
Abstract
Abstract
This study was conducted to direct porcine embryonic stem (pES) cells differentiating into neural lineages and to investigate therapeutic potential of GFP-expressing pES (pES/GFP+) in the rat model of Parkinson's disease (PD). Directed differentiation of pES into neural lineages was induced by suspension culture in medium containing RA, SHH, and FGF combinations without going through embryoid body formation. A high yield of nestin-expressing neural precursors was found in all treatments on day 2 after the 12-day induction. On day 6 after replating, more than 86.2 and 83.4% of the differentiated cells stained positively for NFL and MAP2, respectively. The expression of TH, ChAT, and GABA specific markers were also observed in these NFL-positive neural cells. The undifferentiated pES/GFP+ cells and their neuronal differentiation derivatives were transplanted into the Sprague-Dawley (SD) rat's brain, and their survival and development was determined by using live animal fluorescence optical imaging system every 15 days. The results showed that fluorescent signals from the injection site of SD rats' brain could be detected through the experimental period of 3 months. The level of fluorescent signal detected in the treatment group was twofold that of the control group. The results of behavior analysis showed that PD rats exhibited stably decreased asymmetric rotations after transplantation with pES/GFP+-derived D18 neuronal progenitors. The dopaminergic differentiation of grafted cells in the brain was further confirmed by immunohistochemical staining with anti-TH, anti-DA, and anti-DAT antibodies. These results suggested that the differentiation approach we developed would direct pES cells to differentiate into neural lineages and benefit the development of novel therapeutics involving stem cell transplantation.
Introduction
Domestic swine are demonstrated very similar to the human in anatomic, immunologic, and physiologic characteristics; and the sizes of their organs are fairly comparable to that of human (Phillips and Tumbleson, 1986; Prelle et al., 1999). Moreover, swine have been demonstrated as excellent animal models in therapeutics development for various human diseases, including congenital heart disease (Swindle et al., 1992), hypertension (Zambraski et al., 1992), organ transplantation (Flye, 1992; Hall et al., 1986), pharmacology, and toxicology (Feletou and Teisseire, 1992; Kurihara-Bergstrom et al., 1986). Although the establishment of pluripotent ES cell lines from domestic species is much more difficult than that in murine species, porcine ES (pES) cell lines have been successfully derived from inner cell mass of the blastocysts (Chen et al., 1999a; Li et al., 2003, 2004). The pES cells were very similar to hES cells in many characteristics, including colony morphology, feeder-dependent, and refractory to leukemia inhibitory factor (LIF) in culture, and expression of stage-specific embryonic antigen-3/4 (SSEA-3/4) but not SSEA-1, which is characterized only to murine ES cells (Chen et al., 1999a; Brevini et al., 2007; Yang et al., 2009). Therefore, the study of pES cells might serve as an excellent model in development of regeneration medicine in human.
Parkinson's disease is a degenerative disorder characterized by progressive and selective loss of dopamine (DA) neurons in the midbrain substantia nigra (Olanow and Tatton, 1999). This disorder has being a prime target for cell replacement therapy, given over a decade of successful clinical experiences with fetal ventral mesencephalic cell transplantation in PD patients (Piccini et al., 1999). However, fetal cells transplantation has significant technical, ethical, and practical limitations, partly due to limited availability and variable outcomes (Freed et al., 2001). Due to the self-renewal capacity and multilineage developmental potential, stem cells could be ideal sources for cell replacement therapy (Park et al., 2005; Taylor and Minger, 2005). The challenge in using ES cells for regenerative medicine has been to direct the wide differentiation potential toward the derivation of a specific cell fate. Within the nervous system, the ES cells have been shown to differentiate in vitro into neural progenitor cells, neurons, and astrocytes. Recently, several researches described the conditions to direct ES cells into neurons of midbrain dopaminergic identity by coculture with various cells, with chemically defined condition medium, hormones, and growth factors (Kawasaki et al., 2000; Park et al., 2005; Perrier et al., 2004; Iacovitti et al., 2007; Yan et al., 2005; Yang et al., 2008). These successful induction protocols have provided a powerful tool to control the development and function of ES-derived midbrian DA neurons and made a promise of ES cell therapy on PD in the future.
In this study, we investigated the potential of directed differentiation of pES cells into neural lineages and to demonstrate a valuable yield of neural precurors. We also transplanted pES cells and their derived cells of various neural differentiation stages into the nigrostriatal-lesioned PD rats. The growth and differentiation of the grafted cells in vivo, and the functional recovery of the PD animals were also determined.
Methods
In vitro culture of pES and GFP-expressing pES (pES/GFP+) cells
The pES cell line used in this study was line M215-3, which was derived from the preimplantation blastocyst of the Livestock Research Institute Black Pig No. one (a topcrossing breed established from Taoyuan and Duroc pigs) (Chen et al., 1991, 1999a, 1999b). The GFP-expressing pES (pES/GFP+) cell line used in this study was obtained from electroporation of pES M215-3 with green fluorescent protein (GFP) and nominated as pES/GFP+ 10 (Yang et al., 2009). Both of pES M215-3 (63 to 65 passages) and pES/GFP+ 10 (41–45 passages) have a 36+XX normal karyotype. They were maintained in ES-cell culture medium (ESM) consisted of Dulbecco's modified eagle medium (DMEM; high glucose and no pyruvate; Invitrogen, Grand Island, NY, USA) supplemented with 1 mM L-glutamine (Sigma-Aldrich, St. Louis, MO, USA), 0.1 mM β-2-mercaptoethanol (Sigma-Aldrich), 10 mM MEM nonessential amino acids (Sigma-Aldrich), 0.03 mM adenosine (Sigma-Aldrich), 0.03 mM guanosine (Sigma-Aldrich), 0.03 mM cytidine (Sigma-Aldrich), 0.03 mM uridine (Sigma-Aldrich), 0.01 mM thymidine (Sigma-Aldrich), antibiotics (50 units/mL penicillin G and 50 μg/mL streptomycin sulfate, Invitrogen) and 16% fetal bovine serum (FBS; Invitrogen). The pluripotent pES cells were propagated on a feeder layer of mitomycin C (Sigma-Aldrich) inactivated STO cells (mouse embryonic fibroblasts, CRL-1503, USA) in gelatin-coated Multidish 4 Wells® (Nunc 176740, Roskilde, Denmark) in ESM at 39°C with an atmosphere of 5% CO2 in air. For the maintenance of undifferentiation, pES colonies were regulatory subcultured with 0.25% trypsin–0.02 mM EDTA (Invitrogen) and replated onto fresh STO feeder layers every 5 to 7 days.
In vitro induction of neural differentiation
The neurogenic stimulators used for neural induction in the present study were retinoic acid (RA, 1 μM, Sigma-Aldrich), sonic hedgehog (SHH, 200 ng/mL, R&D Systems, Minneapolis, MN, USA), and fibroblast growth factor (FGF, 100 ng/mL, Sigma-Aldrich), respectively, as described by Barberi et al. (2003). Confluent pES colonies were trypsinized and subjected to a two-step in vitro induction protocol for neural differentiation. They were first taken into a single-cell suspension culture at a concentration of 1 × 105 cells/mL in an Easy Flask® (Nunc 169900) with serum-free ESM containing each of the five different neurogenic stimulator combinations: (1) RA, (2) RA + SHH, (3) RA + FGF, (4) SHH + FGF, and (5) RA + SHH + FGF for neural induction. After 12 days of induction, the pES cell-derived cells were replated onto gelatin-coated four-well dishes in 5 × 103 cells/mL. These cells were cultured in expanding medium containing DMEM/F12 medium (Sigma-Aldrich), 1:50 B27 supplementation (Invitrogen), 2 mM L-glutamine, 50 units/mL penicillin G, and 50 μg/mL streptomycin sulfate (Invitrogen), and supplemented with 20 ng/mL human recombinant epidermal growth factor (hrEGF; Invitrogen), 20 ng/mL human recombinant basic fibroblast growth factor (hrbFGF; Invitrogen), and 1:100 N2 (Invitrogen).
