
Abstract
Abstract
Adherent umbilical cord blood stromal cells (AUCBSCs) are multipotent cells with differentiation capacities. Therefore, these cells have been investigated for their potential in cell-based therapies. Quantum Dots (QDs) are an alternative to organic dyes and fluorescent proteins because of their long-term photostability. In this study we determined the effects of the cell passage on AUCBSCs morphology, phenotype, and differentiation potential. QDs labeled AUCBSCs in the fourth cell passage were differentiated in the three mesodermal lineages and were evaluated using cytochemical methods and transmission electron microscopy (TEM). Gene and protein expression of the AUCBSCs immunophenotypic markers were also evaluated in the labeled cells by real-time quantitative PCR and flow cytometry. In this study we were able to define the best cellular passage to work with AUCBSCs and we also demonstrated that the use of fluorescent QDs can be an efficient nano-biotechnological tool in differentiation studies because labeled cells do not have their characteristics compromised.
Introduction
Cells that are similar to MSCs from bone marrow (BM), have been reported to be present in the adipose tissue, dental pulp, placenta, umbilical cord blood (UCB), and from a selection of fetal tissues, such as spleen, lung, pancreas, kidneys, and amniotic fluid during midgestation (Bianco et al., 2008; Chen et al., 2006; Liu et al., 2009; Mimeault and Batra, 2008). Despite several common characteristics among the MSCs derived from different tissues, various studies show differences regarding cellular differentiation capacity. Therefore, more investigation is necessary, both on a cellular and molecular basis (Bobis et al., 2006; Meirelles et al., 2008).
Classically, MSCs are able to adhere to plastic and express CD29, CD44, CD73, CD90, CD105, and major histocompatibility complex (MHC) type class I antigens, low or nonexpression of hematopoietic cell markers CD14, CD34, and CD45, and absence of MHC class II antigens (Dominici et al., 2006; Flynn et al., 2007; Jin et al., 2009; Liu et al., 2009; Majumdar et al., 1998; Oh et al., 2008).
Another important marker is the CD146, which was established as being a colony-forming unit fibroblast marker (Sacchetti et al., 2007). CD146 is involved in a cell-to-cell junction, and it is present in primitive MSCs (Bardin et al., 2001; Covas et al., 2008; Shi and Gronthos, 2003; Yoshimura et al., 2006). It is also expressed in the signaling pathway that comprises the tyrosine kinases FYN, FAK, and paxillin (Anfosso et al., 2001; Covas et al., 2008). The activation of FAK and paxillin pathways are involved in the MSCs osteogenic differentiation by the interaction with extracellular matrix compounds, such as vitronectin or laminin-5 (Baksh et al., 2007; Covas et al., 2008; Kundu and Putnam, 2006; Salasznyk et al., 2007a, 2007b).
Several MSCs characteristics, such as the potential to differentiate into multiple lineages and the ability to be expanded ex vivo, make these cells very interesting targets for therapeutic use in regenerative medicine and tissue engineering (Dominici et al., 2006; Flynn et al., 2007; Jin et al., 2009; Liu et al., 2009; Oh et al., 2008).
The culture system diversity and the lack of distinct nomenclature for a nonequivalent cell population reflect the difficulty of understanding MSCs as a standardized population. These cells have been named MSCs, Marrow Stromal Cells, Marrow-isolated Adult Multilineage inducible (MIAMI) cells and Multipotent Adult Progenitor Cells (MAPCS) (Baksh et al., 2004; Meirelles et al., 2008; Reyes et al., 2001; Verfaillie et al., 2005).
The nomenclature for MSCs has been recently put forward by the International Society for Cellular Therapy (Horwitz et al., 2005; Meirelles et al., 2008) and discussed at several international meetings. In this study we named MSCs as Adherent Umbilical Cord Blood Stromal Cells (AUCBSCs).
AUCBSCs are obtained after full-term delivery from the newborn umbilical cord. It is a sample that is usually discarded and derived from a painless and noninvasive procedure, without harm to the mother or the infant (Berger et al., 2006; Devine et al., 2001; Jin et al., 2009; Kim et al., 2006; Newcomb et al., 2006; Oh et al., 2008; Rao and Mattson, 2001; Rubinstein et al., 1993; Swanger et al., 2005; Tanaka and Liang, 1995; Yang et al., 2004).
AUCBSCs have been extensively investigated for their potential in cell-based therapies. A marker to track these cells in vivo would be very helpful; therefore, we investigated the in vitro stability of quantum dots as a AUCBSCs marker.
Quantum dots are small highly fluorescent photostable nanoparticles that have been recently adopted for in vitro use (Jaiswal, 2003; Larson et al., 2004; Muller-Borer et al., 2007). Advantages of QDs include a narrow band emission and broadband excitation with a high quantum yield, photostability, luminescence, and resistance to chemical and metabolic degradation (Alvisiatos, 2004; Chan et al., 2002; Jaiswal, 2003; Muller-Borer et al., 2007). Reports suggest that QDs are noncytotoxic (Dubertret et al., 2002; Jaiswal, 2003).
However, detailed analysis reports some cytotoxicity of QDs and reveals it is due to different physicochemical properties, dosage, and exposure concentrations (Braydich-Stolle et al., 2005; Hardman, 2006; Hsieh et al., 2006; Lovric et al., 2005; Muller-Borer et al., 2007; Seleverstov et al., 2006). Most QD studies have utilized nonmammalian or cancer cells, only few studies examined the effect of QDs in MSCs (Hsieh et al., 2006; Jaiswal, 2003; Muller-Borer et al., 2007; Rosen et al., 2007; Seleverstov et al., 2006).
In this study, we demonstrate the fluorescent QDs labeling can be used as an efficient nano-biotechnological tool, due to its stability and capability of preserving the characteristics of the labeled cells. In addition, using cytochemistry and ultrastructural techniques, we determined the AUCBSCs decreasing cellular viability in later passages, defining the best passage for experiments with these cells.
