
Abstract
Abstract
Recent findings indicate that bone marrow mesenchymal stem cells (BMSCs) participate in the process of neovascularization in response to repair to injury and are involved in postinfarction myocardial repair. It is unclear what special characteristics the vascular progenitors of bone marrow origin has. CXCR4+ stem/progenitor cells mobilized to the infarct area and improved the myocardial repair. In present study, we aimed to determine whether CXCR4+BMSCs contribute to the angiogenic capacity in vitro and in vivo. CXCR4+BMSCs were separated by using paramagnetic microbeads and cultured. RT-PCR and FACS analysis confirmed the gene expression phenotype. The uptake of acetylated low density lipoprotein (acLDL) and the tube formation evaluated the function of CXCR4+BMSCs. The effect of CXCR4+BMSCs transplantation on neovascularization was investigated in a murine model hindlimb ischemia. After induced by VEGF, CXCR4+BMSCs expressed the endothelial cells (ECs) phenotype. The expression of EC markers, PECAM-1, and von Willebrand factor (vWF) increased significantly at both the mRNA and protein levels. In addition, CXCR4+BMSCs enhanced the uptakes of Dil-acLDL and form capillary-like tubes in vitro. In vivo the local transfer of CXCR4+BMSCs increased neovascularization in ischemic hindlimb. These results demonstrate that CXCR4+BMSCs differentiate into ECs and contribute to neovascularization in the vascular lesion,, which indicate the important therapeutic implications for cardiovascular diseases and a new cell source for cell-based vascular engineering and repair in the future.
Introduction
The transplantation of culture-expanded endothelial progenitor cells (EPCs) or bone mesenchymal stem cells (BMSCs) successfully increases collateral vessel formation and promote therapeutic neovascularization in both myocardial infarction and ischemic hindlimb models (Kocher et al., 2001; Shintani et al., 2001). Mechanistically, these cells can induce angiogenesis by incorporation into vascular structures and repair the infarction and improve function and survival (Kawamoto et al., 2001; Miller et al., 2008). These findings indicated that autologous EPC or BMSC transplantation improved the neovascularization in patients with coronary and peripheral artery disease (Lian et al., 2008; Tateishi-Yuyama et al., 2002).
The application of cell-based therapy for the treatment of cardiovascular disease remains in its preliminary phase. Assmus et al. (2002) reported that transplantation of progenitor cells derived from bone marrow or circulating blood significantly increased left ventricular ejection fraction and improved myocardial viability in the infarct zone. Wollert et al. (2004) reported that transfer of autologous bone marrow cells promotes improvement of left ventricular systolic function in patients after acute myocardial infarction. However, recent data indicated that the cell therapeutic success was determined by functional properties of transplanted cells, and encounters significant challenges in isolation techniques, scalability, reproducibility, and ease of clinical application (Kawamoto et al., 2001; Real et al., 2008). Further elucidation of these mechanisms may have implications for the understanding of cell based therapeutic angiogenesis.
It was recently shown that CXCR4+ cells increases in patients with acute myocardial infarction (Cheng et al., 2008; Wojakowski et al., 2006). Subcutaneous injection of granulocytecolony stimulating factor (G-CSF) increases the number of functional CXCR4 receptors on the cell membrane and recruitment of CXCR4+ cells (Kollet et al., 2002; Misao et al., 2006). The delivery of modified MSCs expressing CXCR4 improved the postinfarction myocardial repair (Cheng et al., 2008; Morimoto et al., 2007). In addition, vascular endothelial growth factor (VEGF) plays a critical role in proliferation and differentiation of EPCs.
Given the critical role of CXCR4+ progenitor cell, we examined the effects of CXCR4+ BMSCs on the neovascularization in vitro and in vivo. The purpose of the present study was: (1) to determine the expression changes of ECs marker in CXCR4+BMSCs induced by vascular endothelial growth factor (VEGF); (2) to examine the function and ultrastructure of CXCR4+BMSCs; and (3) to evaluate the neovascularization and tissue repair by delivering CXCR4+BMSCs to ischemic hindlimb model.
Material and Methods
Materials
Anti-CXCR4, anti-PECAM-1, anti-vWF, and anti-Flk antibody were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA); anti-GFP antibody was from Abcam (Cambridge, UK). All secondary antibodies were from Proteintech Group (Chicago, IL, USA). DiI-acLDL and Matrigel were purchased from Sigma (St. Louis, MO, USA).
Isolation and culture of CXCR4+BMSCs
BMSCs were separated as previously described (Kim et al., 2005). Bone marrow cells were separated from 2-month-old C57/BL6 female mouse tibias and femurs, plated on culture dishes coated with rat vitronectin and 0.5% gelatin (Sigma) and maintained in endothelial basal medium (EBM; Clonetics, Warrendale, PA, USA) supplemented with EGM SingleQuots and 10% fetal bovine serum (FBS). After cultured for 24 h, nonadherent cells were removed by medium change and fresh medium added to the cells. Medium was changed every 3 days. At passage 3, the cells were harvested and washed with phosphate-buffered saline (PBS) and were preincubated with anti-CXCR4 antibody (10 μg/mL) in FBS and subsequently were preincubated with paramagnetic microbeads coated with antirabbit IgG antibody. CXCR4+BMSCs were separated using a MINI-MACS device (Miltenyi Biotech, Auburn, CA, USA), and purity assessed by FACS analysis was >95%. CXCR4+BMSCs were characterized by immunofluorescence staining; over 98% of cells expressed CD29, CD71, CD90, CD106, and CD117, and did not express the hematopoietic marker CD45 or CD34.