Transplantation and in vivo tracking of pES/GFP+ cells
The pES/GFP+ cells of three different differentiation stages upon neural induction were used for transplantation. They were the original D0 pES/GFP+ cells without induction, the pES/GFP+-derived cells collected at the end of the 12-day neural induction, which were regarded as the D12 neuronal progenitors, and the pES/GFP+-derived neuronal progenitors, which collected on day 6 after replating postneural induction, which were regarded as D18 neuronal progenitors, respectively. These cells were fully trypsinized into single-cell suspension and adjusted to a concentration of 1 × 106 cells per 5 μL in phosphate-buffered saline (PBS; Sigma-Aldrich) directly before transplantation.
For determining the survival and development of grafted cells in vivo, a total of 39 male Sprague-Dawley (SD; BioLASCO, Taiwan) rats of 200–250 g were randomly subjected to control group (n = 8) and treatment groups of D0 pES/GFP+ cells (n = 11), D12 neuronal progenitors (n = 10), and D18 neuronal progenitors (n = 10). The rats were anesthetized with chloral hydrate (400 mg/kg body weight in 0.9% NaCl, i.p.) and each 5 μL of PBS containing 1 × 106 cells were injected into the striatum of rat in the treatment groups with the assistance of a stereotaxic instrument [anterior–posterior (AP) = 0.0 mm, medial–lateral (ML) = −3.0 mm, and dorsal–ventral (DV) = −5.0 mm] (Paxinos and Watson, 1986). Each rat in the control group was injected with 5 μL of cell-free PBS. During the 3-month experimental period, in situ monitoring of these grafted cells were determined by using two live animal fluorescence optical imaging systems every 15 days. The In Vivo Imaging System (IVIS 50, Xenogen Corp., Alameda, CA, USA) was used for noninvasive tracking. The relative fluorescence values of treatment groups were calculated as the fluorescence intensity detected from the region of interest (ROI) on rats in each treatment groups divided by that of the control group at the same time window. At the end of in vivo tracking study, rats were all sacrificed and their brains were collected for final IVIS imaging analysis.
All animal experiments in this study were carried out in accordance with ethical guidelines and following approval of the Livestock Research Institutional Animal Care and Use Committee.
Nigrostriatal lesion and cell transplantation in rat model of PD
For nigrostriatal lesion, each well-anesthetized male SD rats was injected with 3 μL of 6-hydroxydopamine (6-OHDA, 5 μg/μL; Sigma-Aldrich) mixed with 0.05% ascorbic acid at a rate of 1 μL/min into the medial forebrain bundle (nigrostriatal dopaminergic pathway) under the assistance of a stereotaxic instrument (AP = −4.4 mm, ML = −1.2 mm, DV = −7.8 mm; and AP = −4.0 mm, ML = −0.8 mm, DV = −8.0 mm) (Paxinos and Watson, 1986). Four weeks after 6-OHDA treatment, a total of 26 rats revealed more than 300 asymmetric rotations per hour in response to D-amphetamine (Sigma-Aldrich) challenge were considered as well-lesioned PD models (Park et al., 2005), and were used as recipients for cells transplantation. They were randomly subjected into control group (n = 4), sham group (n = 4), and treatment groups, which transplanted with D0 pES/GFP+ cells (n = 5), D12 neuronal progenitors (n = 7), and D18 neuronal progenitors (n = 6), respectively. A dose of 5 μL PBS containing 1 × 106 of pES/GFP+ cells or their derived cells at different differentiation stages was injected into the striatum of each PD model rats in the treatment groups stereotaxically (AP = 0.0 mm, ML = −3.0 mm, DV = −5.0 mm). Each PD model rat in the control group was injected with 5 μL of cell-free PBS, wheres those in the sham group were sham-operated to expose the injection site only.
Amphetamine-induced rotation behavioral analyses
Amphetamine-induced rotation test for PD model rats was carried out monthly after cell transplantation by using an automated Rotameter System (TSE, Germany) (Hudson et al., 1993; Löscher et al., 1996). After receiving intraperitoneal injections of 2 mg/kg D-amphetamine (dissolved in 0.9% saline), each PD model rat was placed in the rotameter bowl. Rotation patterns of the rats were recorded and the number of complete (360o) rotation over a 120-min period was calculated and analyzed by a computerized Rotameter Software 303300-S. The relative rotation rate of each animal was obtained by division of the complete rotation numbers after and before cell transplantation.
Immunocytochemical staining of neural differentiation and transplantation
Cells for immunocytochemical analysis were fixed in 10% formalin at room temperature for 30 min. The cells were washed twice with PBS, then rinsed with 0.3% Triton X-100 (Sigma-Aldrich) in PBS for 10 min and 3% H2O2 (Sigma-Aldrich) for 5 min. Followed, the cells were incubated in blocking buffer (5% FBS in PBS) at room temperature for 2 h. Subsequently, the cells were washed with PBS and incubated overnight with 1:200 dilution of primary antibody. Thereafter, the cells were washed with PBS and then incubated with the secondary antibody for a minimum of 2 h, followed by washing in PBS.
Primary antibodies used for determining the undifferentiation status of pES cells were ES cell-specific markers including octamer-binding transcription factor-4 (Oct-4, Chemicon Cat. #AB3209, Temecula, CA, USA), alkaline phosphatase (AP, Chemicon Cat. #MAB4349), stage specific embryonic antigen-3 (SSEA-3, Chemicon Cat. #MAB4303), stage specific embryonic antigen-4 (SSEA-4, Chemicon Cat. #MAB4304), tumor related antigen-1-60 (TRA-1-60, Chemicon Cat. #MAB4360), and tumor related antigen-1-81 (TRA-1-81, Chemicon Cat. #MAB4381). Primary antibodies used for determining the pES-derived cells were specific against nestin (Chemicon Cat. #AB5922), neurofilament protein light (NFL, Chemicon Cat. #AB9568), microtubule associated protein 2 (MAP2, Chemicon Cat. #AB5622), glial fibrillary acidic protein (GFAP, Chemicon Cat. #IHC2078-6), A2B5 (Chemicon Cat. #MAB312R), tyrosine hydroxylase (TH, Chemicon Cat. #AB152), choline acetyltransferase (ChAT, Chemicon Cat. #MAB5270), and γ-aminobutyric acid (GABA, Chemicon Cat. #MAB316). The secondary antibodies used were the fluorescein (FITC)-conjugated AffiniPure Goat Antirabbit IgG (H + L) (for Oct-4, nestin, NFL, MAP2, GFAP, and TH staining, Jackson ImmunoResearch Cat #111-095-003, West Baltimore Pike, PA, USA), Rabbit Antimouse IgG (H+L) (for AP, SSEA-4, ChAT, and GABA staining, Jackson ImmunoResearch Cat #315-095-003), Rabbit Antirat IgM (for SSEA-3 staining, Jackson ImmunoResearch Cat #312-095-020), and Rabbit Antimouse IgM + IgG (for TRA-1-60, TRA-1-81, and A2B5 staining, Jackson ImmunoResearch Cat #315-095-044). All of primary and secondary antibodies were hybridized in a 1:200 dilution. The undifferentiated mouse ES D3 cells (mouse pluripotent embryonic stem cells, ATCC CRL-1934, USA) were stained as positive controls for pluripotent markers.