Materials and Methods
Harvest of human UCB samples
Fifteen UCB units were collected after obtaining informed consent from the donating mothers in accordance with the Ethics Committee of Instituto Israelita de Ensino e Pesquisa Albert Einstein (São Paulo, Brazil). Informed consent was obtained from all subjects. These UCB samples were collected by venous puncture of the umbilical vein at the time of the full-term delivery and conserved with 100 mM of EDTA anticoagulant at 22°C and soon after processed within 6 h of collection.
Culture of human UBC-derived AUCBSCs
Mononuclear cells (MNCs) were separated by centrifugation in the Ficoll-Hypaque gradient (density 1.077 g/mL; GE, Syracuse, NY, USA). MNCs were seeded into 25-cm2 flasks at a density of 1 × 107 to 108 cells/cm2 containing Dulbecco's modified Eagle's Medium-Low Glucose (DMEM-LG, GIBCO Invitrogen, Carlsbad, CA, USA) supplemented with L-Glutamine 200 mM, Antibiotic–Antimycotic 10,000 U/mL sodium penicillin, 10,000 μg/mL streptomycin sulfate, 25 μg/mL amphotericin B (GIBCO/Invitrogen Corporation), and 10% fetal bovine serum (GIBCO/Invitrogen Corporation). AUCBSCs' cultures were maintained at 37°C in 5% CO2 and nonadherent cells were removed after 48 h. Medium was changed every other day. The experiments were performed in the fourth and eighth passages of the cultures with approximately 90% confluence.
Immunophenotyping of cultured AUCBSCs by flow cytometry
We analyzed cell-surface expression with a predefined set of protein markers. These assays were performed using commercially available monoclonal antibodies, following the manufacturer's instructions. Briefly, the cells at passage fourth and eighth were harvested by a treatment with 0.25% TrypLE Express (GIBCO Invitrogen), washed with phosphate-buffered saline (PBS) (pH = 7.4), and stained with the selected monoclonal antibodies and incubated in the dark for 30 min at 4°C. Cells were then washed and fixed with 1% paraformaldehyde. The following human antibodies were used: CD14-FITC (clone: M5E2; BD Pharmingen, San Diego, CA, USA), CD29-PE (clone: MAR4; BD Pharmingen), CD31-PE (clone: WM59; BD Pharmingen), CD34-PE (clone: 581; BD Pharmingen), CD44-PE (clone: 515; BD Pharmingen), CD45-PerCP-Cy5 (clone: 2D1; Biosciences, San Jose, CA, USA), CD73-PE (clone: AD2; BD Pharmingen), CD90-APC (clone: 5E10; BD Pharmingen), CD106-FITC (clone: 51-10C9; BD Pharmingen), CD166-PE (clone: 3A6; BD Pharmingen), HLA-DR-PerCP-Cy5 (clone: L243; Biosciences), and CD105-PE (clone: 8E11; Chemicon, Temecula, CA, USA). Cells were analyzed using a FACSARIA flow cytometry equipment (BD Biosciences) and data analyses were performed using FACSDIVA software (BD Biosciences) or Flow Jo Software (TreeStar, Ashland, OR, USA).
In vitro differentiation assays
To evaluate AUCBSCs differentiation capacity, cells (fourth and eighth passages, at 70–80% confluence) were subjected to adipogenic, chondrogenic, and osteogenic differentiation in vitro, according to established protocols (Gang et al., 2004; Jiang et al., 2002).
To promote adipogenic differentiation, subconfluent cells were treated with an Alpha Minimum Essential Medium (α-MEM, GIBCO Invitrogen) supplemented with 10% FBS, 1 μM dexamethazone (ACHE), 100 μg/mL 3-isobutyl-1-methylxanthine (Sigma-Aldrich, St. Louis, MO, USA), 5 μg/mL insulin (Lilly, Indianapolis, IN, USA), and 60 μM indomethacin (Sigma-Aldrich). Adipogenic differentiation was confirmed after 21 days of treatment by intracellular accumulation of lipid-rich vacuoles stainable with Oil Red O stains (Sigma-Aldrich). For the Oil Red O stain, cells were fixed with 4% paraformaldehyde for 30 min, washed, and stained with a working solution of 0.3% Oil Red O for 20 min.
For the chondrogenic differentiation assays, subconfluent cells were treated with an α-MEM (GIBCO-Invitrogen) supplemented with 10% FBS, 50 μg/mL ascorbic acid (Sigma-Aldrich), 10 ng/mL TGF-β (R&D Systems, Indianapolis, IN, USA) and 6.25 μg/mL insulin (Lilly) for 21 days. The chondrogenic differentiation was confirmed by staining with Toluidine blue to indicate extracellular matrix mucopolysaccharides accumulation.
To induce osteogenic differentiation, subconfluent cells were cultured in α-MEM (GIBCO-Invitrogen, Carlsbad, CA) supplemented with 10% FBS, 0.1 mM ACHE, 10 μM β-glycerophosphate (Sigma-Aldrich), and 50 μg/mL ascorbic acid (Sigma-Aldrich) for 21 days. Osteogenesis was confirmed by accumulation of mineralized calcium phosphate assessed by the use of Alizarin Red.
AUCBSCs subjected to none of the differentiation protocols were used as a negative control in the differentiation studies.
Ultrastructural characterization by transmission electron microscopy (TEM)
After 3 weeks, the AUCBSCs samples differentiated into adipocyte-like, chondrocyte-like, osteoblast-like cells, and their respective controls at fourth and eighth passages were also ultrastructurally characterized. The differentiated cells were removed from the plate using Tryple Express (GIBCO-Invitrogen).
The pellets of these cells were fixed directly in 0.5% glutaraldehyde fixative solution. Next, the cells were fixed in 2.5% glutaraldehyde in 0.2 M cacodylate buffer for 2 h at 4°C. Two baths of 15 min in a cacodylate buffer followed this process. Postfixing was performed in 1% osmium tetroxide for 1 h at 4°C followed by another two 15-min baths in the same buffer. For contrast, the material was immersed in a solution of uranyl acetate in acetone for 30 min. After dehydration, the material was embedded in Epon resin diluted in acetone (1:1) and incubated at 4°C with agitation for 24 h. The material was then embedded in pure Epon resin and incubated at 60°C for 72 h until complete polymerization. Semi- and ultrathin sections were obtained with the aid of an ultramicrotome. The semithin sections were stained with Azur II (1%) and Methylene Blue (1%). The Ultrathin sections were placed on copper meshes and contrasted with uranyl acetate and lead citrate. The meshes were analyzed and photographed under a transmission electron microscope (PHILIPS CM100).