FACS analysis of CXCR4+BMSCs
FACS analysis was performed as previously described (Eggermann et al., 2003). Briefly, CXCR4+BMSCs were cultured to confluence and were treated with 0, 10, and 20 ng/mL VEGF for 24 h. Cells were collected with trypsin/EDTA and incubated with anti-PECAM-1 antibody or anti-vWF followed by PE conjugated with the corresponding secondary antibody. Isotype-identical antibodies served as controls to exclude nonspecific binding. Quantitative analysis was performed using a FACStar flow cytometer and CellQuest software (Becton Dickinson, San Jose, CA, USA).
Quantitative real-time RT-PCR
We performed the real-time RT-PCR as previously described (Hoffmann et al., 2005). After CXCR4+BMSCs were treated with 0, 10, or 20 ng/mL VEGF for 24 h, total cellular RNA was extracted. RNA was reverse-transcribed into cDNA. Real-time quantitative RT-PCR primers were designed by Beacon Designer 2.1 software. The primer sequence used for PECAM-1 was GAGAAGAGCAGCCGATTCCT (forward) and AACCTCCTTTCACCCCCC (reverse). The primer sequence used for vWF was TGTACCATGAGGTTCTCAATGC (forward) and TTATTGTGGGCTCAGAAGGG (reverse). The mRNA levels were normalized by using GAPDH as a housekeeping gene (forward: CCAATCAGCTTGGGCTAGAG; reverse: CCTGGGAAAGGTGTCCTGTA) and compared with levels in the mouse universal gene. This relative value of target genes to endogenous reference is described as the fold of GAPDH = 2–ΔCt
The uptakes of DiI-labeled acLDL in CXCR4+BMSCs
The acetylation of LDL was performed as described (Basu et al., 1976). CXCR4+BMSCs were treated with 20 ng/mL VEGF for 24 h and were washed twice with ice-cold PBS. Then cells were incubated with 10 μg/mL DiI-labeled acLDL for 2 h at 4°C. CXCR4+BMSCs were washed twice with PBS at 4°C and complete media prewarmed to 37°C was added. The cells were incubated at 37°C for 24 h. DiI-labeled acLDL-treated CXCR4+BMSCs were fixed with formalin for the detection of the uptakes of DiI-labeled acLDL under fluorescence microscopy equipped with a digital camera.
In vitro capillary morphogenesis assay
Matrigel (200 μL at a concentration of 5 mg/mL) was pipetted into 24-well and polymerized at 37°C for 30 min to 1 h as previous descibed (Lee et al., 1999). CXCR4+BMSCs were treated with VEGF 20 ng/mL for 1 h in DMEM with 1% FBS, and 400 μL of CXCR4+BMSCs were plated (3 × 104 cells/mL). After 3 and 6 h, plates were photographed. Angiogenesis was evaluated by counting the total length of the tubu-like structures, using ImageJ software according to the manufacturer's instructions. Six to nine photographic fields from three plates were scanned for each point and counted the length of the tubular structures.
Transmission electron microscopy
Ultrastructural studies were performed as previous described (Houwerzijl et al., 2004). For transmission electron microscopic observation, the CXCR4+BMSCs on the filters were fixed with 2.5% glutaraldehyde for 4 h, followed by 1% OsO4 for 1 h. The specimens were dehydrated with a graded ethanol series and isoamyl acetate. They were desiccated by the critical-point method using CO2 and were coated with metal in an IB-3-type ion coater (EIKO Engineering Co Ltd., Japan). Ultrathin sections were cut with an ultramicrotome (MT-7000, Research and Manufacturing) and examined with a FEI Tecnai G2 scanning electron microscope at an accelerating potential of 80 kV after being counterstained with uranyl acetate and lead citrate.
Mouse hindlimb ischemia model and cell delivery
In the experiment, CXCR4+BMSCs and total BMSCs were stably transfected with plasmid containing green fluorescent protein (GFP) gene to trace these cells in vivo. All animal experiments were performed according to protocols approved by the Institutional Committee for Use and Care of Laboratory Animal. Briefly, 6-week-old C57BL/6J female mice were anesthetized with 50 mg/kg intraperitoneal pentobarbital and a unilateral femoral artery was removed to induce muscle ischemia as previously described (Yamahara et al., 2003; Yang et al., 1995). CXCR4+BMSCs (5 × 105) and total BMSCs treated with 20 ng/mL for 24 h in 100 μL medium were administered systemically through intraventricular injection to six mice 48 h after surgery.
