
Abstract
Abstract
Unlike embryos derived from fertilization, most cloned embryos die during postimplantation development, and those that survive to term are frequently defective. Many of the observed defects involve placenta. Abnormal placentation has been described in several cloned species. Imprinted genes are important regulators of placenta growth, and may be subjected to faulty reprogramming during somatic cell nuclear transfer. We aimed to determine the expression levels and methylation patterns of imprinted genes in placentas of live cloned piglets and dead ones. Quantitative real-time reverse transcriptase-polymerase chain reaction (RT-PCR) analysis showed that the expression of all four imprinted genes (IGF2, H19, PEG3, and GRB10) was significantly reduced in placentas of dead clones compared with placentas of live cloned piglets and controls (p < 0.05). In contrast, both live and dead cloned piglets exhibited steady-state mRNA levels for these genes within the control range (p > 0.05). Transcript levels for these genes in live clones rarely differed from those of controls in both piglets and placentas. Examination of the methylation status of DMR2 of IGF2 and CTCF3 of H19 genes revealed that both genes exhibited significant high methylation levels in placentas of dead clones compared with placentas of live clones and controls. In contrast, both genes showed a normal differential methylation pattern in live cloned piglets and their placentas compared with controls. Importantly, dead cloned piglets also showed a normal pattern. Our results suggest that abnormal expression of imprinted genes in placenta may contribute to the development failure in pig somatic cell nuclear transfer (SCNT), which may be caused by abnormal methylation patterns in differentially methylated regions (DMRs) of imprinted genes as a result of incomplete reprogramming during SCNT.
Introduction
To date, abnormal placentation has been described in several cloned species (Cibelli et al., 2002). Placentas of cloned mice are enlarged without exception (Suemizu et al., 2003), and those of cloned calves have fewer but much larger placentomes, presumably to compensate for the reduced number of sites of maternal–fetal exchange (Chavatte-Palmer et al., 2002). It is reported that from days 30–90, most bovine somatic cell nuclear transfer (SCNT) fetuses died because of abnormal placentas (Hill et al., 2000). Recent studies also found that extraembryonic tissues from cloned porcine embryos or SCNT-derived placentas were frequently abnormal (Chae et al., 2008, 2009; Lee et al., 2007). Abnormal placental development and function probably account for a high proportion of post implantation fetal losses in clones (Wells et al., 1999). It is possible that many of the abnormalities observed in cloned fetuses may be secondary to defects in placental function (Yang et al., 2007). Several genes have been found to be normally expressed in cloned fetuses but abnormally expressed in the placentas, which supports the fact that poor cloning efficiency is largely due to placenta abnormality (Inoue et al., 2002; Yang et al., 2005).
Genomic imprinting, the preferential expression of one parental allele, is regulated primarily by DNA methylation of CpG islands (Li et al., 1993). Naturally, differential epigenetic marks on the two parental alleles are established during gametogenesis (Tremblay et al., 1995); however, nuclear transfer bypasses this step and may establish (or maintain) the wrong imprinting. This may cause the dysregulation of gene expression (Khosla et al., 2001; Mann et al., 2003; Ogawa et al., 2003; Yang et al., 2005). The use of knock-out models of imprinted genes with an altered balance between placental and fetal growth highlights the signaling that occurs between fetus and placenta, to match the fetal demand with the placental supply of nutrients in a tightly controlled process for most of gestation (Angiolini et al., 2006; Reik et al., 2003; Rossant and Cross, 2001). As imprinted genes play an important role in maternal–fetal exchange during gestation, the altered expression of these genes in neurotrophin (NT) placenta may be related to the high mortality rate of cloned embryos during in utero development. However, there are fewer studies focusing on imprinted genes expression in cloned animal's placenta. A study of cloned mice found that several imprinted genes (as well as several nonimprinted genes) showed abnormal low expression in cloned mouse placentas, whereas no differences are seen in fetal expression (Inoue et al., 2002). Abnormal allelic expression pattern of the imprinted Igf2r gene was also found in placentas but not in organs of cloned bovine calves (Yang et al., 2005). These facts indicate that functions of imprinted genes in placenta are highly correlated with developmental potential of cloned embryos. However, there is no report focusing on issues of this type in pig SCNT.