QD-labeled AUCBSCs
Primary antibodies CD29 and CD44 usually used as AUCBSCs markers, both in conjunction with the secondary (antimouse IgG) linked to 565-nm nanocrystals QD (Qtracker 565 Cell Labeling Kit, Invitrogen) were used to label of the undifferentiated AUCBSCs according to the manufactor
Flow cytometry analysis of the AUCBSCs immunophenotypic markers set
The markers CD29, CD44, CD73, CD90, and CD105 were also evaluated by flow cytometry in AUCBSCs in the fourth passage labeled with 655-nm nanocrystals QD. The data acquisition also was carried out by FACSARIA flow cytometry equipment (BD Biosciences) and data analysis was performed using FACSDIVA software (BD Biosciences) or Flow Jo Software (TreeStar).
RNA extraction
For RNA extraction of AUCBSCs labeled with 655-nm nanocrystals QD at fourth passage, we used RNeasy® Mini Kit (Uniscience, Qiagen, Chatsworth, CA, USA), following the manufacturers' instructions. DNase treatment (Uniscience, Qiagen) also was performed for elimination of eventual contamination by genomic DNA. Total RNA was quantified by spectrophotometer and its integrity assessed by running the samples on agarose gels.
cDNA synthesis and gene expression evaluation by quantitative RT-PCR
The cDNA was synthesized using 1 μg of total RNA from AUCBSCs samples at fourth passage with SuperScriptTMIII Reverse Transcriptase kit (Invitrogen), under conditions recommended by the manufacturer. PCR primers were designed using Primer Express® Software v2.0 (Applied Biosystems, Bedford, MA, USA) (Table 1).
The Kit QuantiTect SYBR Green PCR (Uniscience, Qiagen) was used to quantify gene expression by qRT-PCR, under conditions recommended by the manufacturer. For normalization of the data, housekeeping gene Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA was used.
Statistical analysis
Gene expression data were analyzed by one-way repeated-measures ANOVA. For both tests values with p < 0.05 were considered statistically significant.
Results
Isolation and culture of adherent cells from UCB
Fifteen UCB units were collected and processed and after 48 h plating MNCs from these samples; different cell types were observed (data not shown). After 20 days we were able to isolate and expand AUCBSCs primary culture from five of the UCB units that corresponds to an isolation success rate of approximately 33% (data not shown).