Morphometric analysis of lesion formation and the neo-capillaries detection
Four weeks after unilateral femoral artery excision, mice were sacrificed and the calf muscle and tibial bone of either an ischemic hindlimb or a nonischemic hindlinb were harvested, frozen in liquid nitrogen, and prepared for H&E staining or used for detection of cell markers by immunoreactive staining. For GFP-fluorescence detection, 10-μm-thick histological sections were prepared from snap-frozen tissue samples and directly examined under fluorescence microscopy. For detection of endothelial cell markers, the immunochemical staining was used. The procedure was similar to that described previously (Mehes et al., 2001). The specimens were incubated with the anti-GFP or anti-PECAM-1 antibody in the consecution section for 90 min then incubated with secondary antibody for 1 h. The quantification of capillary density was counted as described previously (Bonaros et al., 2006).
Statistical analysis
All results were expressed as mean ± SEM of at least three independent experiments. Statistical significance was evaluated using the unpaired Student t-test for comparisons between two means. A probability value of p < 0.05 was considered significant.
Results
Characterize of CXCR4+BMSCs
The number and purity of transplanted cells determine their potential as cell-based therapy in clinical medicine. To produce high purity transplanted cells from BMSCs, CXCR4+BMSCs were isolated from total BMSCs and treated with 20 ng/mL VEGF in DMEM for 3 days, subcultured with a ratio of 1 to 3 every 2 to 3 days. Morphologically, total BMSCs displayed clusters in an undifferentiated status in our culture conditions, whereas CXCR4+BMSCs displayed a monolayer in culture. In addition, CXCR4+BMSCs grew with a round, long, fiscal, spindle-shaped or whirlpool-shaped appearance, and did not display typical “cobblestone” morphology such as HUVECs (Fig. 1A).
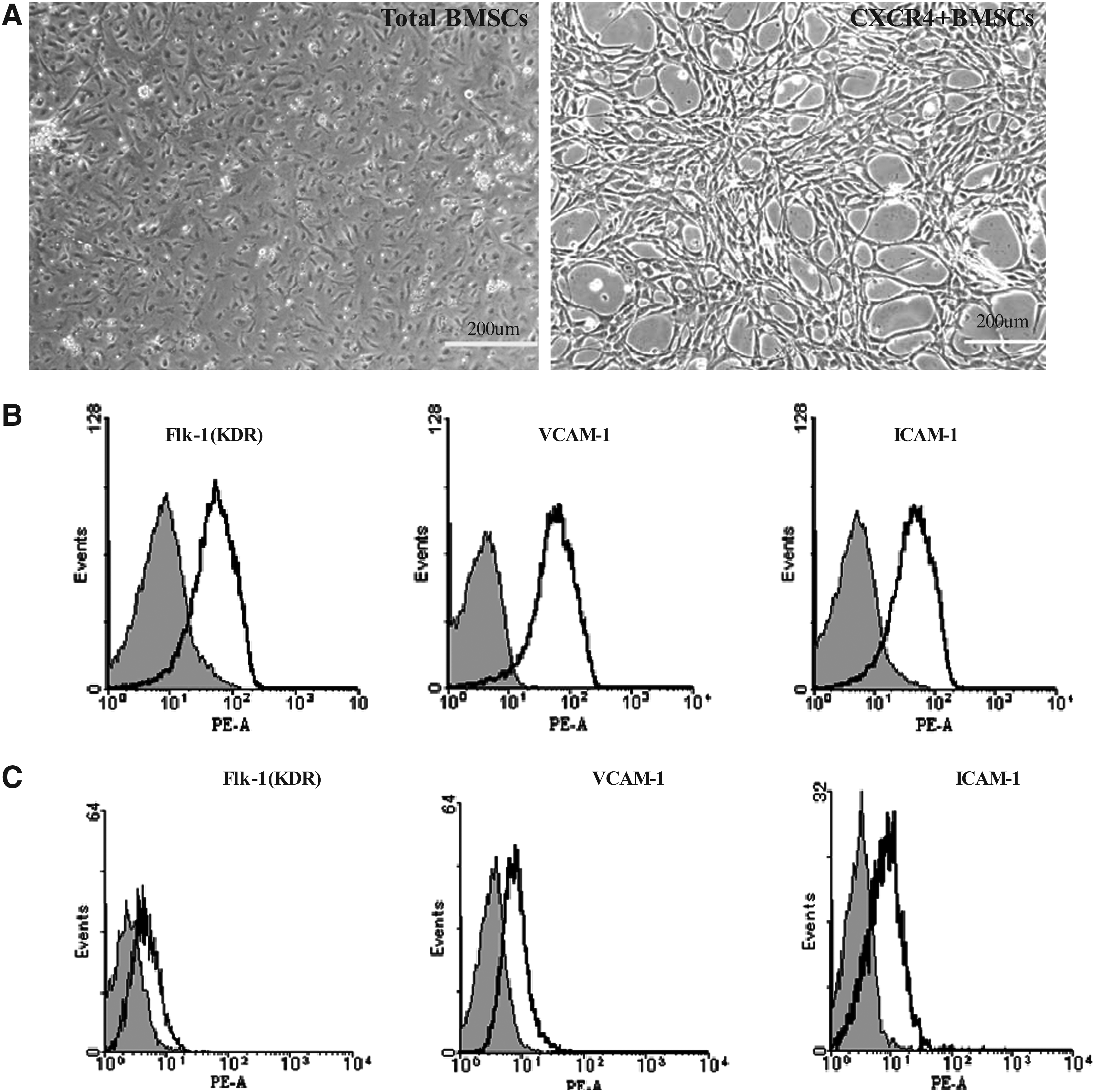
Characteristics of CXCR4+BMSCs. (
We investigated whether the isolated CXCR4+BMSCs were positive for EC linage-specific markers, these CXCR4+BMSCs were detected for Flk-1(KDR), ICAM-1, and VCAM-1 by FACS analysis. As shown in Figure 1B, the expression of marker gene was upregulated in CXCR4+BMSCs induced by VEGF. However, total BMSCs did not show the expression for EC markers (Fig. 1C).