In the end of behavioral analyses, PD rats were sacrificed with an overdose of chloral hydrate (600 mg/kg i.p.) and perfused with chilled 0.9% NaCl followed by 4% paraformaldehyde in 0.1 M PBS. Brains were dissected and fixed for 6 h, and then transferred to 30% sucrose in 0.1 M PBS for a minimum of 24 h. Sections containing substantia nigra and striatum were collected and processed for immunohistochemical staining. Free-floating sections were rinsed with 0.3% Triton X-100 and treated with 3% H2O2 to inhibit residual endogenous peroxidase. Followed, the sections were incubated in blocking buffer (5% FBS in PBS) at room temperature for 2 h, then washed with PBS and incubated overnight with primary antibody. Thereafter, the sections were washed with PBS and incubated with the secondary antibody for a minimum of 2 h, followed by washing in PBS.
The primary antibodies for determining the grafted cells were specific against hrGFP (Stratagene Cat. #240142, La Jolla, CA, USA), tyrosine hydroxylase (TH, Chemicon Cat. #AB1542, Temecula, CA, USA), dopamine (DA, Chemicon Cat. #MAB309), and dopamine transporter (DAT, Chemicon Cat. #MAB369). The secondary antibodies used were the fluorescein (FITC)-conjugated AffiniPure Goat Antirabbit IgG (H+L) (for hrGFP, staining, Jackson ImmunoResearch Cat #111-095-003), the rhodamine (TRITC)-conjugated AffiniPure Rabbit Antisheep IgG (H+L) (for TH staining, Jackson ImmunoResearch Cat #313-025-003), Rabbit Antimouse IgG (H+L) (for DA staining, Jackson ImmunoResearch Cat #315-025-003), and Goat Antirat IgG (H+L) (for DAT staining, Jackson ImmunoResearch Cat #112-025-003). All the primary and secondary antibodies were hybridized in a 1:200 dilution. Fluorescent cells were visualized using an inverted fluorescent microscopy (Axiovert 200M, Carl Zeiss, Germany) equipped with a cooled CCD camera system (CoolSNAPHQ2 Monochrome, Photometerics, USA), and analyzed with MetaMorph 6.Or5 (Universal Imaging, Ypsilante, MI, USA).
For localization of the nucleus, the immunocytochemical and immunohistochemical staining of each sample was accompanied with 4′,6-diamidino-2-phenylindole (DAPI) staining.
The fluorescein-conjugated secondary antibodies were carried out without addition of the primary antibodies as negative controls. There was no nonspecific binding of the secondary antibodies exhibited.
Statistical analysis
All results were derived from at least three independent experiments in this study. The results of the induction efficiency of differentiation, the engrafting and behavioral studies were all presented as mean ± SEM. Data were analyzed by analysis of variance using the General Linear Model (GLM) procedure and Student's t-test of SAS (SAS Institute, 1996). A significant difference was determined as the p-value was less than 0.05.
Results
In vitro culture of pES and pES/GFP+ cells
Both pES and pES/GFP+ cells were maintained on feeder layers of mitotically inactivated STO and formed colonies of typical undifferentiated morphology (Fig. 1) (Yang et al., 2009). They were epithelial-like compact colonies with abundant lipid-like vacuoles. The cells comprising pES colonies were translucent with little cytoplasm and a large nucleus containing two or more prominent nucleoli, similar to that of murine ES cells but slightly darker in color. The border of the colony was distinct, but not the boundaries between cells. Monitoring of undifferentiation status of pES cells was performed by daily visual observation and regular determination of their expression of ES cell-specific markers including Oct-4, AP, SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 (Fig. 2) (Yang et al., 2009).

The colony morphology of undifferentiated pES cells of line M215-3. The pES cell colonies were maintained on feeder layers of mitotically inactivated STO cells and formed epithelial-like compact colonies. (Scale bar: 200 μm.)
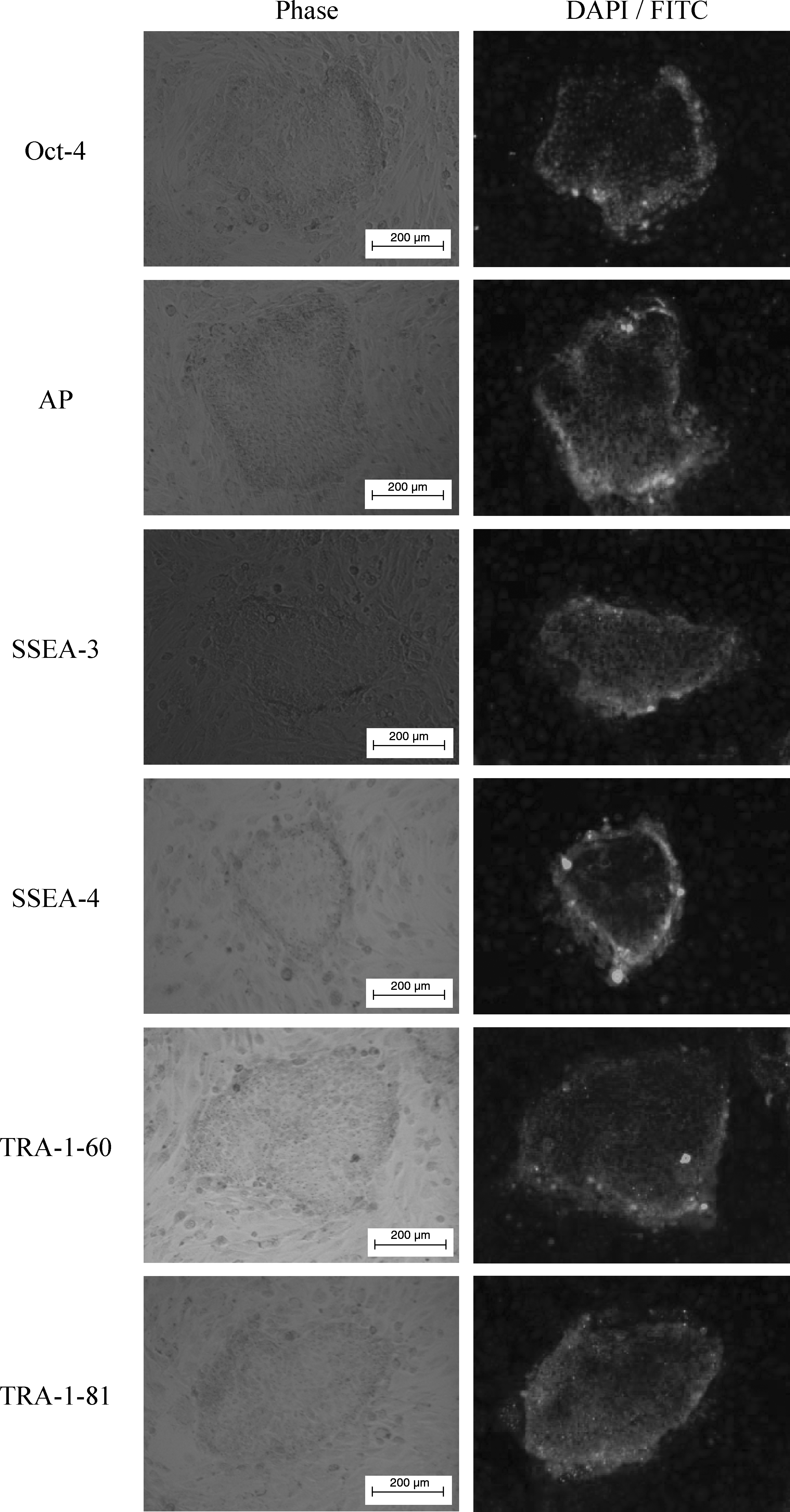
Expression of pluripotent markers Oct-4, AP, SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 of the pES colonies determined by immunocytochemical staining. (Scale bars: 200 μm.).