Methylation analysis of imprinted genes has been used to study nuclear reprogramming during cloning (Liu et al., 2008; Mann et al., 2003; Young et al., 2001). Many imprinted genes contain one or more differentially methylated regions (DMRs) exhibiting allelic methylation differences (Ferguson-Smith and Surani, 2001; Miyoshi et al., 2006). Of those identified, IGF2 and H19 have been most widely studied. Furthermore, the methylation patterns of these genes have been studied in gametes and porcine preimplantation embryos (Han et al., 2008; Park et al., 2009). So we analyzed the methylation status of IGF2 and H19 to clarify whether the aberrant expression is caused by abnormal methylation patterns.
To date, direct evidence that aberrant expression and methylation of imprinted genes in placenta of cloned piglets causes abnormal fetal development is lacking. We propose that many of the imprinted genes express abnormally in cloned pig placenta rather than fetus or piglet; this may contribute to problems with placental development and finally cause development failure of cloned fetus. We suspect that the aberrant expression may be caused by abnormal methylation patterns in DMRs of imprinted genes as a result of incomplete reprogramming during SCNT. To address these questions, we compared expression levels of four imprinted genes (IGF2, H19, PEG3, and GRB10) among dead cloned piglets, live cloned piglets, and natural breeding piglets as well as their corresponding placentas by quantitative real-time reverse transcriptase-polymerase chain reaction (RT-PCR). We also compared methylation status of IGF2 and H19 by bisulfite sequencing.
Materials and Methods
Production of cloned piglets and tissue collection
In vitro maturation (IVM), SCNT, and embryo culture were performed according to established methods (Liu et al., 2007). Briefly, adult fibroblast cells were obtained from a newborn piglet (Northeast Min Pig—a local breed). Tissues from the ear were rinsed in Dulbecco's phosphate-buffered saline (PBS) (Sigma, St. Louis, MO, USA). The tissues were minced into small pieces (1–3 mm). The minced fetal tissues were incubated in DMEM (Gibco, Grand Island, NY, USA) supplemented with 0.25% trypsin-EDTA (Gibco) solution for 10 min at 37°C, followed by pipetting for dissociation. Dissociated cells were cultured in DMEM supplemented with 10% fetal bovine serum (FBS) (Gibco) at 38.5°C in a humidified atmosphere of 5% CO2 until confluent. The cells were cryopreserved in a medium of 90% v/v FBS and 10% v/v DMSO (Sigma), and then stored in liquid nitrogen until use.
Porcine ovaries were collected from a local slaughterhouse and kept in saline at 32–37°C. Antral follicles (3–5 mm in diameter) were aspirated with an 18-gauge needle. Aspirated oocytes with an evenly granulated cytoplasm and three uniform layers of compact cumulus cells were selected and cultured in four-well plates (Nunc, Naperville, IL, USA) containing 500 μL of TCM199 (Gibco) based medium for 42 h. At the end of IVM, the cumulus cells of cumulus–oocyte complexes (COCs) were removed by vortexing for 2 min in TCM199 based medium supplemented with 0.5% hyaluronidase (Sigma). Matured eggs with the first polar body were moved to TCM199-based medium supplemented with 7.5 μg/mL cytochalasin B (Sigma). The first polar body and protrusion membrane were then removed with a beveled pipette. A single donor cell was injected into the perivitelline space and electrically fused using two direct pulses of 120 V/mm for 30 μsec in fusion medium. The fusion medium consisted of 0.28 M mannitol, 0.1 mM MgSO4, and 1 mM CaCl2 (Sigma). Fused eggs were cultured in PZM-3 medium for 1 to 2 days in an atmosphere of 5% CO2 and 95% air at 38.5°C prior to embryo transfer.