The ultrastructural analysis of the adherent cells demonstrate the cytological description, that is, a spindle morphology with cytoplasmic extensions (Fig. 1A, B, and E), and oval nuclei (Fig. 1A and B) following cell morphology (Fig. 1C), dispersed chromatin and visible nucleoli (Fig. 1A, D, F, and H). Figure 1H describes irregular morphology with a nucleus and evident nucleolus, suggesting high transcriptional activity. The cytoplasm of these cells showed well-developed rough endoplasmic reticulum, both circular and lamellar (Fig. 1A, C, D, E, F, G, and H), elongated or round mitochondria (Fig. 1A, D, E, and F) with well-defined ridges (Fig. 1G), Golgi complex (Fig. 1F), and large amounts of lipid droplets (Fig. 1A, C, D, E, and H). For all experiments described in this study were used the five established cellular strains.
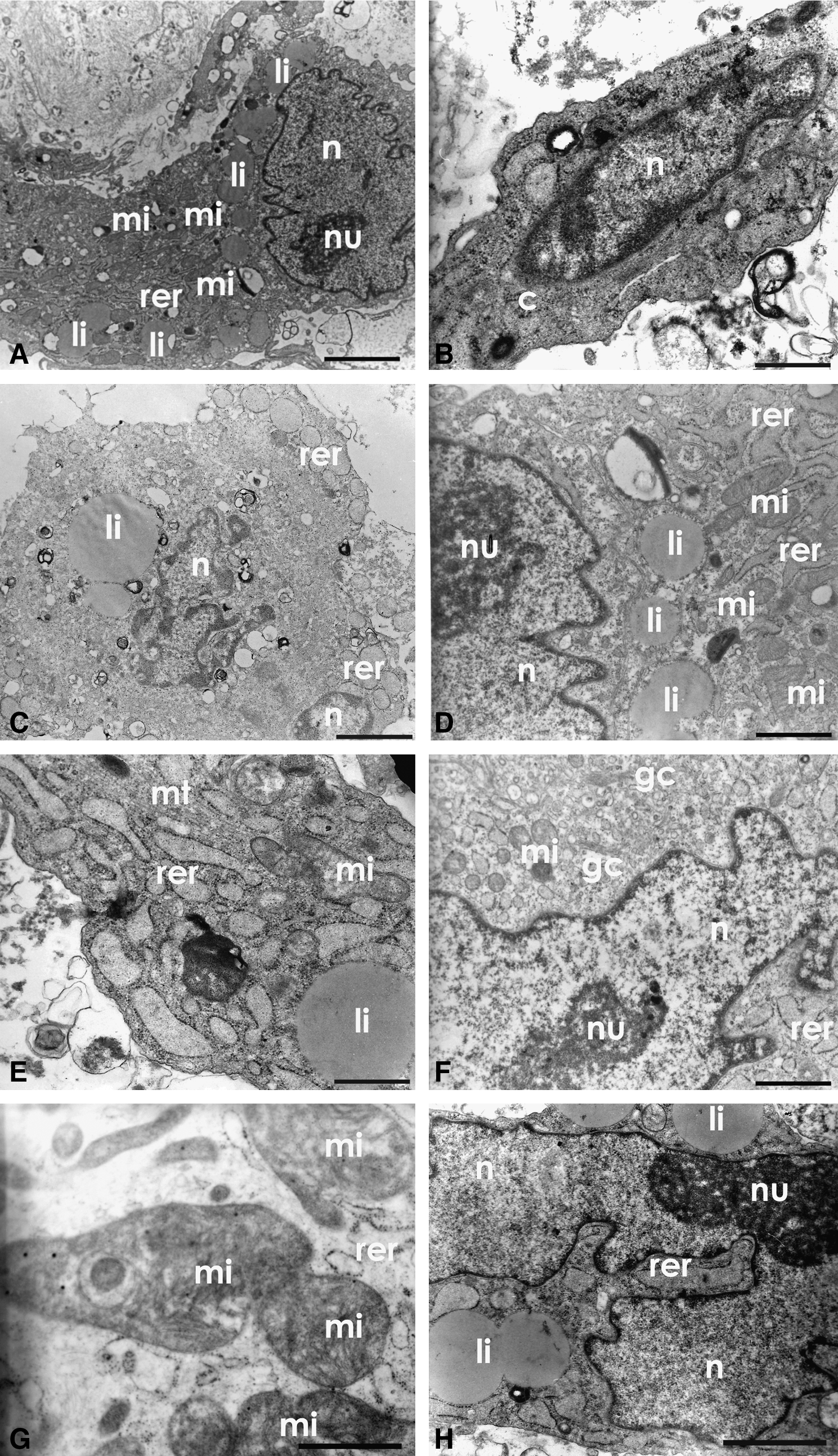
Ultrastructural analysis (TEM) of the AUCBSCs primary culture. (
In vitro differentiation assay, cytochemical, and TEM analysis of AUCBSCs in the fourth passage
During the fourth passage, the differentiation process was confirmed in five cellular strains. After 21 days in culture with adipocyte differentiation medium, the adipocyte-like cells differentiation was confirmed by Oil Red cytochemical test (Fig. 2B). These cells showed marked morphological changes compared to their undifferentiated control (Fig. 2A), that is, the adherent cells differentiated into adipocyte-like cells were shown to be oval, with peripheral basophilic nuclei due to the presence of many lipid droplets, showed in red, as demonstrated by the test (Fig. 2B). The electron micrograph of these cells confirmed the presence of many lipid droplets compressing the nucleus to the cell periphery (Fig. 2E and F). The ultrastructural analysis also described the presence of lipids often associated with mitochondria (Fig. 2E and F), indicating possible production of lipids.
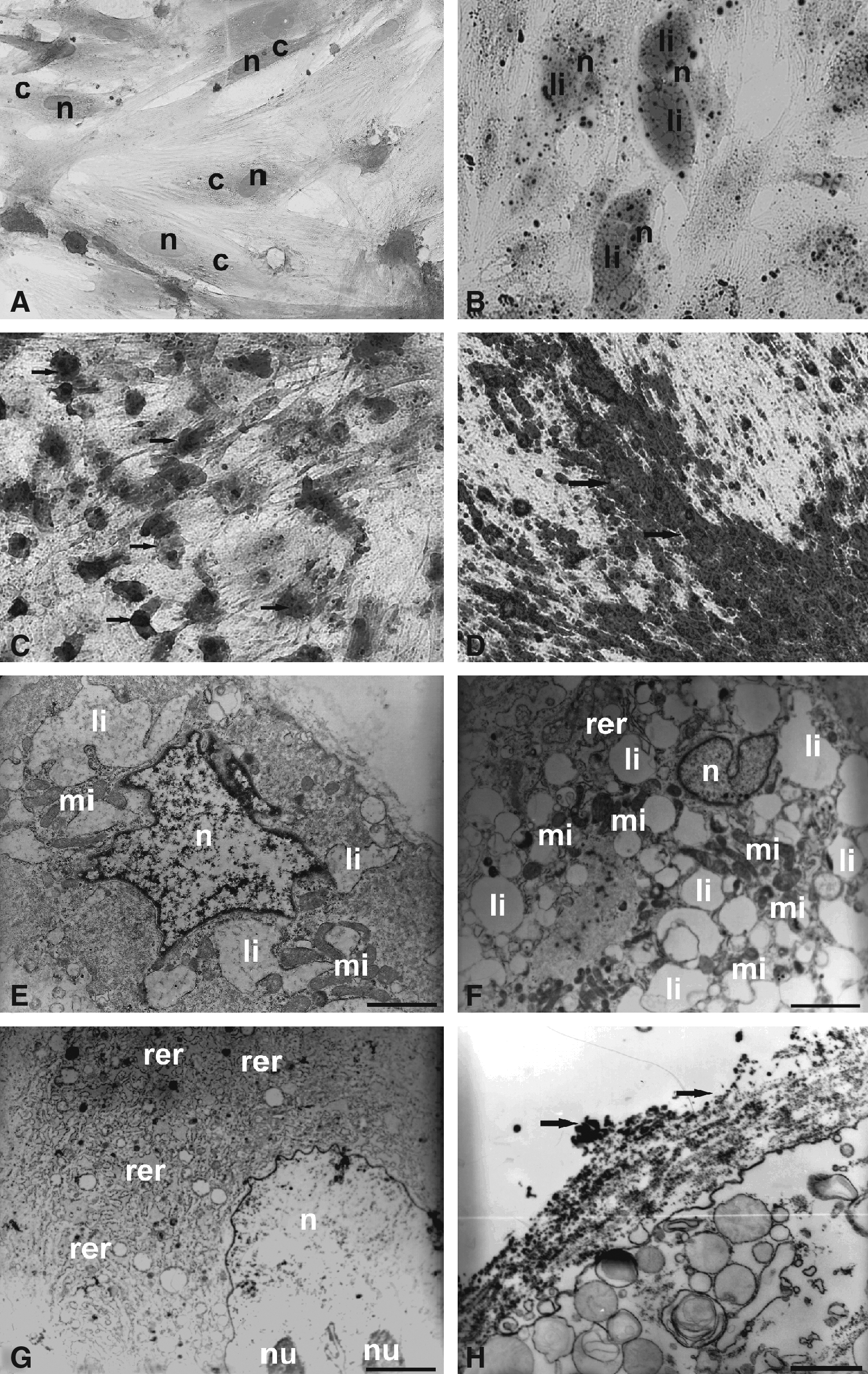
Differentiated cells obtained from AUCBSCs at fourth passage. (
The differentiation of adherent cells in the fourth passage into chondrocyte-like cells, analyzed after 21 days of treatment, showed changes from their previously fibroblastic morphology (Fig. 2A) into strongly basophilic round cells (Fig. 2C). The ultrastructural analysis also suggests the initiation of a chondrogenic mechanism, based on the morphology of the cells, showing rounded cells with evident nucleoli and the presence of a well-developed rough endoplasmic reticulum in the cytoplasm (Fig. 2C and G).