In vitro induction of pES cells into neural differentiation
In vitro neural differentiation of pES cells was induced by suspension culture in medium containing different combinations of neurogenic stimulators and then replated onto gelatin-coated four-well dishes in expanding medium. In all treatments, the deriving cells formed a neural network-like structure on day 6 after replating (Fig. 3A). The cells with morphology of unipolar-, bipolar-, and multipolar-like neurons with both neuron cell body and axon structures were also observed on day 6 after replating (Fig. 3B, C, and D). The response of pES cells to the neural differentiation induction in this study was very similar to that of hES cells described by Zhang et al. (2001).
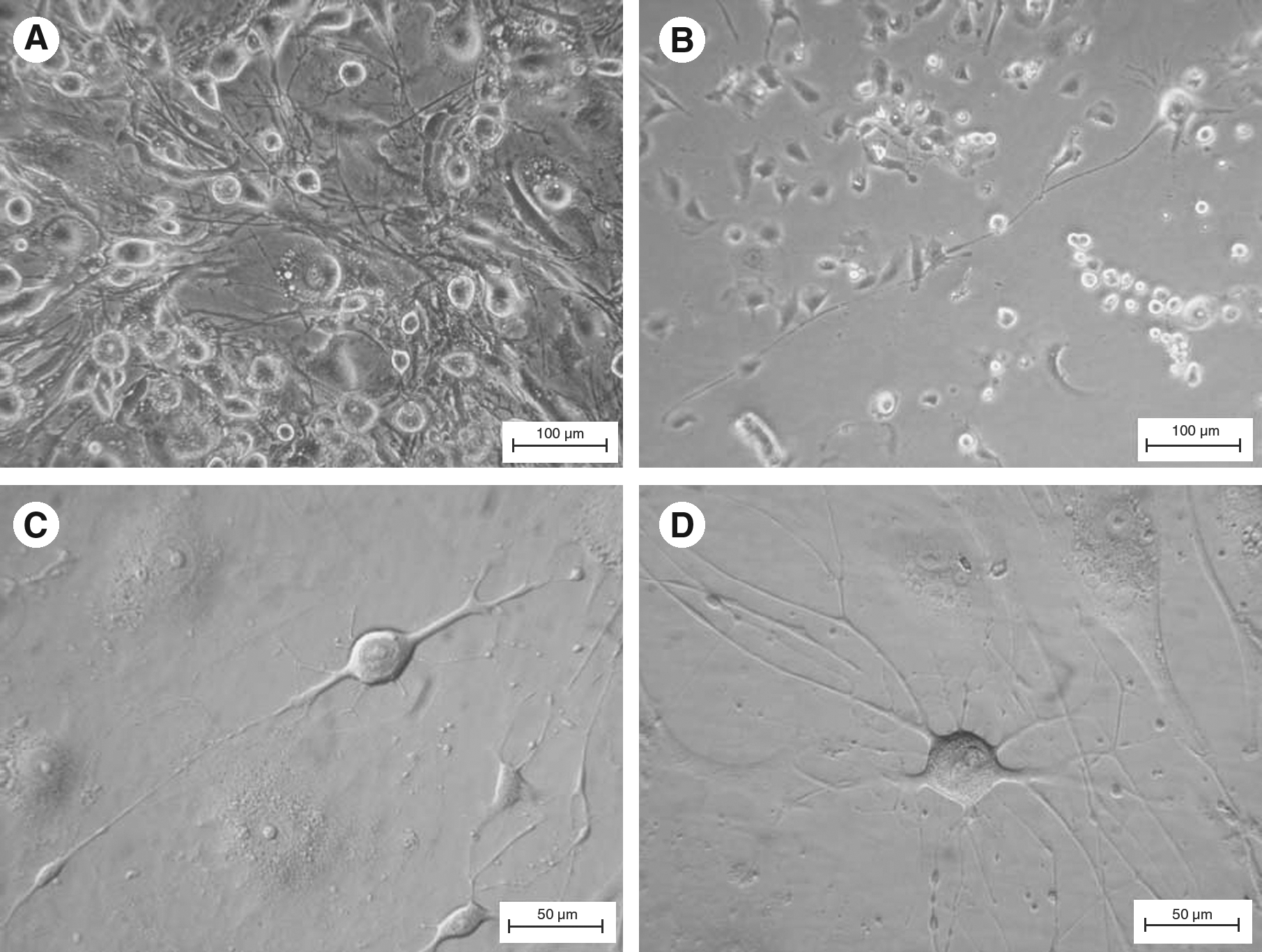
The morphology of pES-derived cells under directed differentiation. The neural network structure was observed on day 6 after replating (
Efficiency of differentiation induction of pES cells into neural lineages
The immunocytochemical analysis with neural specific antibodies including nestin, NFL, MAP2, GFAP, A2B5, TH, ChAT, and GABA were used to identify the expression of neural lineage-specific markers of the cells differentiated from pES cells after induction and expanding culture. The results indicated that more than 86.1% of the cells in all treatments were positively stained by antibody specific against nestin on day 2 after replating (Table 1 and Fig. 4A). There was no significant difference in the expression of this neural precursor marker for the cells among treatment groups (p > 0.05).

Immunocytochemical staining of pES-derived cells. After 12 days of directed differentiation, the differentiated cells expressing nestin were found on day 2 after replating (
The values in the same row with different letters were significantly different (p < 0.05).
RA, retinoic acid; FGF, fibroblast growth factor; SHH, sonic hedgehog.
On day 6 after replating, more than 86.2 and 83.4% of differentiated cells were stained positively for NFL and MAP2, respectively (Fig. 4B and C). The expression of these two mature neuron markers among treatments were also not significant (p > 0.05) (Table 1). As determined by GFAP staining, the best efficiency of astrocytic differentiation for pES cells among treatments was only 54.3%, which was induced by RA + SHH + FGF (p < 0.05). On the other hand, the efficiency of oligodendrocytic differentiation induced by RA (84.8%) was better than those treated by RA + SHH, RA + FGF, and SHH + FGF (p < 0.05) as determined by A2B5 staining (Table 1). The expressions of several lineage-specific biochemical markers, including TH, ChAT, and GABA, were observed while expanding culture. The expression patterns and levels of TH (dopaminergic, Fig. 4D), ChAT (cholinergic, Fig. 4E), and GABA (GABAergic) specific markers of these NFL-positive neural cells in different treatments were shown in Table 2. When compared to other four treatments, pES cells treated with RA-only rarely differentiated into dopaminergic lineage (8.7%, p < 0.05). However, RA-only treatment seemed to much favor the pES cells for the GABAergic differentiation among all other treatments (80.5%, p < 0.05). On the other hand, all the five different neurogenic stimulator combinations tended to have the similar efficiency on directing the pES cells to differentiate into cholinergic cells (p > 0.05).
The values in the same row with different letters were significantly different (p < 0.05).
In vivo tracking and immunocytochemical staining of grafted pES/GFP+ cells
The results of in vivo tracking for grafted cells using IVIS 50 showed that fluorescent signals from pES/GFP+ cells in the injection site of SD rats' brain could be detected through the experimental period (Fig. 5), and the relative intensity of fluorescent signal detected in treatment groups were all twofold above that of the control group. The highest relative fluorescence intensity detected in rats transplanted with D0 pES/GFP+ cells, D12 neuronal progenitors, and D18 neuronal progenitors was 3.61 ± 0.46 (on day 45, n = 11), 2.99 ± 0.35 (on day 60, n = 10), and 1.97 ± 0.52 (on day 45, n = 10) folds of that of the control (n = 8), respectively (Table 3). Rats were sacrificed in the end of experiment and further confirmed that the fluorescence signals were luminesced from the fluorescent porcine cells that transplanted into their brain previously (Fig. 6). The presence of grafted pES/GFP+ related cells in the injection site of rat's brain were further confirmed by immunohistochemical staining with anti-GFP antibody after scarification at the end of in vivo tracking (Fig. 7). These results suggested that the pES/GFP+ cells and their derived neuronal progenitors could serve as a traceable target in the cell transplantation trials, and provided as a valuable study model for the development of novel therapeutics involving the transplantation of stem cells.