Potential surrogate gilts at more than 8 months old were checked for estrus twice a day. The SCNT embryos were transferred to the oviducts of the natural cycling gilts on the first day of standing estrus. The SCNT embryos (approximate 150 embryos/surrogate) were transferred into the oviducts of each synchronized recipient. Eighteen recipients were used as surrogate mother (matured LWD; Landrace × Large White × Duroc crosses). Three recipients became pregnant, and 14 piglets were born. Nine of them were born alive (three died within 1 day after birth); two of them died at birth; three of them died in uterus (like mummy).
Piglets in our study consisted of three groups. Group 1 included three live cloned piglets that were healthy newborn and lived healthy for more than 2 years up to now. Group 2 included five dead cloned piglets that died at or shortly (within 1 day) after birth. Group 3 included four natural breeding piglets with normal phenotype and without history of health disorders, which were used for control. One sample from the ear and one sample from the placenta were collected for each piglet. All samples from piglets and placentas were collected within 1 day after birth; the dead cloned piglets refer to piglets died at or shortly (within 1 day) after birth. The breed of all piglets analyzed was the Northeast Min Pig (a local breed). All samples were immediately frozen in liquid nitrogen after collection.
Quantitative real-time PCR
Total RNA was extracted using the PureLinkTM Micro-to-Midi system (Invitrogen, Carslbad, CA, USA) according to the manufacturer's instructions. RNA quality was confirmed by ratios of A260/A280 (all between 1.8 and 2.0) and also by gel electrophoresis for the presence of two clear ribosomal RNA bands. Only RNA samples that did not show signs of degradation were used in this study. DNase I (Promega, Madison, WI, USA) was then added to digest the genomic DNA in the samples by incubating at 37°C for 20 min. Finally, DNase was inactivated at 75°C for 5 min. At this point, the total RNA was used for reverse transcription (RT). The RT reaction was performed using PrimeScriptTM RT Reagent Kit (TaKaRa, Japan) under the following conditions. The 20-μL reaction volume contained 1 μL 100 μM random hexamer primer, 4 μL 5 × RT buffer, 1 μL RT enzyme Mix, 1 μL Total RNA (<500 ng), and 13 μL RNase Free dH2O. The reaction mixture was incubated at 37°C for 15 min, followed by 85°C for 5 sec to inactivate the RT enzyme. The cDNA was stored at −20°C until further analysis.
Quantification of all transcripts was performed by quantitative real-time PCR. PCR was performed using SYBR Premix Ex TaqTM (TaKaRa) and the 7300 Real-Time PCR System (Applied Biosystems, Bedford, MA, USA). Sequences and GeneBank accession number of the primer used for amplification of the target genes are presented in Table 1 (primers were derived from Jiang et al., 2007). Reactions were performed in 96-well optical reaction plates (Applied Biosystems). Each reaction mixture (50 μL) contained 1 μL (<25 ng) cDNA solution, 1 μL 10 μM of each specific primer, 1 μL 50 × SYBR Green Dye, and 25 μL 2 × Ex Taq. Thermal cycling conditions were 95°C for 10 sec, followed by 40 PCR cycles of 95°C for 5 sec and 60°C for 31 sec, and finally a melting cycle consisting of 95°C for 15 sec, 60°C for 1 min, and 95°C for 15 sec. The quantification of gene transcripts of four imprinted genes (IGF2, H19, PEG3, and GRB10) was carried out in three replicates. The 18S ribosomal RNA was used as an endogenous control throughout this study. The specificity of the PCR reaction was confirmed by a single peak in the dissociation curve and also by a single band in agarose gel electrophoreses (see Supplementary Information Fig. S1). As negative controls, tubes were always prepared in which cDNA was omitted during the real-time reaction.