VEGF induces PECAM-1 and vWF expression in CXCR4+BMSCs
CXCR4+BMSCs were exposed to 10 and 20 ng/mL VEGF for 24 h, and real-time PCR was used to quantify the mRNA level of PECAM-1 (CD31), a glycoprotein localized to the endothelial cell junction where it forms Ca2+-independent cell–cell adhesions and used as a mature endothelial cell linage-specific marker (Jackson et al., 2000). Compared with controls, PECAM-1 mRNA expression was significantly upregulated 23 ± 2- and 55 ± 4-fold after 10 and 20 ng/mL VEGF treatment for 24 h, respectively (p < 0.01, Fig. 2A). The cellular protein level of PECAM-1 was determined by flow cytometric analysis. Flow cytometric analysis demonstrated that the PECAM-1 protein expression increased to 7.25 ± 2.43% and 48.42 ± 7.69% after 10 and 20 ng/mL VEGF treatment for 24 h, respectively (p < 0.05, Fig. 2B), compared with control groups.
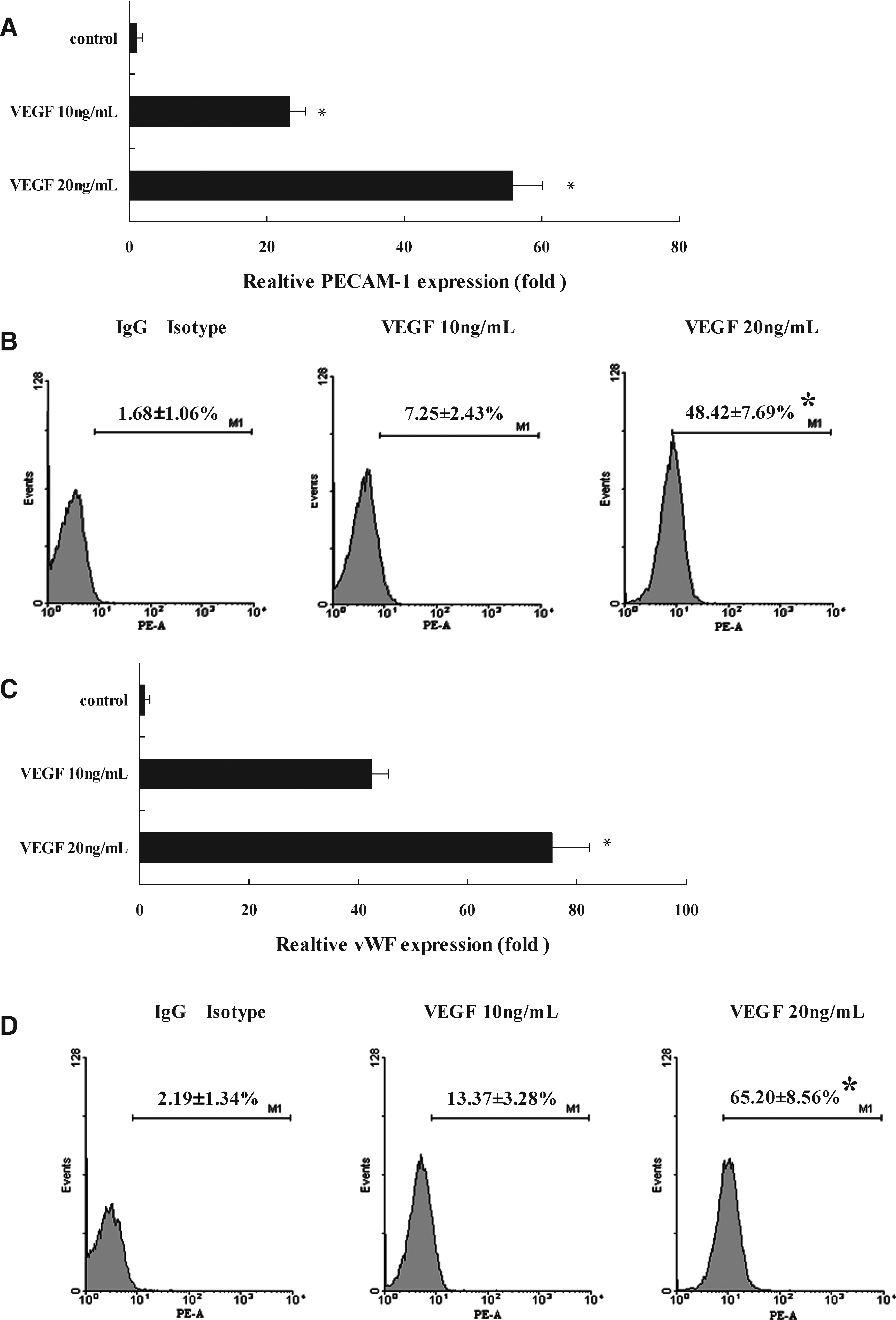
The expression change of VEGF on PECAM-1 and vWF mRNA and protein levels in CXCR4+BMSCs. (
The expression level of vWF, a glycoprotein of protomeric subunits derived from endothelial cells that promoted platelet adhesion (Galbusera et al., 1997), were examined in CXCR4+BMSCs. The mRNA levels of vWF were significantly upregulated 42 ± 3- and 75 ± 6-fold after 10 and 20 ng/mL VEGF treatment for 24 h, respectively (p < 0.01, Fig. 2C), ompared with control groups. Flow cytometric analysis determined the cellular vWF protein level. As shown in Figure 2D, the protein expression of vWF was markedly upregulated 13.37 ± 3.28% and 65.20 ± 8.56% (p < 0.05), respectively when CXCR4+BMSCs were exposed to 10 and 20 ng/mL VEGF for 24 h. These results indicate VEGF induces the expression of PECAM-1 and vWF at both the mRNA and protein levels in CXCR4+BMSCs.
The acLDL uptake and the ultrastructure in CXCR4+BMSCs