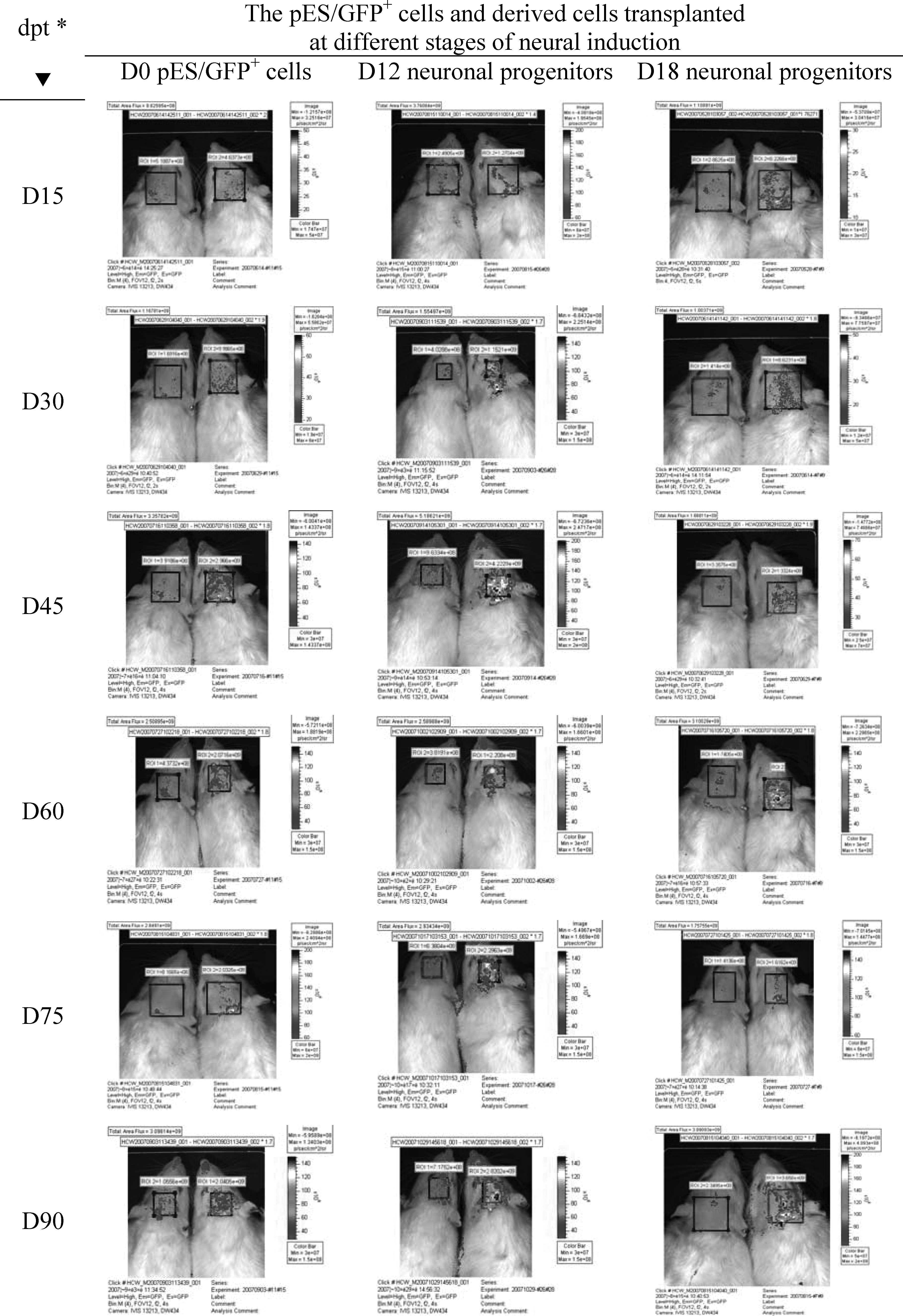
Determination of the fluorescence signals from SD rats grafted with D0 pES/GFP+ cells, D12 neuronal progenitors, and D18 neuronal progenitors. Images showed the continuous tracking of the same rats with In Vivo Imaging System (IVIS 50, Xenogen Corp., Alameda, CA, USA) though 3 months of experimental period. (Left: control; right: treatment; *dpt: days posttransplantation.)
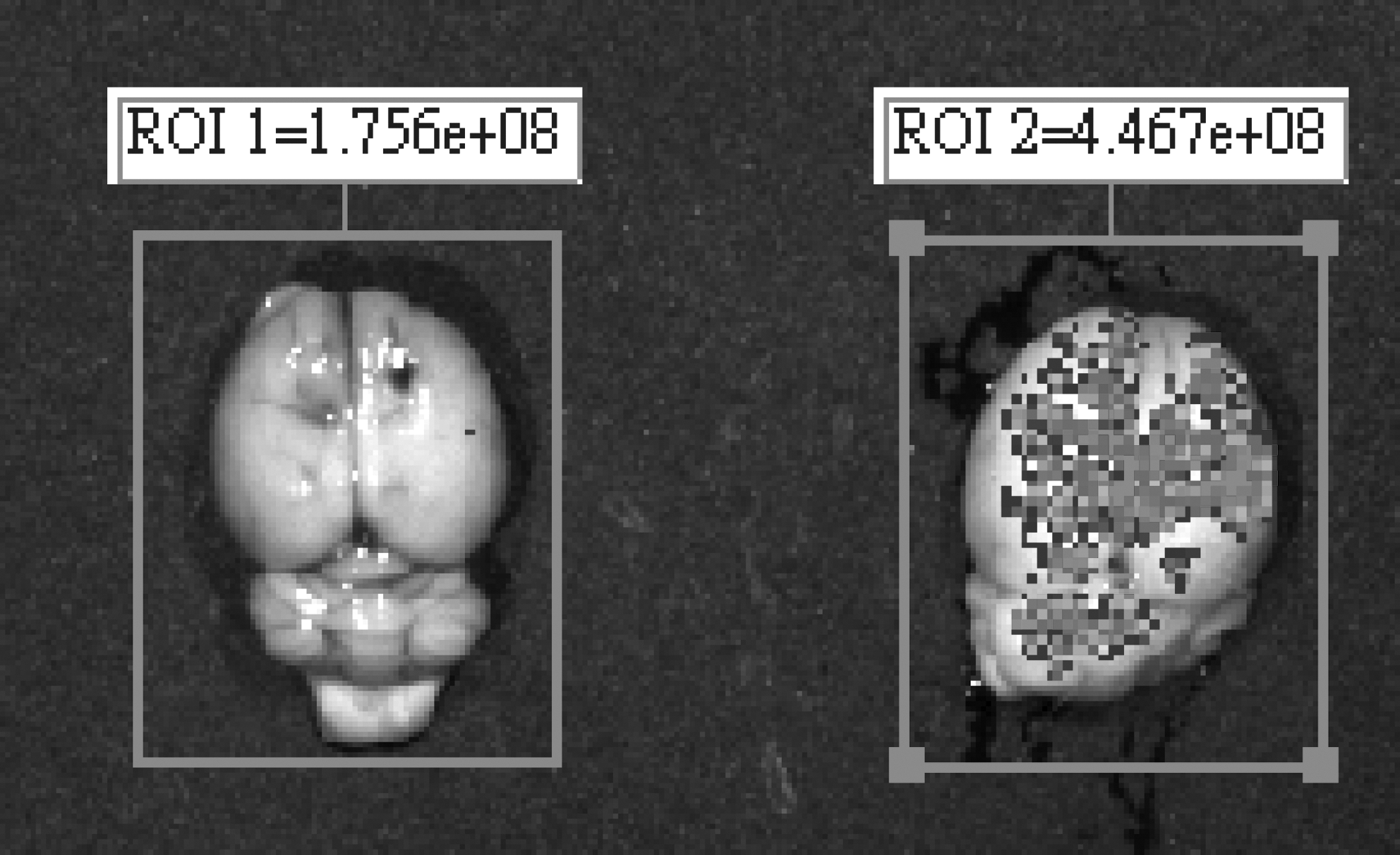
The IVIS image of SD recipient's brain confirmed that the fluorescence signals were luminesced from the pES/GFP+ cells that transplanted into the brain previously. (Left: control; right: treatment.)