The comparative CT method was used for relative quantification of target gene expression levels. Each pair of primers was confirmed for equal amplification efficiency to primers of the endogenous control (18S ribosomal RNA). CT value was calculated by the Sequence Detection System software (Applied Biosystems). The ΔCT value was defined as CT (target gene) − CT (18S rRNA). The ΔΔCT value was defined as ΔCT (sample) − ΔCT (calibrator). The relative expression level of a target gene is calculated as: 2−ΔΔCT.
Bisulfite sequencing analysis
Genomic DNA was extracted from tissue samples using the Universal Genomic DNA Extraction Kit (Takara) and followed by treatment with sodium bisulfite to convert all unmethylated cytosine to uracil using EZ DNA Methylation-GoldTM Kit (Zymo Research, Orange, CA, USA) according to the manufacturer's protocol. Briefly, purified genomic DNA (200–500 ng) was denatured at 98°C for 10 min. After denaturation, the sample was incubated in 130 μL CT (cytosine to thymine) conversion reagent in the dark at 64°C for 2.5 h. Modified DNA was then desalted, purified, and diluted with 20 μL of distilled water. Subsequently, bisulfite PCR (BS-PCR) amplification was carried out using 1–2 μL (10–50 ng) of modified DNA per PCR run.
Because limited sequence information of other imprinted genes is available in porcine, we only examined methylation status of IGF2 and H19. DMRs of these two genes were based on previous studies (Han et al., 2008). A region of 268 bp between exon8 and exon9 of the IGF2 gene and a region of 205 bp upstream of the promoter of the H19 gene were selected, respectively, as their differentially methylated regions. Nested PCR amplifications of bisulfite-treated DNA were performed using primers as described in Table 2 (primers were derived from Han et al., 2008). PCR amplification was performed using Hot Start TaqTM Polymerase (TaKaRa) in 20 μL reaction volume with a thermo profile of 98°C for 5 min, 94°C for 30 sec, 72°C for 30 sec, for 40 cycles, followed by 72°C for 10 min. The annealing temperature used for each primer pair is shown in Table 2. Two microliters of products from the first amplification reaction were used in the second PCR reaction. The amplified products were verified by electrophoresis on a 1% agarose gel and gel-purified using the Agarose Gel DNA Purification Kit (Takara). Purified fragments were cloned into the pMD18-T Vectors (TaKaRa). The positive clones confirmed by PCR were sequenced using an automatic sequencer (ABI PRISM-77). Only clones with ≥95% cytosine conversion were selected (see Supplementary Fig. S2). At least 10 replicates were performed for each DMR locus in each sample, and at least two separate bisulfite treatments were carried out for each locus to confirm the results.
Statistical analysis
The levels of gene expression among three groups were analyzed using analysis of variance (ANOVA) and t-test by StatView (SAS Institute Inc., Cary, NC, USA); p values < 0.05 were considered significantly different.
Results
Expression levels of imprinted genes in cloned piglets and their placentas
Expression levels of four imprinted genes were compared among live cloned piglets, dead cloned piglets, control piglets, and their corresponding placentas (Fig.1). For the ease of comparison, the average of expression levels of each gene from control group was set as 1.
