblast cores and a trophoblast covering. The tissues were isolated and established using previously described methods (Chan et al., 2004). Briefly, the obtained tissue was minced finely and digested enzymatically with 0.25% collagenase (Sigma, St. Louis, MO, USA) and 10 U/mL DNase I (Sigma) in DMEM/F-12 (GibcoBRL/Invitrogen, Grand Island, NY, USA). Dissociated hUECs and hFTMCs were cultured in DMEM/F-12 supplemented with 10% fetabl bovine serum (FBS) (Hyclone, Logan, UT, USA), 4 mM glutamine (GibcoBRL/Invitrogen), 20 mM HEPES, 100 U/mL penicillin, and 100 mg/mL streptomycin (Sigma). When the cells reached confluence, they were digested with 0.125% trypsin solution and split into a 1:2 ratio into a 90-mm dish.
Immunocytochemistry
The primary antibodies used in this study were as follows: SSEA-1, SSEA-4, TRA-1-60, and TRA-1-81, purchased from Millipore/Chemicon (Billerica, MA, USA), are mouse–antihuman; OCT-3/4, purchased from Millipore/Chemicon and nestin from Santa Cruz Biotechnology (Santa Cruz, CA, USA) are rabbit–antihuman; Vimentin and Cytokeratin 18, purchased from Dako Ltd. (High Wycombe, UK), are mouse– antihuman antibodies. All antibodies were diluted in phosphate-buffered saline (PBS). Antibody localization was showed using secondary antibody conjugated to fluorescent isothiocyanate (FITC, Sigma).
Alkaline phosphatase assay
Alkaline phosphatase activity was detected with a kit containing NBT/BCIP as the substrate (Roche Molecular Biochemicals, Indianapolis, IN, USA).
Karyotyping
To analyze the karyotype of hESC, hFTMC, and hUEC, the chromosomes of the cells were visualized using G-band staining. First, cells were incubated with 0.2 μg/mL colcemid (Gibco/Invitrogen) for 6 h at 37°C in 5% CO2. Cells were then collected by centrifuge and resuspended in 0.075 M KCl solution (Sigma). After 25 min incubation at 37°C, the cells were fixed with 3:1 methanol:acetic acid before dropped onto precleaned chilled glass slides. Chromosomes were visualized using G-band staining. At least 20 metaphase spreads were counted and five Giemsa banded karyotypes were analyzed.
Teratoma formation
The hESC colonies cultured for more than 20 passages on MEF were harvested and injected with a sterile 25-gauge needle into the thigh muscle of 4-week-old severe combined immunodeficiency (SCID) mice (Slac laboratory, Shanghai). The injected mice were killed 12 weeks later, and tumors were fixed with embedded in paraffin and examined histologically after hematoxylin-eosin staining.
Reverse transcriptase-polymerase chain reaction (RT-PCR)
The confirm of differentiation of hESCs in vitro are made by RT-PCR analysis. Harvested hESC were transferred into the embryonic body (EB) culture medium and cultured continuously for 14 days to form EBs (Ma et al., 2008). EBs were collected for RNA isolation using TRIzol reagent according to the manufacturer's protocol (GibcoBRL/Invitrogen). Total RNA was quantified with a spectrophotometer (Spec3000; BioRad, Hercules, CA, USA) and RT-PCR was performed with 40 ng of total RNA in each sample. To confirm the differentiation ability of hESCs, RT-PCR was performed to analyze the expression of NF-68, keratin (ectoderm), CMP, kallikrein, enolase (mesoderm), a-FP and a1-AT (endoderm). OCT-4 was used as a positive control for undifferentiated hESC. PCR cycles consisted of an initial denaturation step at 95°C for 5 min, followed by 30 cycles of 1 min of denaturation at 94°C, 1 min of annealing at 58°C, and 1 min of extension at 72°C. A final extension was performed at 72°C for 10 min. PCR products were visualized by ethidium bromide staining following 1.2% agarose gel electrophoresis.