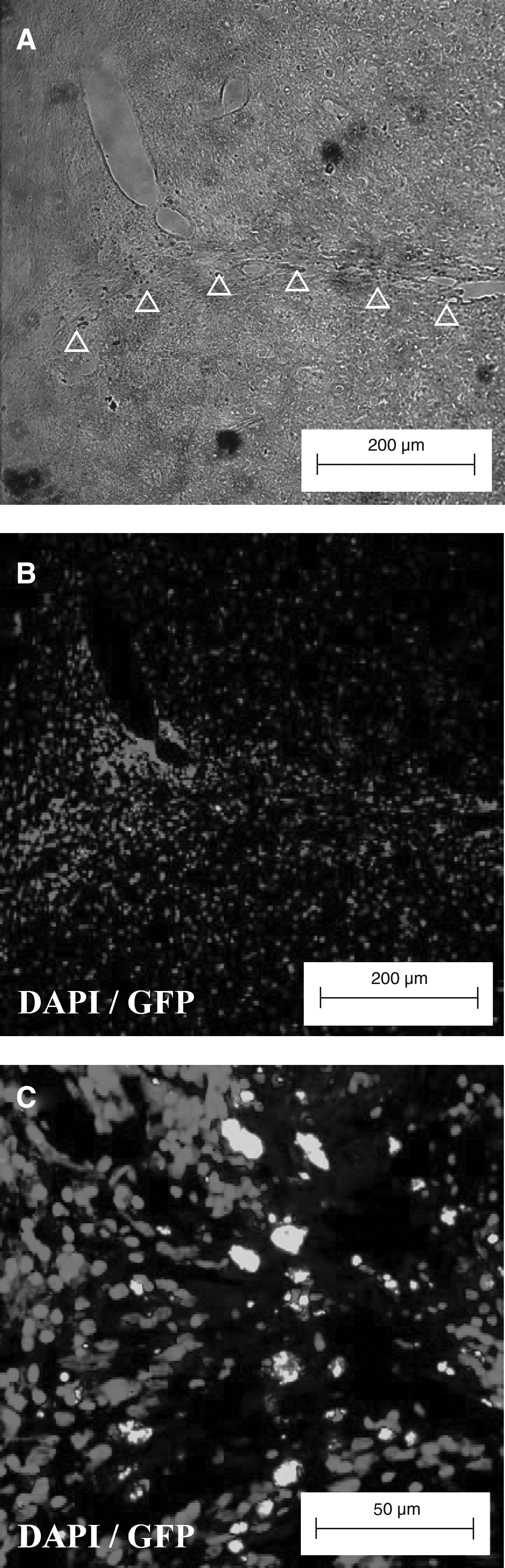
The presence of grafted pES/GFP+ cells in the injection track (Δ) (
dpt, days posttransplantation.
Relative fluorescence intensity was calculated as the fluorescence intensity detected from the region of interest (ROI) of the rats in treatment group divided by that detected from the rats in the control group at the same time window.
The values in the same column with different letters were significantly different (p < 0.05).
Nigrostriatal lesion and cell transplantation in the rat model of PD
A total of 77 SD rats were subjected to 6-OHDA treatment for nigrostriatal lesioning. Four weeks later, 26 (33.8%) of them were determined as well lesioned and suitable to be used as PD model animals, by using the rotational asymmetry test under the influence of D-amphetamine. They were then divided randomly to control (n = 4), sham (n = 4), and three treatment groups, which were transplanted with D0 pES/GFP+ cells (n = 5), D12 neuronal progenitors (n = 7), and D18 neuronal progenitors (n = 6), respectively. The results of amphetamine-induced rotation behavioral analysis showed that the relative rotation rate detected at the first, second, and third month after treatment was 1.00 ± 0.00, 1.25 ± 0.21, and 1.43 ± 0.21 for the control group, whereas it was 1.33 ± 0.18, 1.54 ± 0.15, and 1.43 ± 0.08 for the sham group, respectively. For the PD rats transplanted with D0 pES/GFP+ cells, their relative rotation rate detected at the first, second, and third month after treatment was 0.82 ± 0.17, 1.11 ± 0.20, and 1.82 ± 0.22, respectively. The PD rats transplanted with D12 neuronal progenitors exhibited a relative rotation rate of 0.91 ± 0.15, 1.25 ± 0.18, and 1.35 ± 0.10 at the first, second, and third month after treatment, whereas the relative rotation rate of the PD rats transplanted with D18 neuronal progenitors was 1.05 ± 0.12, 1.01 ± 0.11, and 0.92 ± 0.02, respectively. The results of progressive increase of relative rotation rates over the 3-month period after treatment for the PD rats grafted with D0 pES/GFP+ cells and D12 neuronal progenitors indicated that there was no functional recovery after cell transplantation. However, the PD rats grafted with the D18 neuronal progenitors exhibited stable decreases in relative rotation rates along the time after treatment and reached a improvement with significant difference at the third month when compared to that of the control group (p < 0.05) (Fig. 8). In addition, immunohistochemical analysis of the rats' brains after behavioral analyses further confirmed that the grafted pES/GFP+-derived D18 neuronal progenitors still survived and differentiated into dopaminergic lineage in the striatum as determined by staining with anti-TH, anti-DA, and anti-DAT antibodies (Fig. 9).
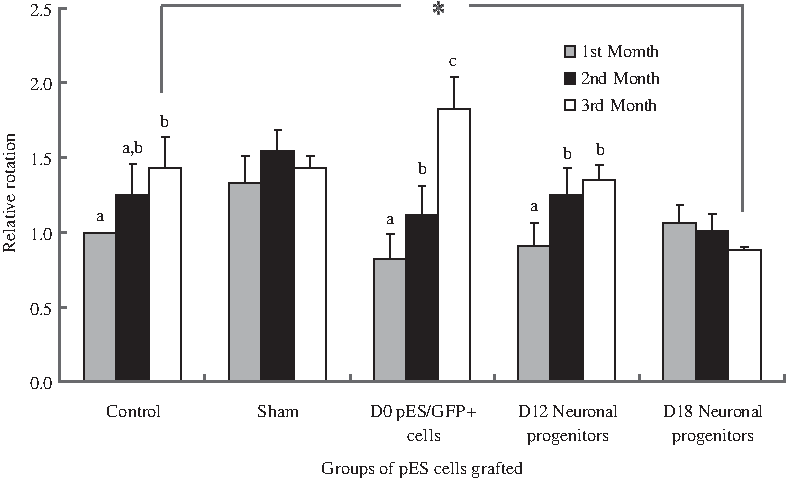
Functional recovery of the PD rats in the control (n = 4), sham (n = 4), and treatment groups of which grafted with D0 pES/GFP+ cells (n = 5), D12 neuronal progenitors (n = 7), and D18 neuronal progenitors (n = 6) were analyzed by amphetamine-induced rotation test. Relative rotation rates were presented as mean ± SEM for each animal as compared to its pretransplantation evaluation. The levels of relative rotation with different letters in the same group and * symbol between groups were significantly different (p < 0.05).