The metachromasia found in cytochemical analysis of these cells may be related to the possible presence of cartilaginous matrix acids elements, such as glycosaminoglycans or mucopolysaccharides (Fig. 2C).
The differentiation of adherent cells in the fourth passage into osteoblast-like cells was also confirmed after 21 days of treatment showing strong cytochemical marking of Alizarin Red, which indicates the presence of calcium deposits (Fig. 2D). This process was ultrastructurally characterized by the presence of calcium in the extracellular matrix. In this case we observed a smaller quantity of calcium deposits due to the enzymatic treatment with tripsin to remove cells from culture flask for these analyses (Fig. 2H).
Our results confirmed the mesenchymal nature of the isolated cells in the fourth passage and their multipotency. These results were essential as a control prior to the study using nanocrystals, and also to reach a sufficient number of cells in this early passage for the experiments accomplishment.
TEM analysis of AUCBSCs in the eighth passage
AUCBSCs in the eighth passage were also submitted to the same treatment for differentiation in adipocyte-, osteoblast-, and chondrocyte-like cells, but the cells died before the experiment terminus (data not shown). These results show that these cells present loss of their multipotenciality.
The data above corroborate electronic micrographs of these cells showing the frequent presence of concentric myelin figures (Fig. 3B and D) and intense cytoplasmic vacuolization (Fig. 3A–D). There is also the vesiculation of rough endoplasmic reticulum (Fig. 3A, C, D, and E), disintegration of mitochondrial crests (Fig. 3A), as well as the presence of several apoptotic bodies (Fig. 3F) and the intense chromatin condensation (Fig. 3G and H).
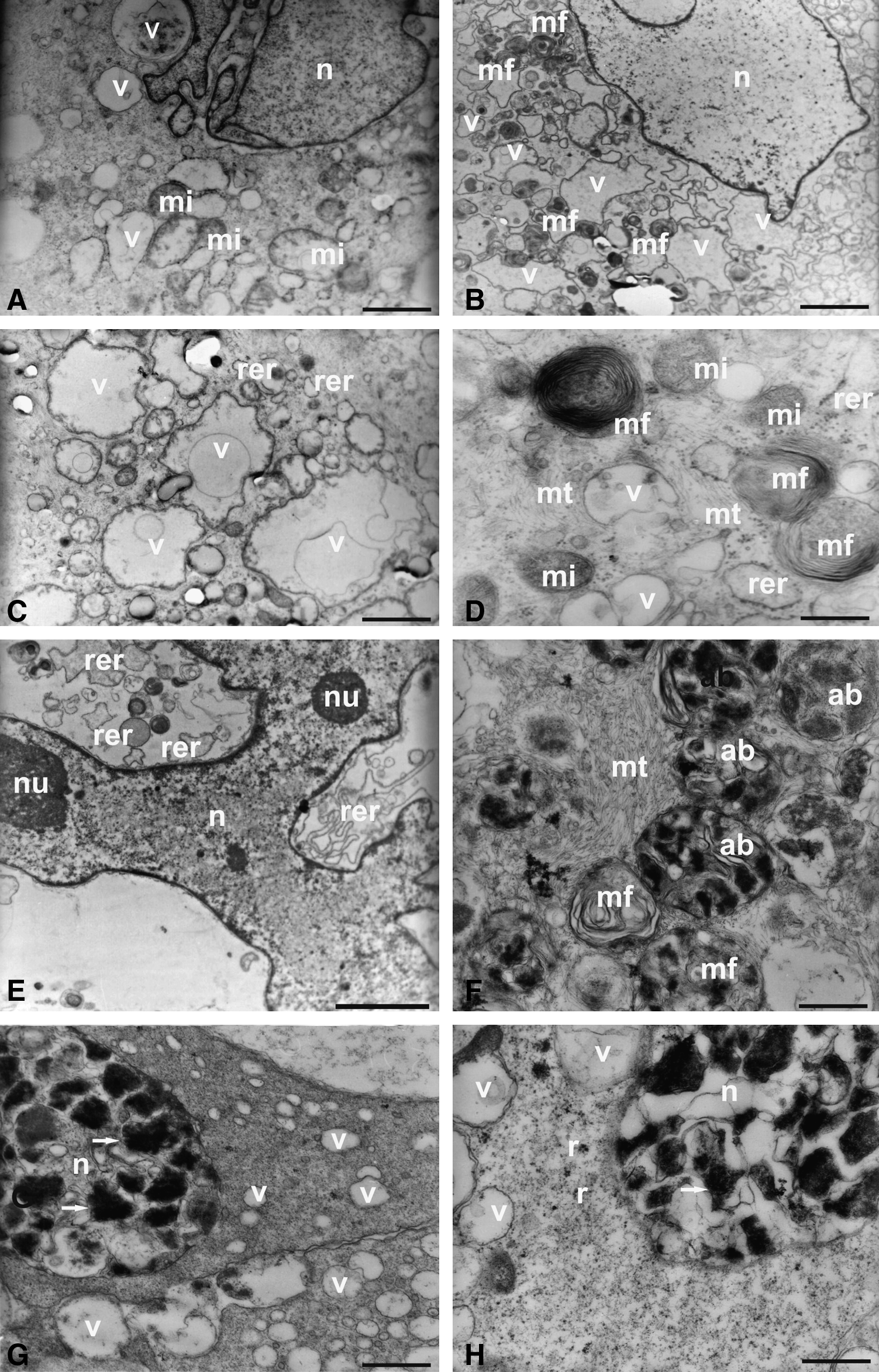
Ultrastructural analysis (TEM) of the AUCBSCs at eighth passage. (
These results show that the UCB-MSC in the eighth passage are not viable to be used in experiments of cellular differentiation due to loss of their multipotenciality when compared with UCB-MSC in the fourth passage. Thus, all experiments were performed with the cells in the fourth passage.