qPCR analysis
The levels of mRNA expression of six genes (Oct4, Nanog, Sox2, UTF1, DPPA5, and Lin41) (Cai et al., 2006) represented the undifferentiated ESC state were quantify by real-time qPCR. Total RNA was extracted from undifferentiated hESCs and retrotranscription was performed as just mentioned. PCR reactions were carried out by a 7500 sequence detection System (Applied Biosystems, Bedford, MA, USA) using a SYBR Green qPCR kit (Applied Biosystems) according to the manufacturer's instructions. The content of selected genes was normalized to the content of GAPDH, and nontemplate control was performed for every gene as blank control. Standard curves were generated using 10 ng cDNA per 20 μL reaction volume. All PCR products were checked by melting curve analysis to exclude the possibility of multiple products or incorrect product size. PCR analyses were conducted in triplicate for each sample.
Cycle/DNA profile analysis
hESC colonies cultured on each feeder layer were harvested and dissociated into single cells by trypsin-EDTA (Sigma) and 0.1% (v/v) collagenase type IV (Sigma); single-cell suspensions of about 6 × 105–1.5 × 106cells/mL in PBS were fixed in 70% ethanol at 4°C for more than 1 h. After being washed thoroughly, 20 μg/mL RNase were added to 200 μL of filtered hESC suspension and incubated at room temperature for 30 min, then 200 μL (40 μg/mL) propidium iodine stain was added (Beckman Coulter, Fullerton, CA, USA). After incubation at 0°C for 30 min, cell were stained and measured by flow cytometry (EPICS-XL, Beckman Coulter, Miami, FL, USA).Cell cycle/DNA profiles from the third to the fifth day hESC cultured on each feeder layer were analyzed by flow cytometry.
Growth curves
On the first day of passaging of hESC cultured on different feeder layer, we respectively selsected 50 clones of similar size. Then every day we chose 10 clones from them and digested them into a singular cell to calculate the mean cell population contained per clone. We use the cell number at day 1 as 100% and computed the propagate of hESC; from the second to the sixth days we continuous monitored them to get a growth curve of hESC maintained on different feeder layers.
Neural differentiation of SDU-hESCm-1 and SDU-hESCm-2 cells on three feeder layers
After the hESCs were cultured in the ES growth medium for 7 days, neural differentiation was then induced by the neural differentiation medium (NDM) consisting of DMEM/F12-based medium containing N-2 supplement (Gibco), L-glutamine, penicillin/streptomycin, and 10 ng/mL bFGF. After being cultured for another 7 days, the hESCs were physically removed from each feeder layer and dissociated into a small clump and plated on four-well plates coated with Poly-D-Lysin/laminin to culture for 5 days; then differentiated cells were detected by Immunocytochemistry.
Statistical analyses
Statistical analysis for each parameter, significance of main effects was determined using the GLM procedure of SPSS 12.0. Significance of differences among individual treatment means was determined by the least-square means method. Differences were considered significant at p < 0.05.
Results
Characterization of feeder layers derived from hUECs and hFTMCs
Primary cultures of hUECs and hFTMCs gave rise to an epithelial morphology (Fig. 1). After several passages they transformed to spindle-shaped fibroblasts. The hUECs were larger, and formed a thicker layer than hFTMCs and MEFs. Immunstaining showed that more than 90% hUECs and hFTMCs were vimentin positive, indicating their possible mesenchymal origin. G-banding of the human feeder cells showed their normal karyotype (hUEC: 46, XX and hFTMC: 46, XY).