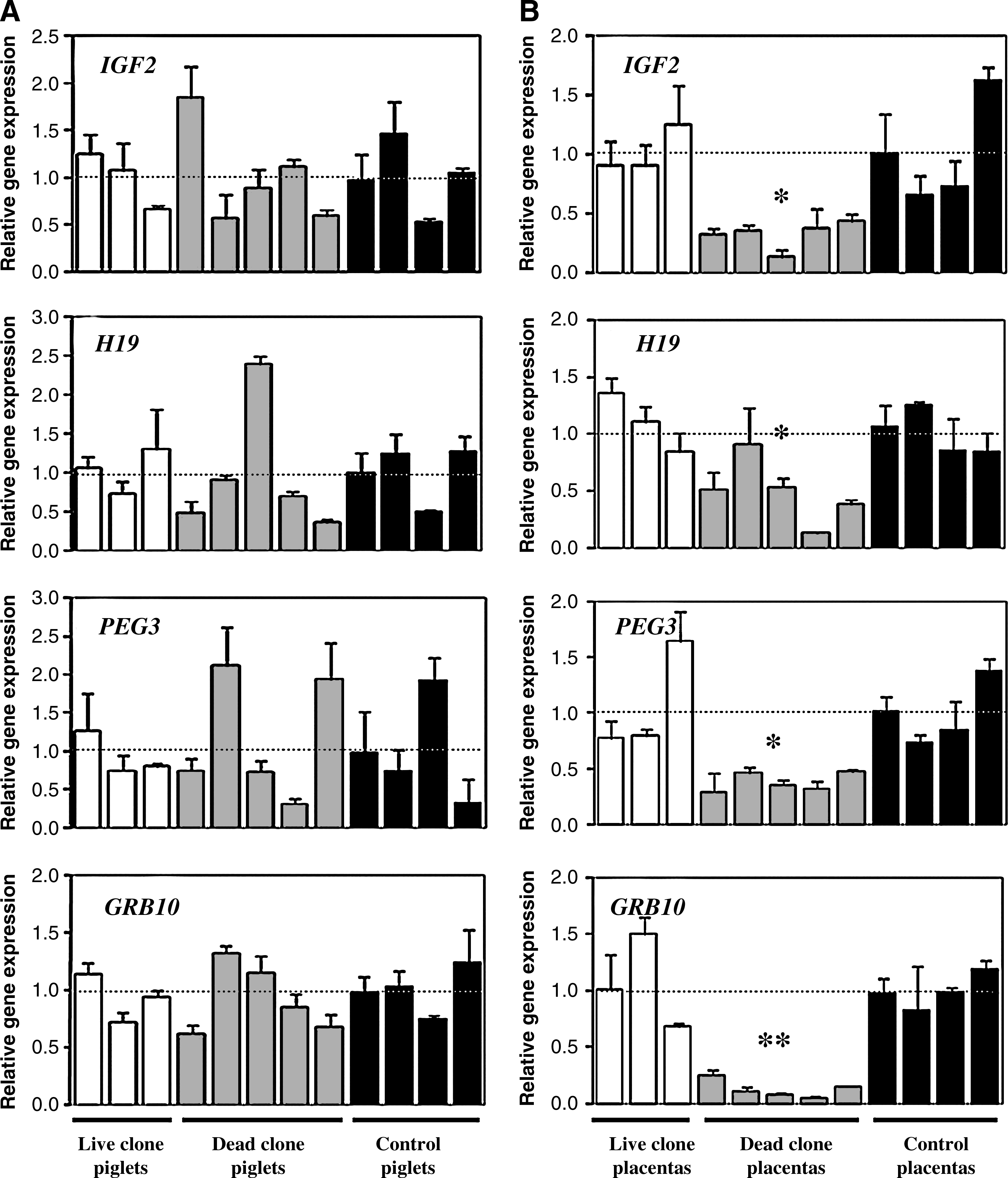
Transcript levels of four imprinted genes (IGF2, H19, PEG3, and GRG10) in cloned piglets (
Quantitative real-time PCR analysis showed that the expression of all four imprinted genes (IGF2, H19, PEG3, and GRB10) was significantly reduced in placentas of dead clones compared with placentas of live clones and placentas of controls (p < 0.05). Contrastingly, the dead cloned piglets exhibited steady-state mRNA levels for these genes within the control range (p > 0.05). Transcript levels for these genes rarely differed from those of controls in both live cloned piglets and their corresponding placentas.
Bisulfite analysis of IGF2 and H19 DMRs in cloned piglets and their placentas
The methylation pattern at DMR2 of IGF2 and CTCF3 of H19 in live cloned piglets, dead cloned piglets, control piglets, and their corresponding placentas are shown in Figures 2–5.