CD29- and CD44-QD labeling of AUCBSCs
Undifferentiated AUCBSCs in the fourth passage when labeled with CD29- and CD44-QD conjugated with the secondary (antimouse IgG) linked to 565-nm nanocrystals QD showed stronger expression of CD29 (Fig. 4A) and CD44 (Fig. 4C) monoclonal antibodies linked to nanocrystals, by membrane fluorescence when compared with the controls from the immunofluorescence reactions (Fig. 4A′ and C′). Ultrastructurally, the nanocrystals were detectable by their electrondensity. The electronic micrographs show the presence of nanocrystals in the plasmatic membrane, cytoplasm, and in the nucleus (Fig. 4B–F).
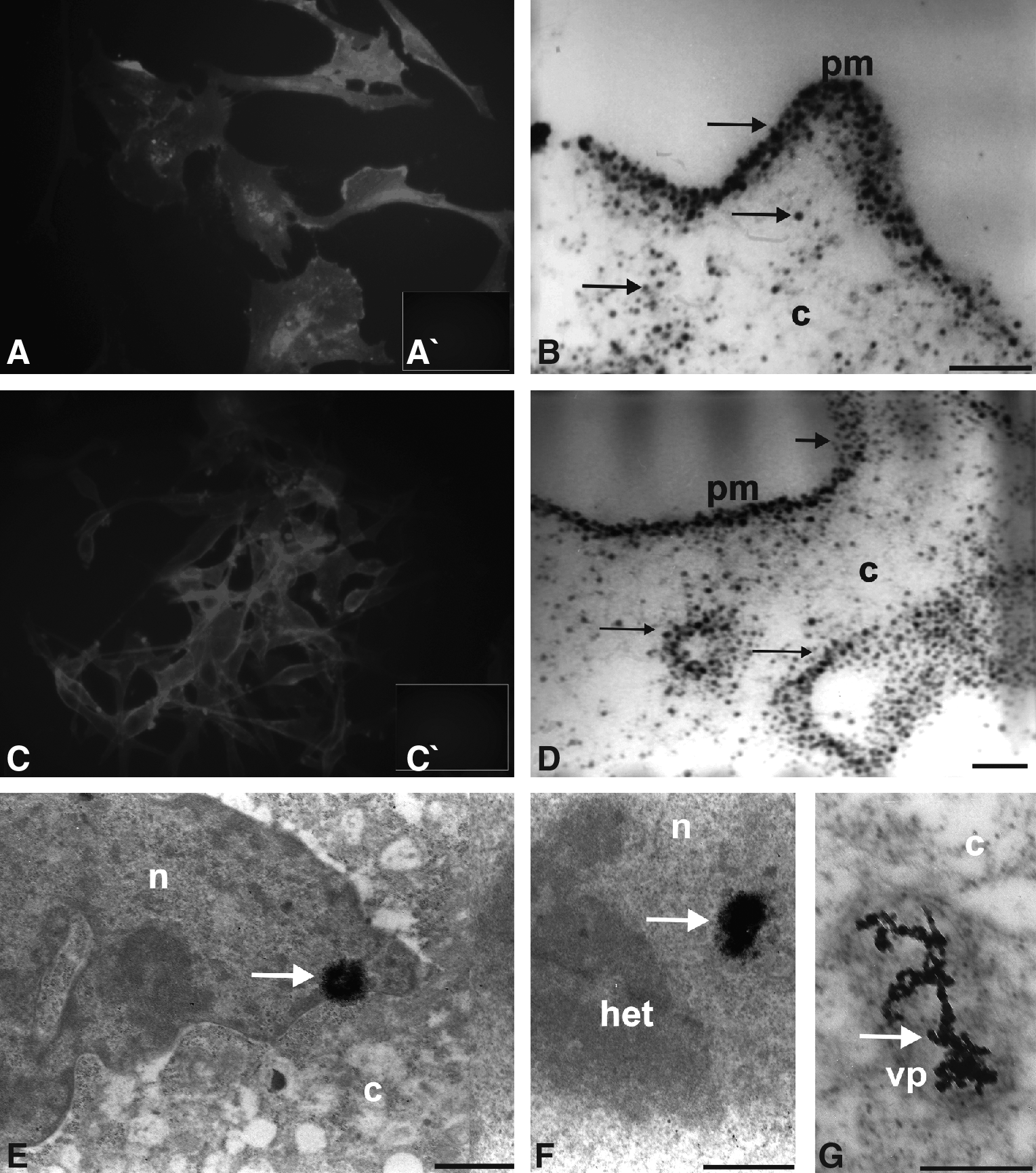
Immunofluorescence and TEM analysis of the undifferentiated AUCBSCs at fourth passage marked by CD29- and CD44-QD conjugated with the secondary (antimouse IgG) linked to 565 nm nanocrystals QD. (
The nanocrystals immunolocalization is often scattered in the cytoplasm, and concentrated in pinocytics vesicles (Fig. 4G). These pinocytics vesicles are likely to be obtained by the process of endocytosis mediated by receptors, probably corresponding to the specific markings found in the reactions of the fluorescence microscopy analysis (Fig. 4G).
Undifferentiated AUCBSCs in the fourth passage were also labeled in the culture with nonlinked nanocrystals, 655-nm nanocrystals QD, which were internalized by the cells, showing a diffuse pattern of labeling, excluding the nucleus (Fig. 5A). It was observed that this process did not influence the ability of AUCBSCs differentiation process into adipocytes-, chondrocytes-, and osteoblasts-like cells, and that these cells, even while differentiated, retained the nanocrystals (Fig. 5H and I).