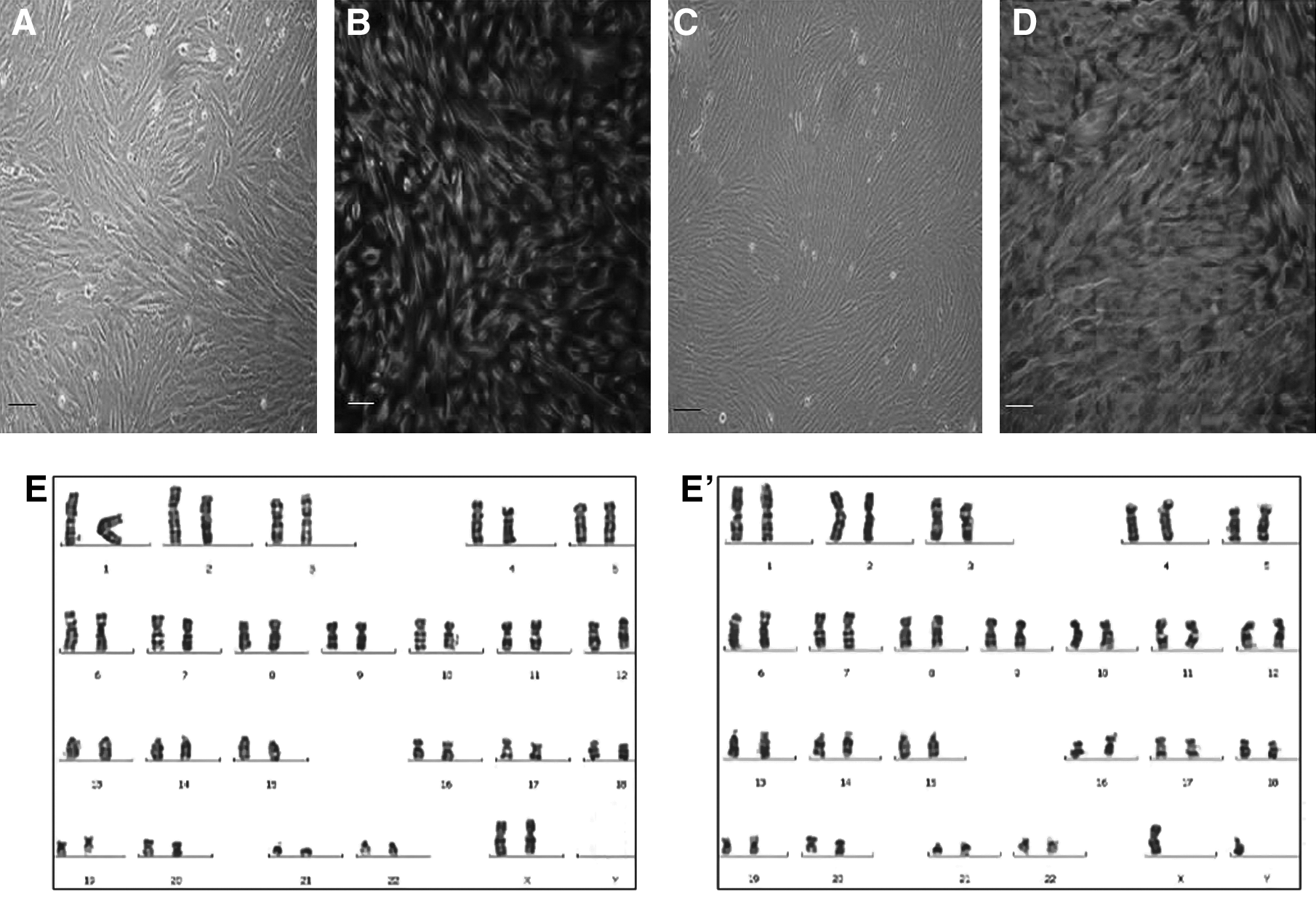
Characterization of feeder cells derived from uterine endometrial cells (hUECs) and human fetal trophonema cells (hFTMCs). (
Characterization of the Chinese hESC lines, SDU-hESCm-1 and SDU-hESCm-2, maintained on MEFs
The two Chinese hESC lines expressed transcription factors OCT-4, and glycolipids SSEA-4, TRA-1-60, and TRA-1-81, and alkaline phosphatase activity, but no SSEA-1 (Fig. 2). G-banding showed that the two Chinese hESC lines both had normal 46, XX karyotypes. At 12 weeks after being injected with undifferentiated hES cells of both SDU-hESCm-1 and SDU-hESCm-2, teratomas were formed in the abdominal cavity of the SCID-beige mice (Fig. 2). Well-differentiated tissues representing three germ layers including primitive neural tube, gastrointestinal epithelium, and cartilage were found. EBs cultured in suspension for 14 days also showed the expression of tissue-specific markers, including ectoderm markers NF-68 and Keratin, mesoderm markers CMP and Kallikrein, and endoderm markers enolase, a-FP, and al-AT (Fig. 2).