Methylation status of DMR2 of IGF2 gene in cloned and natural breeding piglets. Lollipops represent all examined CG dinucleotides. Black and white circles represent methylated and unmethylated CpGs, respectively. Each line represents a separate clone.

Methylation status of DMR2 of IGF2 gene in placentas of cloned and natural breeding piglets. Lollipops represent all examined CG dinucleotides. Black and white circles represent methylated and unmethylated CpGs, respectively. Each line represents a separate clone.

Methylation status of CTCF3 of H19 gene in cloned and natural breeding piglets. Lollipops represent all examined CG dinucleotides. Black and white circles represent methylated and unmethylated CpGs, respectively. Each line represents a separate clone.

Methylation status of CTCF3 of H19 gene in placentas of cloned and natural breeding piglets. Lollipops represent all examined CG dinucleotides. Black and white circles represent methylated and unmethylated CpGs, respectively. Each line represents a separate clone.
Both genes, IGF2 and H19, exhibited significant high methylation levels in placentas of dead clones (IGF2 87.3%; H19 75.8%) than those in placentas of live clones (IGF2 44.1%; H19 20.2%) and placentas of controls (IGF2 40.2%; H19 14.6%). In contrast, both genes showed a normal differential methylation status in live cloned piglets (IGF2 72.9%; H19 50.6%) and their placentas compared with controls (IGF2 75.7%, H19 51.8%). Importantly, dead cloned piglets also showed a normal methylation status of these two genes (IGF2 78.3%; H19 50.2%). Therefore, an aberrant methylation pattern appears to occur frequently in imprinted genes of dead cloned piglets associated placentas rather than the piglets themselves.
Discussion
It is generally accepted that failures in development of the placenta are the main causes of fetal and prenatal death of nuclear transferred embryos (Wilmut et al., 2002). Recent research also provided evidence that extraembryonic tissues or placentas derived by SCNT were frequently abnormal (Chae et al., 2008, 2009; Lee et al., 2007). However, direct evidence that aberrant expression and methylation of imprinted genes in placenta of cloned piglets causes abnormal fetal development is lacking. Here, we reported for the first time that expression and methylation of imprinted genes in placenta of cloned pig was associated to fetal developmental potential in pig SCNT.
Expression levels of four imprinted genes among live cloned piglets, dead cloned piglets, natural breeding piglets, and their corresponding placentas were examined. We found that expression levels of all four genes (IGF2, H19, PEG3, and GRB10) in dead cloned piglets associated placentas were significantly reduced; however, expression levels of these genes in dead cloned piglets were normal. This was consistent with findings in cloned mice and cloned cattle (Inoue et al., 2002; Yang et al., 2005). A study of cloned mice found that several imprinted genes (as well as several nonimprinted genes) showed abnormally low expression in cloned mouse placentas, whereas no differences were seen in fetal expression (Inoue et al., 2002). Abnormal allelic expression pattern of the imprinted Igf2r gene expression was also found in placentas but not in organs of cloned bovine calves (Yang et al., 2005). A recent report provided a comprehensive study of imprinted genes in swine, which found that abnormalities existed largely in parthenogenetic placenta, such as decreased chorioallantoic folding, decreased chorionic ruggae, and reduction of fetal–maternal interface surface, at day 30 of gestation (Bischoff et al., 2009). These results suggest that the placenta is especially vulnerable to problems of imprinted genes in cloned animals. As the trophectoderm eventually gave rise to the placenta, abnormal placenta development may be affected by trophectoderm defects (Koo et al., 2002, 2004). Tetraploid complementation suggests that abnormal cells derived from nuclear transfer are preferentially incorporated into the trophectoderm rather than the inner cell mass (ICM) (Eakin and Behringer, 2003; Nagy et al., 1990; Wakayama et al., 2006). As reprogramming of imprinted genes is especially error-prone, and the majority of imprinted genes involves in placental function (Coan et al., 2005; Rossant and Cross, 2001), the placenta would be preferentially vulnerable to problems of imprinted genes brought about by SCNT.