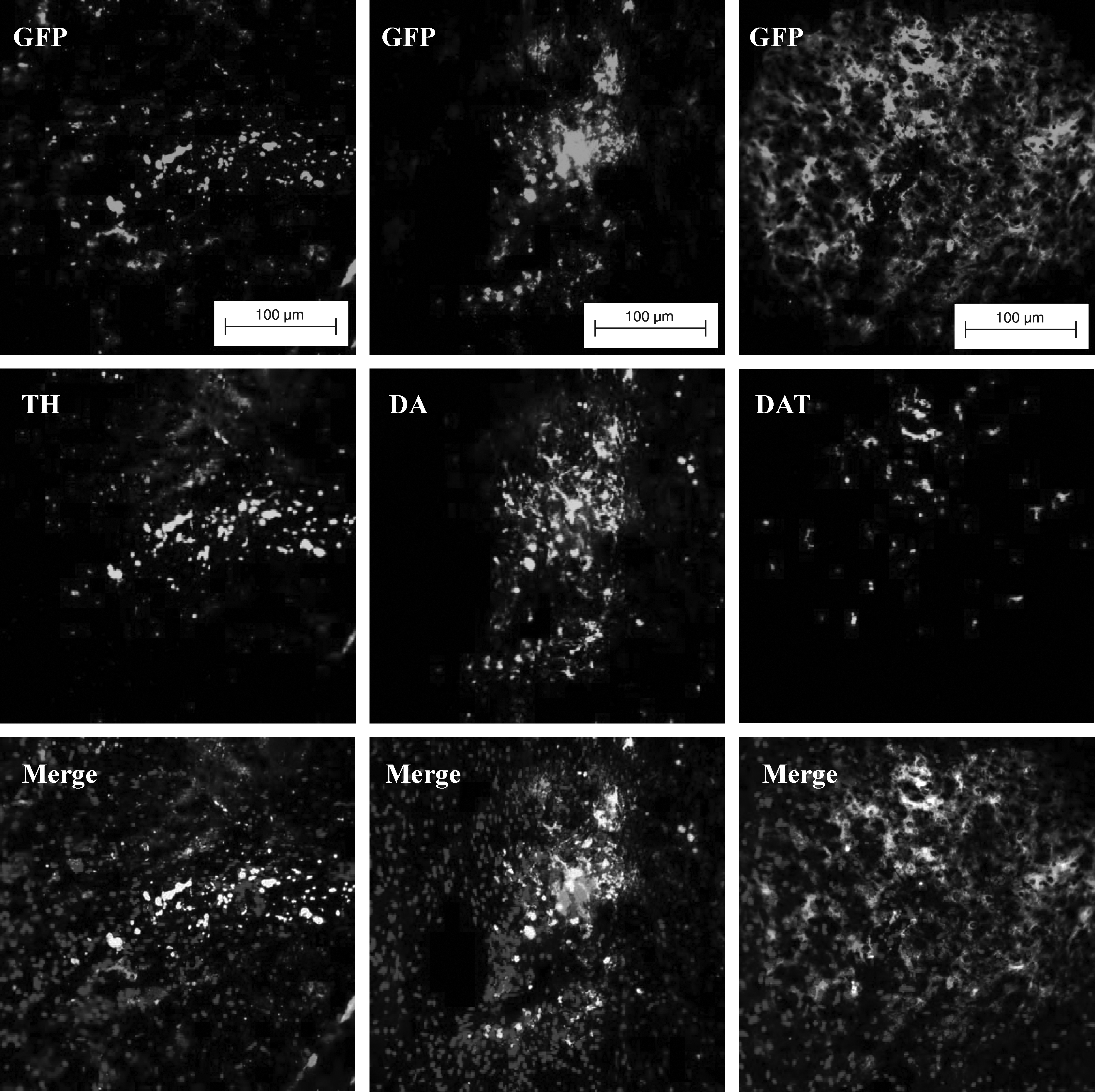
Grafted pES/GFP+-derived D18 neuronal progenitors were identified by anti-TH, anti-DA and anti-DAT antibodies and indicated dopaminergic differentiation. (Scale bar: 100 μm.)
Discussion
The challenge for using ES cells in developmental biology and regenerative medicine has been to direct the wide differentiation potential toward the derivation of a specific cell fate. Although the ES cells can give rise to a large variety of cell types in vitro, they appear to have a bias for neural lineages. They seem to be very readily used for neural cells generation, and that appears to be a default differentiation pathway (Aberdam et al., 2007; Smukler et al., 2006; Tropepe et al., 2001). Two major strategies were used to differentiate ES cells into the neural fate, including embryoid body (EB)-based lineage selection (Barberi et al., 2003; Carpenter et al., 2001; Lee et al., 2000; Li et al., 2005; Martinat et al., 2006; Rodríguez-Gómez et al., 2007; Strübing et al., 1995; Wichterle et al., 2002; Yan et al., 2005; Zhang et al., 2001), and coculture with PA6 cells that possesses stromal cell-derived inducing activity (SDIA) (Brederlau et al., 2006; Kawasaki et al., 2000; Kim et al., 2006; Thinyane et al., 2005; Yoshizaki et al., 2004; Zeng et al., 2004). The in vitro aggregation of ES cells into EBs was spontaneous and random, and always led to generation of the heterogeneous populations and did not lead to the desired pure population of a single cell lineage (Jüngling et al., 2003; Schulz et al., 2003). Recently, Zhou et al. (2008) indicated that the quality and seeding density of EBs were the two critical factors might influence the efficiency of neural differentiation from mouse ES cells. They enriched the nestin-positive cells from 50% up to approximately 92% by producing high-quality EBs and seeding them in a low density (Zhou et al., 2008). However, the efficiency of specification differentiation of these neural precursors was relatively low and diversified. For promoting the efficiency of neural differentiation, supplement the medium with growth factors is known to be effective for the propagation of neuroepithelial progenitors (Svendsen et al., 1998; Vescovi et al., 1999). Under in vitro conditions, the frequency of neuronal cells emerged from spontaneous differentiation of mES and hES cells were low (Reubinoff et al., 2000; Strübing et al. 1995), but was significantly increased by RA addition (Guan et al., 2001; Schmidt et al. 2001). Retinoic acid is a morphogen and plays an important role in caudalizing neuroepithelia during development (Maden, 2002). Study on RA-induced differentiation of hES cells demonstrated efficient neural differentiation via EB formation and resulted in largely specified neurons of GABAergic and glutamatergic lineages (Carpenter et al., 2001; Guan et al., 2001). Neural inducing techniques based on conditioned media without going through the step of EB formation had been developed for directing differentiation of ES cells into neural cells (Cazillis et al., 2005). Therefore, we hypothesized that exclusion of EB formation from the protocol and supplement of growth factors for neural induction might benefit the generation of homogenous neural cell types from pES cells.
In the previous studies, we have demonstrated that the pES and pES/GFP+ cells were pluripotent and could be induced to differentiate into cells of all three primary germ layers, including neural lineages (Chen et al., 1999a, 1999b; Yang et al., 2009). In the present study, the efficiency of different morphogen combinations of RA, SHH, and FGF on neural differentiation induction in pES cells were compared. We found that all the different combinations efficiently enriched the population of nestin-expression neural progenitor cells (>86.1%) and promoted the generation of mature neurons of NFL- and MAP2-positive at a high frequency above 86.2 and 83.4%, respectively (Table 1). These results might suggest that our EB-free induction protocol was an efficient approach for directing the pES cells to differentiate toward neural lineages.