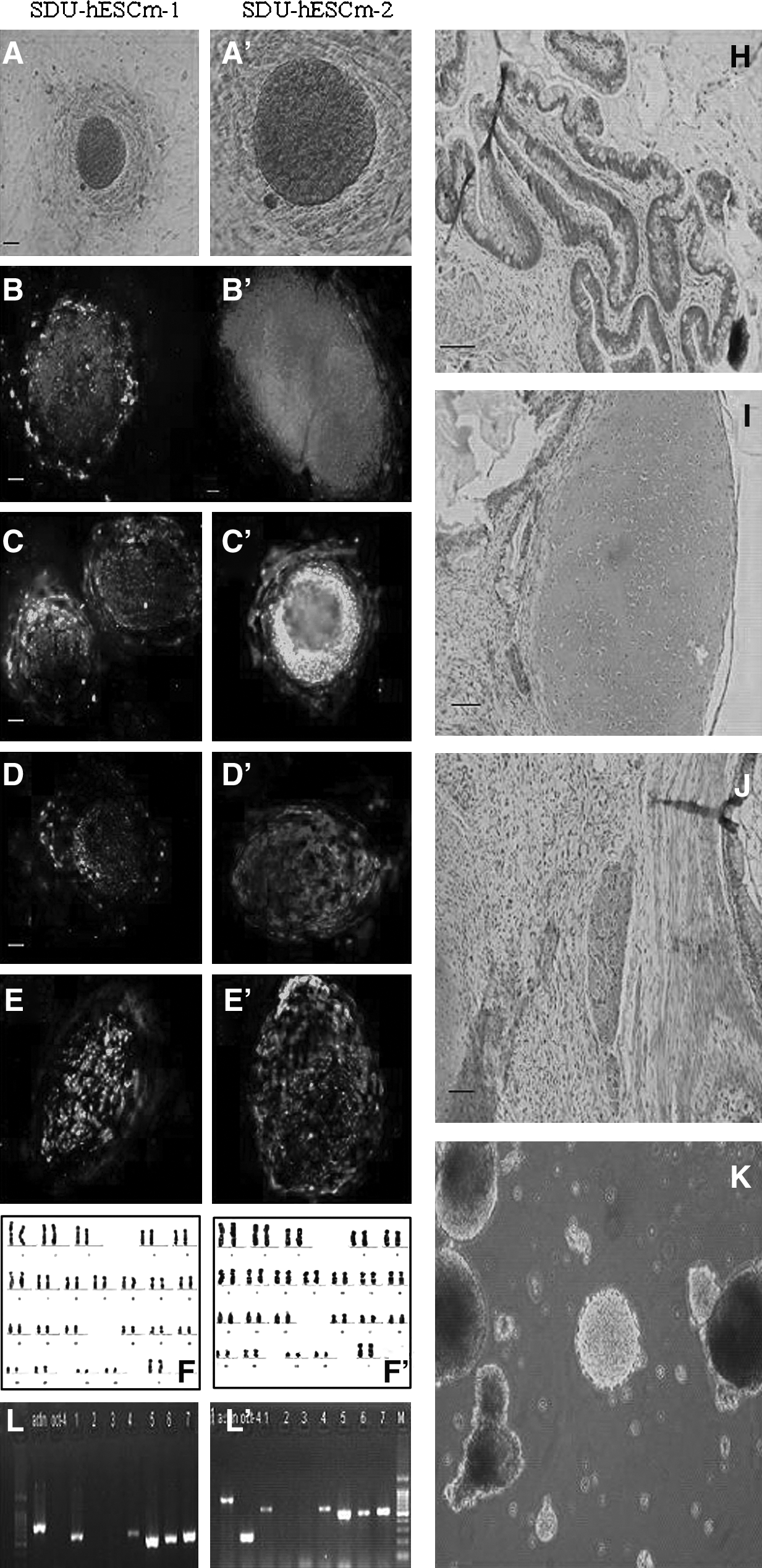
Characterization of the two Chinese hESC lines, sdu-hESCm-1 and sdu-hESCm-2, cultured on MEFs. Staining of the two Chinese hESC lines for alkaline phosphatase activity (
Expression of undifferentiated markers in two Chinese hESC lines cultured on human feeder layers
The two Chinese hESC lines were maintained on two human feeder layers, hUECs and hFTMCs; at least 10 passages showed similar expression patterns of undifferentiated markers as that on MEF. Immunocytochemistry showed that the hESCs cultured on hUECs or hFTMCs expressed SSEA-4, TRA-1-60, TRA-1-81, and OCT-4, but negative for SSEA-1 (Fig. 3A–D, A′–D′, and F–I, F′–I′). The two Chinese hESC lines maintained on hUECs and hFTMCs also showed active alkaline phosphatase activity (Fig. 3E, E′, and J,J′).