In an effort to determine whether aberrant expression of imprinted genes is caused by abnormal methylation patterns of their DMRs, we analyzed the methylation status of DMR2 of IGF2 and CTCF3 of H19 genes. Because there are not enough methylation-sensitive restriction enzyme sites, we could not investigate DMRs with the methylation-sensitive restriction enzyme technique. Instead, we chose the BS-PCR technology, which can be used to assess the methylation status of all cytosines in a desired sequence (Warnecke et al., 1998). We found that both genes exhibited hypermethylation in placentas of dead clones compared with placentas of controls. In contrast, both genes showed a normal differential methylation pattern in dead cloned piglets (compared with controls). There was no difference in methylation pattern in both live cloned piglets and their corresponding placentas. In natural breeding animals, expression of IGF2 and H19 is reciprocal, when one of these genes is transcribed, another gene is silenced. Although the reduced expression level of H19 is related to the hypermethylation status of CTCF3, unexpected, we did not find repressed expression of H19 is associated with an increased expression of IGF2 in placentas of dead cloned piglets. In cloned mouse, Ogawa et al. (2003) also found that the reduced expression of H19 was not associated with increased expression of Igf2 in case of hypermethylation of the H19 DMR. We suspect that the shared enhancer elements that these two genes compete for might be disrupted (Caspary et al. 1998). The mechanism needs further study.
In our study, we found that aberrant expression and methylation exist in placentas rather than in piglets. As we need the cloned piglets to investigate their subsequent development, we did not collect other tissues for analysis, we choose ear tissue as representative of the piglet to compare with the placenta. Analysis of expression and methylation of these genes in multiple tissues is more convincing, because expression and methylation of these genes may be tissue-specific. For example, expression of imprinted genes has been found to vary with tissue type in cloned piglet (Jiang et al., 2007). Similarly, methylation of DMR2 of the Igf2 gene is tissue-specific in the mouse (Weber et al., 2001). So comparison of the expression and methylation of imprinted genes in other tissues is our further consideration.
All cloned pigs in this study were derived from the same donor cell line and were therefore genetically identical, and several cloned pigs shared the same foster mother, but they had large variations in gene expression and epigenetic modification in placentas. This is consistent with other reports from cloned cattle and mice (Kohda et al., 2005; Li et al., 2005). This suggests that epigenetic reprogramming is an independent event in each cloned embryo.
In conclusion, our results indicate that aberrant methylation of imprinted genes exist in placenta rather than in fetus or piglets, which may lead to their abnormal expression and finally affect the normal developmental regulation of fetal and placenta during porcine SCNT. It may be valuable to determine the superior epigenetic modifications of imprinted genes for cloned embryo's trophoderm cells and to establish a procedure for selecting and sorting them to produce cloned animals efficiently. On the other hand, although dead cloned piglets associated placentas exhibited aberrant imprinted gene expression, expression of dead cloned piglets was indistinguishable from controls. This indicates that epigenetic errors accumulated during SCNT may be preferentially incorporated into trophoblast lineage rather than the ICM lineage. In this respect, and because applications such as therapeutic cloning are not considered to use trophoblast cells (Hochedlinger and Jaenisch, 2003), ntESCs derived from ICM cells perhaps have great potential for therapeutic applications.
Footnotes
Acknowledgments
The authors are thankful to Dr. Yongjun Shu for his generous consultation on statistical analysis. The authors are also thankful to colleagues in the “Lab of Embryo Biotechnology” for their helpful discussions. This work was supported by National Natural Science Foundation of China, NSFC (30871431), the National High Technology Research and Development Program (863 Program) of China (2008AA101006), and the New Century Talent Foundation of Heilongjiang Province (1153-NCET-007).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.