The uptakes of DiI-labeled ac-LDL are specific functional markers for ECs in vitro (Eggermann et al., 2003). To further verify whether VEGF induces an endothelial-like phenotype in CXCR4+BMSCs, we measured the uptake of DiI-acLDL by CXCR4+BMSCs under VEGF treatment. Immunofluorescence analysis revealed that CXCR4+BMSCs were positive for DiI-acLDL uptake (Fig. 3A). These results demonstrated that CXCR4+BMSCs uptake DiI-acLDL and have the functional markers of ECs.

The uptake of acLDL and the ultrastructure of CXCR4+BMSCs. (
To understand whether CXCR4+BMSCs have the ultrastructure of ECs, we observed the ultrastructure of CXCR4+BMSCs in electron microscopy. As shown in Figure 3B, CXCR4+BMSCs exhibited elongated and indented nuclei and a distorted cytoplasmic membrane with a few short cytoplasmic projections, which were very similar to ECs in morphologically. In addition, some electron-dense structure appeared in the cytoplasm and might be Weibel-Palade bodies.
The tube formation and the junction of CXCR4+BMSCs
The tube formation is a specific functional marker for ECs in vitro. Capillary-like tube formation on Matrigel was also enhanced in VEGF-treatment CXCR4+BMSCs. Cells displayed increased formation of a tube-like network when examined microscopically at 12 h (Fig. 4A) after being plated on Matrigel following 1 h of VEGF treatment. The tube formation on Matrigel was enhanced in CXCR4+BMSCs under VEGF treatment. Total tube length in a field of view was 0.4 ± 0.05, 1.1 ± 0.16, and 1.3 ± 0.21 mm per mm2 the cultures with control, 10 and 20 ng/mL VEGF, respectively. Compared with controls, mean total tube length was increased sevenfold (p < 0.05, Fig. 4B). These findings suggest that VEGF induced CXCR4+BMSCs to perform functions of an endothelial phenotype.