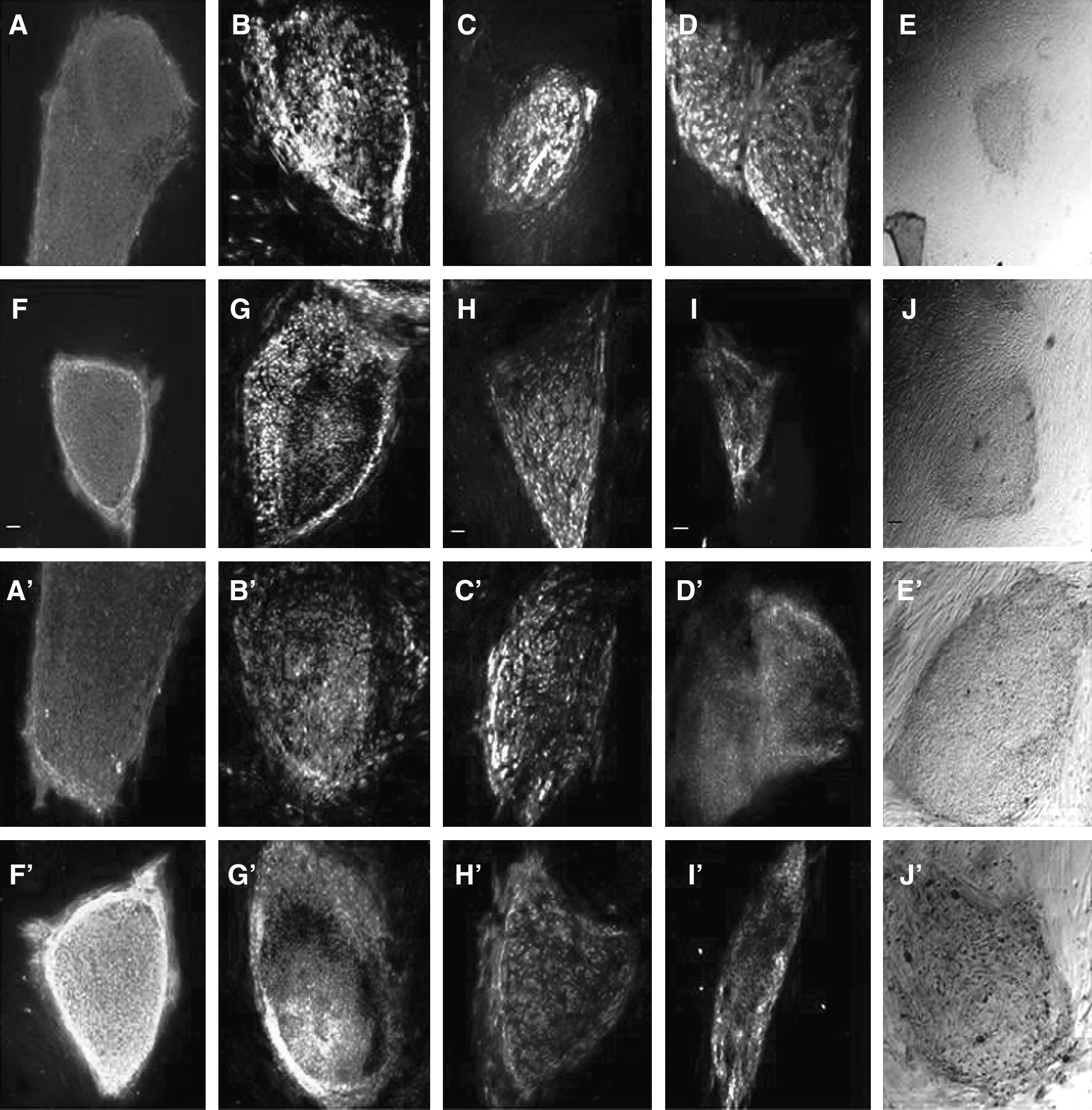
Undifferentiated markers expressed in the two Chinese hESC lines cultured on feeder cells derived from hUECs and hFTMCs. (

qRT-PCR analysis of gene expression in the two Chinese hESC lines cultured on MEF, hUEC, and hFTMC feeder layers
To compare the undifferentiated hESC state cultured on three kinds of feeder layers, we selected six genes including Oct4, Nanog, Sox2, UTF1, DPPA5, and Lin41, which were generally accepted as marker for pluripotency signature (Cai et al., 2006). The gene expression of the hESCs on MEF was considered as 100%, and the SDU-hESCm-1 cultured on the hUECs and hFTMCs showed lower expression levels than those on the MEF in six genes; gene expression levels of SDU-hESCm-2 cultured on the hUECs in Oct4 and Sox2 showed higher expression levels than those on the MEFs and hFTMCs; other gene expression levels of SDU-hESCm-2 cultured on the hUECs and hFTMCs showed lower expression levels than those on the MEFs. However, there were no significant differences in expression of two Chinese hESCs in six genes between human and mouse feeder layers (Fig. 3K and K′).
The two Chinese hESC lines maintained on hUECs and hFTMCs exhibited higher growth rates
The growth curves in Figure 4 show significant differences in percent increase in the number of cells at day 5 in culture between the three different layers. The hESCs cultured on hUEC feeder layers had greatest growth rate, the cells on MEF feeder layers had lowest growth rate (Fig. 4E and E′).