Fluorescence labeling and TEM analysis of the undifferentiated and differentiated AUCBSCs at fourth passage marked with 655-nm nanocrystals QDs. (
The ultrastructural data of AUCBSCs in the fourth passage differentiated into adipocyte-like cells, showed the presence of nonlinked nanocrystals, such as electrondense groups, preferably inserted in vesicles (Fig. 5C), emphasizing that the process of differentiation was not altered by this finding, which is shown by Oil Red cytochemistry (Fig. 5B).
The AUCBSCs in the fourth passage differentiated into chondrocyte-like cells showed the nanocrystals associated with rough endoplasmic reticulum, which are well developed in these cells (Fig. 5E). Again, the process of differentiation, as evidenced by the toluidine blue test was not compromised by the labeling (Fig. 5D).
Ultramorphologically the AUCBSCs in the fourth passage differentiated into osteoblast-like cells showed the presence of nanocrystals dispersed in the cell cytoplasm, and expression of an extracellular calcium matrix (Fig. 5G). The occurrence of calcium accumulation was confirmed by the Alizarin Red cytochemical assay (Fig. 5F).
Immunophenotypic analysis of the QD nonlabeled and QD labeled AUCBSCs by flow cytometry
Nanocrystals (655 nm) QDs labeled and nonlabeled AUCBSCs' immunophenotypic set of markers (CD29, CD44, CD73, CD90, and CD105) were assessed by flow cytometry. We observed that five established strains of AUCBSCs in the fourth passage express all MSC immunophenotypic set of markers, highlighting CD90 expression that seems to be an important marker for undifferentiated AUCBSC state (Fig. 6).

Expression levels of AUCBSCs-related proteins expressed by undifferentiated AUCBSCs at fourth passage labeled with 655-nm nanocrystals QDs by flow cytometry. CD29: nonlabeled AUCBSCs; CD29 QDs: AUCBSCs labeled with QDs; CD44: AUCBSCs nonlabeled; CD44 QDs: AUCBSCs labeled with QDs; CD73: AUCBSCs nonlabeled; CD73 QDs: AUCBSCs labeled with QDs; CD90: AUCBSCs nonlabeled; CD90 QDs: AUCBSCs labeled with QDs; CD105: AUCBSCs nonlabeled; CD105 QDs: AUCBSCs labeled with QDs (n = 5; five AUCBSCs strains). *All the test values p < 0.05 were considered statistically significant.
These results show that AUCBSCs labeled with 655-nm nanocrystals QDs did not change their characteristics.
Immunophenotypic markers gene expression in QDs labeled and nonlabeled AUCBSCs by qRT-PCR
We analyzed the gene expression of as CD29, CD44, CD73, CD90, and CD105 passage. We observed that the labeling of undifferentiated AUCBSCs with 655-nm nanocrystals QD does not affect the expression of the characteristic immunophenotypic markers (Fig. 7). As described before, the expression of CD90 in undifferentiated cells was very marked. For normalization of the data, housekeeping gene Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA was used. These data corroborate the observed results in the flow cytometry.
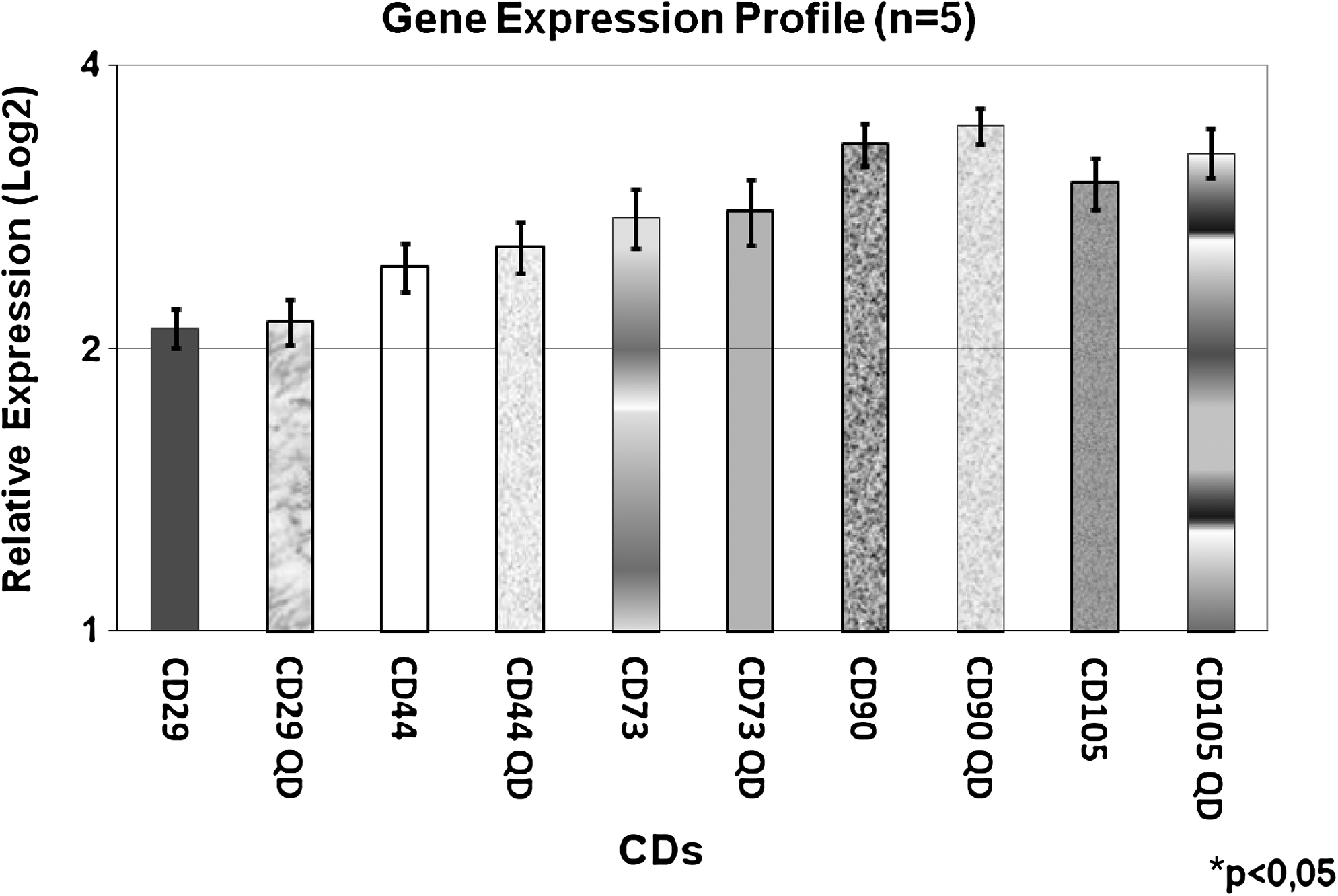
Gene expression of undifferentiated AUCBSCs at fourth passage labeled with 655-nm nanocrystals QDs by qRT-PCR. CD29: nonlabeled AUCBSCs; CD29 QDs: AUCBSCs labeled with QDs; CD44: nonlabeled AUCBSCs; CD44 QDs: AUCBSCs labeled with QDs; CD73: AUCBSCs nonlabeled; CD73 QDs: AUCBSCs labeled with QDs; CD90: AUCBSCs nonlabeled; CD90 QDs: AUCBSCs labeled with QDs; CD105: AUCBSCs nonlabeled; CD105 QDs: AUCBSCs labeled with QDs (n = 5; five AUCBSCs strains). *All the test values p < 0.05 were considered statistically significant.