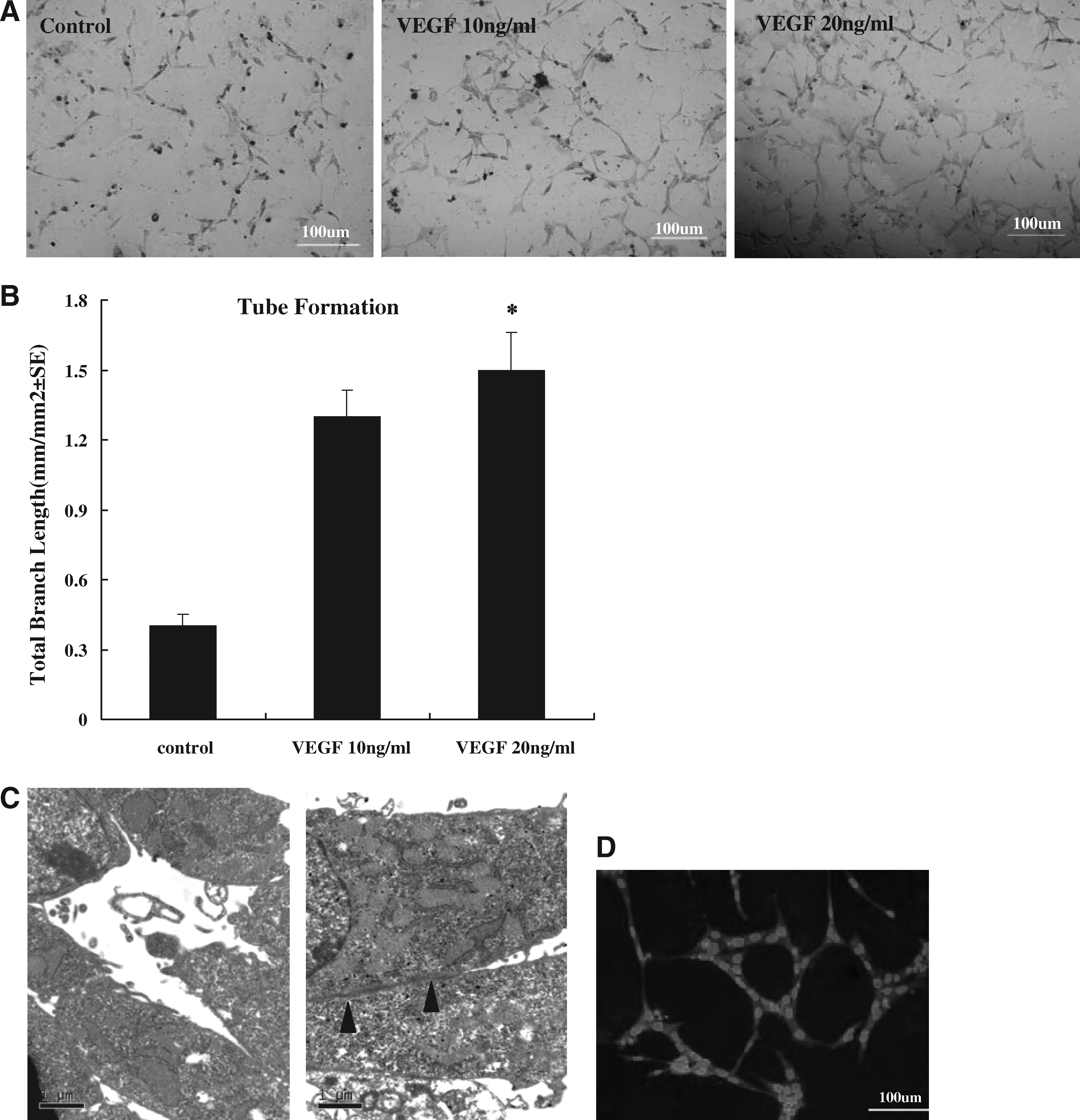
Effect of VEGF on tube formation of CXCR4+BMSCs in vitro. (
We confirmed an intact microvascular network in CXCR4+BMSCs by transmission electron microscopy analysis. The formation of capillaries tube was found and unaltered in size among CXCR4+BMSCs (Fig. 4C), one of characteristic morphology of ECs. In addition, the CXCR4+BMSCs also showed the cell–cell junction as ECs. Endothelial cell-to-cell junctions are complex structures formed by different adhesive molecules (Dejana et al., 1995; Lampugnani et al., 1995). ECs possess adherens junctions and tight junctions similar to those described in epithelial cells. Adherens junctions are ubiquitous along the vascular tree and are formed by transmembrane proteins belonging to the cadherin superfamily. Tight junctions comprise three types of transmembrane proteins, occludins, claudins, and junctional adhesion molecule. We examined whether CXCR4+BMSCs expressed VE-cadherin. As shown in Figure 4D, CXCR4+BMSCs were positive for VE-cadherin. The junction of CXCR4+BMSCs liked to the tight junctions and adherens junctions. The increase expression of PECAM-1 and vWF of CXCR4+BMSCs at both the mRNA and protein levels previously demonstrated these junctions. CXCR4+BMSCs differentiated into EC phenotypes including endothelial markers and angiogenic properties.
Local transfer of CXCR4+BMSCs increase the neo-capillary formation
To evaluate the in vivo effect of CXCR4+BMSCs on neo-capillary formation after arterial ischemic limb, mice were transplanted with CXCR4+BMSCs or total BMSCs to ischemic limbs 3 days after femoral artery ligation. Figure 5A shows representative photographs of ischemic and nonischemic skeletal muscle. Morphometric analysis revealed that neo-capillaries formation significantly increased in ischemic skeletal muscle tissues.