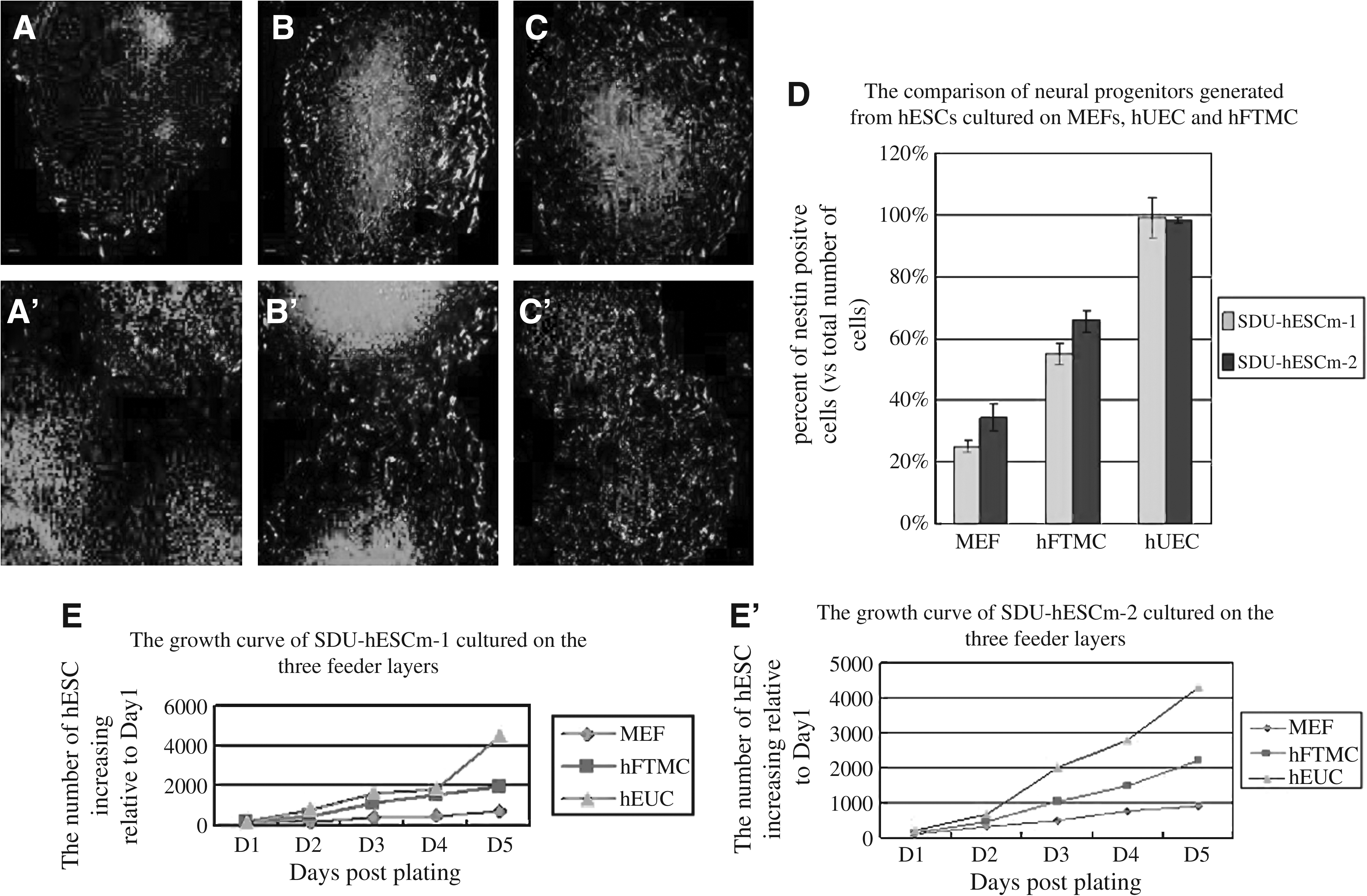
(1) The two Chinese hESC lines expansions on MEF, hUEC, and hFTMC display distinct growth characteristics under the same culture conditions. The growth curves show significant differences in percent increase in number of cells grown on three different feeders. The two Chinese hESC lines grow slower on MEFs than other two human feeders. Cells grow on hUEC faster than on hFTMC. Statistical differences for percent increases in cell numbers between hUEC or hFTMC and MEF at 5 days are significant **p < 0.01 (
Difference in DNA contents of the two Chinese hESC lines cultured on MEFs, hUECs, and hFTMCs
DNA flow cytometric analysis was performed to measure the cell cycle and DNA profile of the two Chinese hESC lines cultured on MEFs, hUECs, and hFTMCs. On the third, fourth, and fifth day of culture, the two Chinese hESC lines maintained on hUEC feeder layers showed a significant higher composition of S stage cells than that on MEF and hFTMC feeder layers (p < 0.05), especially on the third day of culture (p < 0.01). In addition, on the third day of culture, the two Chinese hESC lines expanded on the hFTMC feeder layers showed significant higher composition of S stage cells than that cultured on the MEFs (p < 0.05) (Table 1).