Discussion
Some of the most promising research in the field of regenerative medicine has focused on the use of MSCs. These cells, by definition, are undifferentiated cells with significant proliferative capabilities. MSCs have multilineage differentiation potential and are a valuable source of progenitors cells (Bielby et al., 2007; Bobis et al., 2006; Caplan and Bruder, 2001; Jin et al., 2009; Kolf et al., 2007; Meirelles et al., 2008).
There are some studies that demonstrated better cellular viability in earlier than later passages of MSCs. However, none of them showed the best passage through morphological and ultrastructural characterization or suggested the ideal passage for further studies with these cells (Baxter et al., 2004; Itahana et al., 2001; Kestendjieva et al., 2008; Sethe et al., 2006).
Here, we preferred to name MSCs as AUCBSCs because their adherence to the culture flask, proliferative, and cellular differentiation capacity. In this study we demonstrated for the first time through ultrastructural characterization that AUCBSCs culture in the fourth passage maintains the fibroblast morphology and differentiation capacity into mesodermal lineages. In addition, these cells characteristics are altered by long periods of cultivation as observed in the eighth passage. Through TEM and cytochemical analysis, we demonstrate a set of morphological findings that characterize AUCBSCs in the fourth passage cells as being viable.
Senescent cells are defined as cells with a less replicative capacity as well as with altered morphologic characteristics (Itahana et al., 2001; Sethe et al., 2006). The AUCBSCs ultrastructural analysis in the eighth passage showed concentric myelin figures, intense cytoplasmic vacuolization, vesiculation of the rough endoplasmic reticulum, disintegration of mitochondrial crests and the presence of several apoptotic bodies and intense condensation of chromatin. These finds seems to be related to cellular senescence leading to reduced potential of differentiation (data not shown).
Thus, this work suggests that AUCBSCs from early passages should be used. We also suggest that researchers should certify that the AUCBSCs used in their studies are healthy and are giving rise to reliable and reproducible data.
The labeling of cells with QDs has been recognized as an important alternative method of tracking cells. However, the possible cytotoxic effects of QDs were not extensively evaluated. To date, little is known about the potential cytotoxic effects of QD labeling on AUCBSCs. Muller-Borer et al. (2007) showed in a cardiac microenvironment that functional integration of rat MSCs labeled with QD was resistant to chemical and metabolic degradation, with long-term photo-stability and minimal cytotoxic effect.
Recent findings using confocal microscopy and TEM analysis showed the presence of QDs aggregates in endosomal vesicles localized in the peri-nuclear region in labeled MSCs (Rosen et al., 2007; Seleverstov et al., 2006). These works showed that both low and high concentrations of QDs appear to be cyto-compatible with the MSCs and capable of labeling proliferating cells in vitro.
Here, we investigated how the labeling with QDs affects AUCBSCs characteristics. We demonstrated that this approach is feasible and noncytotoxic to the AUCBSCs in the fourth passage. For the first time we demonstrate that the labeling of AUCBSCs with QDs does not change the expression of immunophenotypic markers set usually expressed in AUCBSCs.
We decided to evaluate the AUCBSCs differentiation only in the cells labeled with unconjugated QD (655 nm), because this labeling method seems to be more stable, dispenses specific markers, and maybe useful to a variety of cell types.
Our results suggest that QD labeled AUCBSCs can be used in trafficking experiments, even 21 days after labeling. AUCBSCs differentiated in the three mesoderm lineages still presenting QDs aggregates in pinocytics vesicles and in the cytoplasm as shown in the micrographs. In fact, Ballou and coworkers (2004) showed that QDs' cellular fluorescent labeling is stable and remain in vivo for at least 4 months.
More important, we demonstrated that AUCBSCs labeled with QDs, besides retaining the labeling for more than 21 days also maintain unaltered their in vitro differentiation potential.
Summarizing, in this study for the first time we characterized AUCBSCs ultrastructurally and defined that these cells at later passages should not be used in cellular experimentation, because they have their morphology and differentiation potential compromised. We also demonstrated that AUCBSCs labeled with QDs maintain their characteristics and viability unaltered.
Additionally, this study will contribute as a source of information for further in vivo studies.
Footnotes
Acknowledgments
We thank all the members from the Banco Público de Sangue de Cordão Umbilical do Hospital Israelita Albert Einstein who assisted us in the umbilical cord blood samples collection. We are also grateful to Laboratório de Microscopia Eletrônica, Departamento de Biologia, UNESP de Rio Claro, SP, Brasil, and Antonio T. Yabuki and Morita Iamonte for the technical support. This work was financed by Instituto de Ensino e Pesquisa Albert Einstein, Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Author Disclosure Statement
The authors declare no conflict of interest.