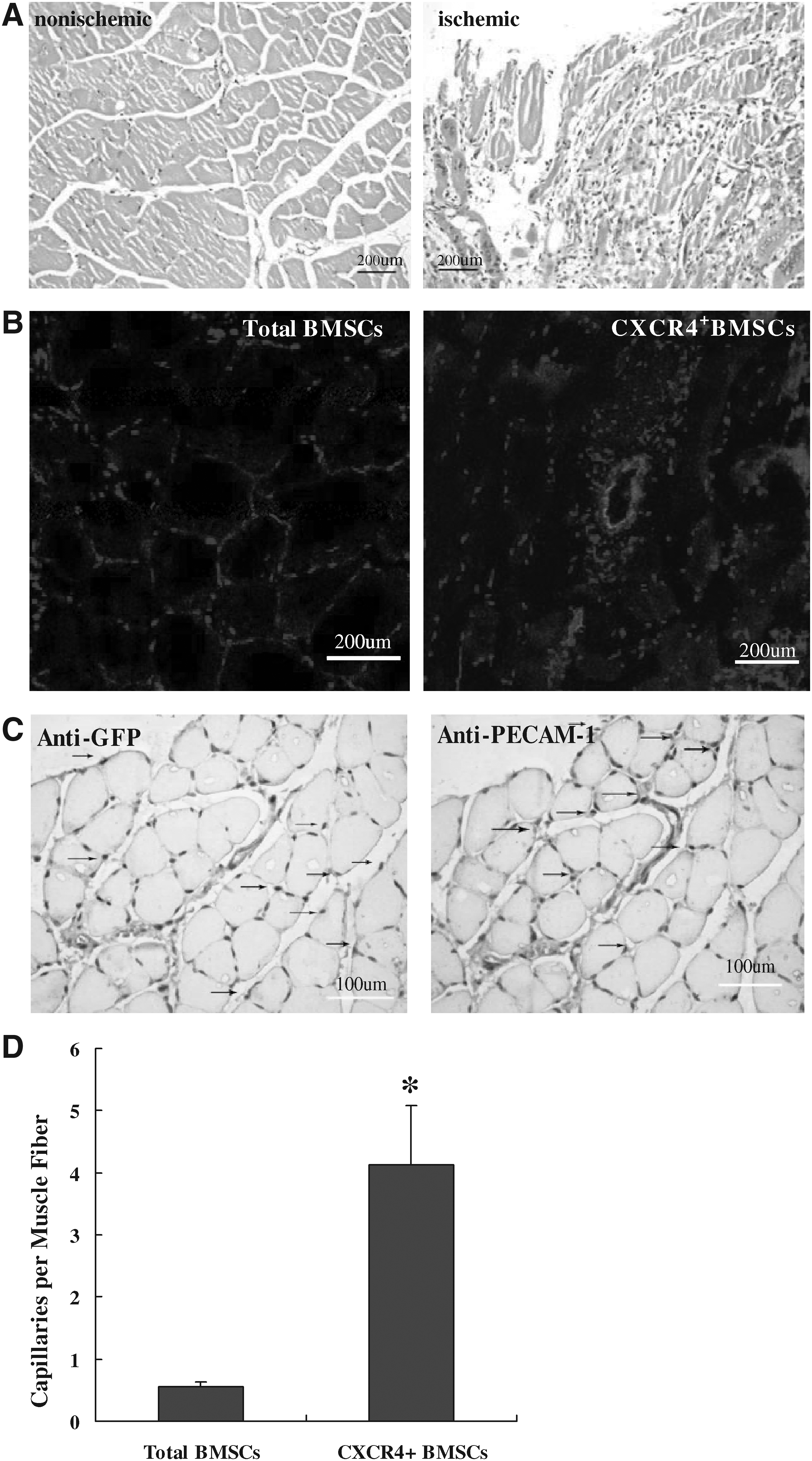
Transplanted CXCR4+BMSCs contribute to reendothelialization and repair. CXCR4+BMSCs and total BMSCs (5 × 105) transfected with plasmid encoding GFP were injected 3 days after under femoral arteries ligation (n = 6). After 28 days, skeletal muscle samples were harvested. (
We injected the CXCR4+BMSCs or total BMSCs transfected by GFP plasmid to ischemic limbs under femoral arterial ligation, after 4 weeks we directly detected the GFP-positive cells in skeletal muscle tissue on ischemic limbs under a fluorescent microphotograph (Fig. 5B). The luminal surface of the artery showed confluence in some ischemic muscle areas. In addition, immunochemical staining confirmed most double-positive cells in neo-capillaries for GFP and PECAM-1 on the consecution section (Fig. 5C). The other stained cells are positive for either anti-GFP or anti-PECAM-1 in neo. We analyzed the capillary density in the recovered tissues (Fig. 5D). The number of capillaries in each muscle fiber significantly increased in the CXCR4+BMSCs group (8.45 ± 1.20 pmf ) in comparison with control groups (1.43 ± 0.34 pmf, p < 0.05). These results revealed that local transfer of CXCR4+BMSCs significantly contribute to neovascularization formation and repair of ischemic tissues.
Discussion
In the present study, we demonstrated that VEGF induced expression of mature EC-specific markers PECAM-1 and vWF at both the mRNA and protein levels in CXCR4+ progenitor cells from BMSCs. In addition, we observed that CXCR4+BMSCs could uptake acLDL and tube-like structures on Matrigel and showed the ultrastructures of EC-like using an electron microscope. Then local applications of CXCR4+BMSCs in ischemic hindlimb model promoted neovascularization formation. The findings of this study indicate that CXCR4+BMSCs are one kind of EPCs, and play an important role in the neo-vascularization after arterial injury or ischemia. These findings strongly suggest that CXCR4+BMSCs have therapeutic implications for vascular diseases and are a promising cell source for cell-based vascular engineering and repair of injured vessels.