DNA contents were measured by flow cytometry using propidium iodine staining. A for hUEC and MEF, B for hFTMC and MEF, C for hUEC and hFTMC.
p < 0.05, **p < 0.01.
Directed neural differentiation of the two Chinese hESC lines Cultured on MEFs, hUEC, and hFTMC
Possible differences in pluripotency of the two Chinese hESC lines cultured on human and mouse feeder layers were also examined during the directed neural differentiation. Neural differentiation was induced by added neural differentiation medium into the two Chinese hESC lines cultures on hUEC, hFTMC, and MEF feeder layers. After 7 days in the neural differentiation medium, the hESCs were removed from each feeder layer to PDL/laminin-coated substrates for another 5 days. Immunostained nestin-positive cells were manually counted and were expressed as a percentage of the total DAPI labeled cells. A significant difference was found in the number of nestin-positive cells differentiated from the two Chinese hESC lines cultured on human and mouse feeder layers (Fig. 4A–C and A′–C′). The greatest percentage Nestin-positive cells (99.0 ± 6.7% and 98.2 ± 1.04%) were found in the two Chinese hESC lines cultured on hUEC feeder layers; next were on hFTMC feeder layers (55 ± 3.2% and 65.7 ± 3.21%), and the lowest was on MEF feeder layers (25 ± 1.8% and 34.6 ± 4.27%).