Identification of the specifically differentiated neuronal populations deriving from pES cells was determined by immunocytochemical analysis (Table 2). Specification differentiation induced only by RA seemed to be the least efficient to give rise to dopaminergic and astrocytic differentiations. These results of astrocytic and dopaminergic differentiation induced by RA only in this study were 7.7 and 8.7%, respectively. It was similar to the reports using RA to direct the differentiation of hES cells (Carpenter et al., 2001) and mES cells (Perrier et al., 2004). On the other hand, RA-only induction seemed to prefer directing pES cells toward GABAergic differentiations (80.5%) when compared to other treatments. These results accorded with the reports of RA-induction in hES cells (Guan et al., 2001; Schmidt et al. 2001). Furthermore, an enhanced effect on generating A2B5-positive cells (84.8%) and a low efficiency of generating GFAP-positive cells (7.7%) was found simultaneously in the RA treatment. This differentiation diversity of glial lineages from pES cells responding to the RA induction was unclear and needed to be studied further.
In this study, treatments with individual and combining addition of RA, SHH, and FGF led a greater increase of TH-positive cells to a proportion above 89.4%, much greater than that induced by RA-only treatment (Table 2). The crucial roles of SHH and FGFs in the patterning of the ventral midbrain and hindbrain for in vivo development have been described (Ye et al., 1998). Lee et al. (2000) further demonstrated that the signaling of SHH and FGF could direct the ES cell-derived neuroepithelial cells toward dopaminergic differentiation. The results in our study also confirmed that the treatments of RA + SHH, RA + FGF, SHH + FGF, and RA + SHH + FGF could effectively increase the yield of dopaminergic neurons deriving from pES cells.
A revolutionizing approach of in vivo imaging for cell biological research, gene expression, and tumor progression is the fluorescent proteins, and the usefulness of them as noninvasive probes for in vivo tracking (Chudakov et al., 2005; Hadjantonakis et al., 2002; Hoffman, 2008; Popova et al., 2008; Yang et al., 2009). The continued tracking of grafted cells on recipients has a profound meaning of their distribution, development, progression, and migration when stem cells were introduced into the model organism. Although numerous researches have reported the detection of GFP in visceral organs of live nude mice, GFP has been considerably more difficult to detect or has been virtually undetectable in severe combined immunodeficiency (SCID) mice (Winnard et al., 2006). In the present studies, the pES/GFP+ cells we previously established could be detected and monitored in situ with noninvasive optical instruments after they were grafted. The results showed that fluorescent signals from pES/GFP+ cells at the injection site of SD rats' brain could be detected through the experimental period of 3 months. The intensity of fluorescent signals detected in treatment groups was about twofold stronger than that of the control group. We sacrificed the SD rats at the end of monitoring experiment and further confirmed that the fluorescence signals were luminesced from the pES/GFP+, which transplanted into the brain previously. We simplified the distribution tracking of grafted pES/GFP+ cells based on the IVIS 50 optical imaging in living animals, and provided a convenient and continued monitoring approach rather than a chromogenic substrate such as lacZ and firefly luciferase, and prelabeling methods such as magnetic resonance imaging (MRI) and positron emission tomography (PET).
Brain is the immune-privileged site due to the blood–brain barrier, an atypical physical barrier (Ferguson et al., 2002). Drukker et al. (2002) reported that the undifferentiated stem cells expressed low levels of major histocompatibility complex class I (MHC-I), which is the pivotal element for graft rejection. However, the tumorgenic potential will still be a major concern if human ES-derived cells were to apply to clinic (Brederlau et al., 2006). Several researches reported the grafted mES (Bjorklund et al., 2002; Nishimura et al., 2003; Thinyane et al., 2005) and hES (Brederlau et al., 2006) might give rise to teratomas. The necessity of nearly pure population without carried over tumorgenic stromal cells and undifferentiated cells to reduce the low tumorgenesis had been suggested by Park et al. (2005) and Yang et al. (2008). In the present study, no tumorgenesis was found at transplantation site when examined histologically. This might be due to the collection of nearly purified pES/GFP+ cells population for injecting by mechanical method, and the directed differentiation excluded mesodermal and endodermal cells for grafting. In addition, the lack of the tumorgenesis of our grafted pES/GFP+ and their derived cells might be related to the low tumorgenic potential of in vivo teratoma formation when injected into SCID mice (Yang et al., 2009).
In our porcine–rat xenografting therapy model of PD, each does of 1 × 106 pES/GFP+ cells of different differentiated stages (pES/GFP+ cells, D12 and D18 neuronal progenitors) were used for therapeutic transplantation in PD model rat. The relative rotation rate did not decrease through the 3 months experimental period for the PD rats transplanted with D0 pES/GFP+ cells and their derived D12 neuronal progenitors. Contrary, the PD rats grafted with D18 neuronal progenitors showed a stable decrease of relative rotation rates along the 3-month experimental period. This result suggested that the Parkinsonian behavioral defects of the PD rats could be improved by transplantation with much differentiated pES-derived neuronal progenitors. Several researches concentrated on the therapeutic potential of ES cells in animal models of PD, but the results of functional recovery were various. Several researches indicated that grafted mES- or hES-derived dopaminergic neurons could integrate into the host striatum, exhibited extensive neural fiber outgrowth, and significantly reduced functional deficits in animal models of PD (Barberi et al., 2003; Bjorklund et al., 2002; Kim et al., 2002; Martinat et al., 2006; Nishimura et al., 2003; Rodríguez-Gómez et al., 2007; Shim et al., 2004; Takagi et al., 2005; Yang et al., 2008). Some researches showed partial and modest recovery from Parkinsonian behavioral defects though the grafted mES- or monkey ES-derived cells survived and innervated in the host striatum (Muramatsu et al., 2009; Yoshizaki et al., 2004). However, negative results on improving rotational behavior deficits in PD modeling animals were also observed in the transplantation of hES-derived dopaminergic cells; and the immunohistochemical analyzes indicated that TH+ cells exhibited on the injection site, but no extended TH+ neural fiber and neural connection observed (Brederlau et al., 2006; Park et al., 2005; Zeng et al., 2004). The inconsistency might be attributable to the cell preparation, differentiation protocol, survival and functional exhibition of grafted dopaminergic neurons. The presence of dopaminergic neurons alone is not sufficient for function contribution, and the intrinsic factors such as host microenvironment, trophic support, and immunological factors must be taken into consideration to achieve in vivo development of grafted cells (Drukker, 2008; Park et al., 2005; Yang et al., 2008).
In conclusion, the directed differentiation induction approach of pES cells developed in this study can provide a valuable yield on the process of in vitro differentiation of pES cells into functional neurons. Also, transplantation of pES-derived D18 neuronal progenitors prepared by this approach also shows a potential possibility for the treatment of PD. The ES cells of porcine origin offer some distinct advantages over other species and might serve a better research model because they are immunologically and physiologically much more similar to the humans. The information obtained in this study involving directed differentiation induction and the application of the resultant cells to improve the function of PD model rats suggested the potential of further application of pES cells in the study of replacement medicine and functional degenerated pathologies.
Footnotes
Acknowledgments
This work was supported from the grants of NSC93-2313-B-061-002 by National Science Council, 96AS-1.1.3-LI-L1 and 97AS-1.1.3-LI-L1 by the Livestock Research Institute, Council of Agriculture, and TCSP-01-02 by Buddhist Tzu-Chi General Hospital, Taiwan.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.