BMSCs have the potential to differentiate into a variety of specialized cells such as osteocytes, adipocytes, ECs, and smooth muscle cells. The BM-derived EPCs could promote early reendothelialization of the denuded vessels after injury and potentiate their vascular repair (Gulati et al., 2003). BMSCs are cell mixtures, and limit their use for the therapeutic potential of transplantation of cardiovascular diseases. We isolated CXCR4+ progenitor cells from BMSCs and found that CXCR4+BMSCs that were induced by VEGF increased PECAM-1 and vWF at both the mRNA and protein levels. These findings indicate that CXCR4+BMSCs had the phenotype of EPCs or ECs.
Increasing evidence indicates the importance of progenitor cells derived from BM in vascular development, homeostasis, and remodeling. Although the role of the various cytokines induced and their effects on the mobilization of circulating progenitor cells is unclear, the chemokine, stromal-derived factor-1 (SDF-1), binds to its specific receptor, CXCR4, and plays a role in the mobilization and differentiation of stem/progenitor cells (Oswald et al., 2004). SDF-1 has shown to be important for trafficing of BM-HSCs and the homing to the ischemic tissue. Several type cells such as cardiomyocytes, muscle-derived fibroblasts, and endothelial cells secrete SDF-1 (Ceradini et al., 2004; Ratajczak et al., 2003). Recent studies indicated that SDF-1 was produced in response to tissue damage under hypoxic and ischemia, and that played an important role in the mobilization of CXCR4+EPCs (Abbott et al., 2004). Given that SDF-1/CXCR4 interaction is a critical factor in mobilization and homing of progenitor cell, CXCR4+ progenitor cells may be involved in ischemia tissue repair in response to enhanced SDF-1 expression.
Some groups reported that Flk-1+ cells derived from ES cells could differentiate into cardiovascular cells, and suggested Flk-1+ was a marker for vascular progenitor cells (Hirashima et al., 2003; Sone et al., 2003; Yamashita et al., 2000). Recently, studies reported that the circulating CXCR4+ progenitor cells increased after cardiac surgery, mobilized, and recruited to regenerating tissues and neovascularization (Ceradini et al., 2004; Jin et al., 2006; Petit et al., 2007). The reports indicated that CXCR4 was the special marker of these stem/progenitor cells. In the present study, we demonstrated that CXCR4+BMSCs could differentiate into EC-like cells, and that CXCR4 might be used as a sorting marker for the isolation of EPCs or BMSCs. These findings suggest that this novel marker is beneficial for obtaining EPCs with high purity.
A panel of genetic, antigenic, and functional assays is required to provide optimal characterization of mature endothelial populations differentiated from BMSCs. We demonstrated that CXCR4+BMSCs had EC characteristics phenotypes and angiogenic properties based on the following observations. First, CXCR4+BMSCs displayed the morphology of EC in culture and increased the mRNA and protein expressions of PECAM-1 and vWF under VEGF treatment. Second, CXCR4+BMSCs rapidly uptake Dil-acLDL and had the potential ability of tube formation. Third, the ultrastructure of CXCR4+BMSCs displayed the junctions of EC-like and Weibel-Palade-like bodies in vitro. Finally, CXCR4+BMSCs were observed in the injury vascular artery and neo-capillaries formation after arterial ischemic limb experiments. It is thus likely that CXCR4+BMSCs shared a similar phenotype and function as EPCs.
The ultimate goal of understanding stem cell biology is the potential for using these cells therapeutically in clinical medicine. Accumulating evidence indicates that adult or vascular progenitor cells (Kong et al., 2004; Xu et al., 2004), such as BMCs (Fujiyama et al., 2003; Tanaka et al., 2003; Xu et al., 2004) and EPCs (Kong et al., 2004; Strehlow et al., 2003; Werner et al., 2003) or EPCs influence neoangiogenesis, have beneficial effects on neovascularization following ischemia and reendothelialization after vascular injury, thereby providing a potentially novel therapeutic option for prevention of vascular diseases such as atherosclerosis, restenosis, and graft failure. Shiba et al. (2009) reported the CXCR4 expression of BMSCs enhances therapeutic angiogenesis in ischemic cardiovascular diseases. Seeger et al. (2009) reported CXCR4+BMSCs exhibit an increased therapeutic neovascularization potential in acute ischemia. In the present study, CXCR4+BMSCs recruited to the site of the injured artery where they increased the neo-capillaries formation and resulted in neo-angiogenesis vessels in ischemic muscle, indicating that CXCR4+BMSCs contributed to reendothelialization and tissue repair and increased the efficiency of stem cell therapy.
In conclusion, we demonstrated that CXCR4+BMSCs could differentiate into ECs in vitro and recruited to the ischemic limb where they resulted in the early development of neointimal formation in vivo. Our findings raise the possibility that CXCR4 was selected as the surface marker of EPCs. Thus, we have established a novel method for producing EPCs from BMSCs, which regenerated the neovascularization in ischemic tissue and repair vascular lesion after arterial injury. These findings provide important details on targeting therapy that promote BMSCs differentiation and activation, and provide direct evidence to support adult stem cell-based therapeutic approaches.
Footnotes
Acknowledgments
This work was supported by grants from the Natural Science Foundation of China (No. 30570725).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
The first two authors contributed equally to this work.