Discussion
In the present study, a number of cellular and molecular techniques were used to examine the abilities of human feeder layers derived from adult uterine endometrial cells (hUECs) and fetal trophonema matrix cells (hFTMCs) to support the growth, stemness, genetic stability, and pluripotency of human embryonic stem cell lines in comparison with mouse feeder layers (MEFs). We demonstrated that both mouse and human feeder layers maintained major hESC features after prolonged culture at least 10 passages. The two Chinese hESC lines cultured on these three feeder layers expressed similar undifferentiated markers, had a normal karyotype and teratoma formation. However, the two Chinese hESC lines cultured on human feeder layers derived from hUECs and hFTMCs showed higher growth rate and greater neural differentiation capability compared to those on MEFs. These results suggest that human feeder layers derived from hUECs and hFTMCs, in addition to eliminating risk factors from xenogeneic feeders, are more efficient in supporting a long-term growth and neural differentiation of hESCs than MEFs.
Our finding about hUECs and hFTMCs supporting the two Chinese hESC lines cell growth is consistent with previous reports that the feeder layers derived from human uterine endometrial cells have the advantage that they can be used for many passages, whereas MEF feeder cells can only be used as feeder cells for a limited number of passages (Lee et al., 2005). It is important to point out that the supportive potential of the MEFs, hUECs, and hFTMCs was indirectly evaluated by measuring expression of stem cell markers on cocultured hESCs. As a result, the major factors that contribute to suitability of feeder cells for long-term culture of hESCs remain largely unclear. On the other hand, although the molecular mechanisms underlying the role of feeder cells in hESC culture are not well known, it is believe that feeder cells play at least two roles in maintaining undifferentiated hESCs: (1) providing a solid surface and expressing adhesion ECM molecules to enhance hESC attachment, and (2) supporting the growth and survival of hESCs through the production of growth factors (Eiselleova et al., 2008). During development in vivo, undifferentiated hESCs are surrounded by the hUECs and hFTMCs; therefore, we had intention to recreate an in vivo hESC niche, thereby generating the requisite intracellular signals to allow self-renewal rather than differentiation of these cells. It was found that production of TGFβ1, activin A, and FGF-2 varied considerably among the human-derived feeder cell lines (Eiselleova et al., 2008). The difference in the ability of human and mouse feeder layers to promote the undifferentiated growth of hESCs may be attributable to its characteristic growth factor production. Mouse feeder cells are superior in the production of activin A, but do not secrete any measurable FGF-2. Conversely, human feeder layers have the ability to release up to several tenths pg/mL FGF-2 every 24 hours. The hUECs express many growth factors (IGF, EGF, and TGFb), cytokines (CSF, LIF, and interleukin-1 and −6) (Luo et al., 2003), and cell adhesion molecules (ECMs and integrin) (Selam et al., 2002) throughout the menstrual cycle. Some of these factors are known to be key regulators related to hESC elf-renewal. The hFTMCs are primarily composed of fibroblasts, derived from extraembryonic mesoderm, and epithelial cells, termed trophoblasts. During development in vivo, inner cell mass are surrounded by the trophoblasts. Undifferentiated hESC derived from inner cell mass indicate that human fetal trophonema have more tightly link with hESC, thereby generating the requisite intracellular signals to allow self-renewal rather than differentiation of these cells (Olga et al., 2005). hUECs and hFTMCs both are ideal candidates for the growth of hESC.
The further work required to establish clinical grade feeder cell lines for hESC line culture is significant and costly. Much work is being done to find feeder-free culture systems but these are at an early stage of development and there may be consequences that affect the value of the hESCs for research and development. These challenges should be viewed in the context of the huge amount of work that will be required over many years to develop robust differentiation protocols and establish fully defined procedures and adequate safety data for embryonic stem cell products.
Footnotes
Acknowledgments
This study was supported by the grants from the Key Point Project of Science and Technology Development of Shandong (2002DD1DBA1), the National High-tech R&D Program of China (863 Program) (2006AA02Z4A4), the National Basic Research Program of China (973 Program) (2006CB944004), and National Natural Science Foundation of China (30801237 and 30670777).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.