
Abstract
Abstract
Recently, several research groups have shown that germ cells can be produced in vitro from pluripotent embryonic stem cells (ESCs). In the mouse, live births of offspring using germ cells induced from ESCs in vitro have been reported. Furthermore, some efficient methods for inducing the useful number of germ cells from ESCs have also been developed. On the other hand, in primates, despite the appearances of germ cell-like cells including meiotic cells were observed by spontaneous differentiation or introducing transgenes, it has not been determined whether fully functional germ cells can be derived from ESCs. To elucidate the property for the germ cells induced from primate ESCs, specification of the promoting factors for the germ cell development and improving the efficiency of germ cell derivation are essential. Leukemia inhibitory factor (LIF) has been reported as one of the important factors for mouse primordial germ cell (PGC) survival in vitro. However, the effects of LIF on germ cell formation from pluripotent cells of primates have not been examined. The aim of this study is to determine whether LIF addition can improve in vitro germ cell production from cynomolgus monkey ESCs (cyESCs). After 8 days of differentiation, LIF added culture induced dome-shaped germ cell colonies as indicated by the intense expression of alkaline phosphatase activity (ALP). These cells also demonstrate high-level expression of the germ cell-marker VASA, OCT-4, and BLIMP-1, and show SSEA-1 expression that supports their early stage germ cell identity. Finally, we observed that adding LIF to differentiating cultures inhibited meiotic gene expressions and increased the percentage of ALP-positive cells, and demonstrate that the addition of LIF to differentiation media increases differentiation of early germ cells from the cyESCs.
Introduction
In rodents, production of the germ cells that possess the property for fertilization and development from embryonic stem cells (ESCs) has already been reported (Nayernia et al., 2006), and the method of deriving germ cells from ESCs took notice as useful models to observe germ cell development of mammals. Recently, we and some researchers reported that the ESCs of primates differentiate to germ cells spontaneously (Clark et al., 2004; Teramura et al., 2007). Furthermore, in a more recent report, the strategy for increasing the number of the germ cells that can enter the meiosis by overexpressing some germ cell-related genes has been shown (Kee et al., 2009). However, functional matured germ cells, which can fertilize and progress fetal development, have not been determined. To develop the way for inducing functional germ cells from the primate ESCs, determination of the factors to promote germ cell development in vitro and improving the efficiency of germ cell derivation is essential.
Leukemia inhibitory factor (LIF) is a crucial factor for maintaining pluripotency and self-renewal in mouse ESCs (Niwa, 2001), and ordinarily used in pluripotent cell culture in rodents. Furthermore, it has been determined that LIF is important for maintenance of primordial germ cells (PGCs) in the mouse (De Felici et al., 1992; Koshimizu et al., 1996). Although the LIF-STAT3 signal does not relate to the signal pathway for maintain pluripotency in primates and LIF is not essential for self-renewal of the primate ESCs (Sumi et al., 2004), it was reported that LIF elevate the survival rate of human PGCs in vitro (De Felici et al., 2004). However, the detailed functions of LIF have not been well examined in the germ cells of primates, and the effectiveness for germ cell formation from pluripotent cells has not been examined. In the present study, we used human recombinant LIF to improve the efficiency for germ cell differentiation from cynomolgus monkey ESCs (cyESCs), and examined their function by observing the expression of germ cell specific marker genes and meiosis-specific marker genes.
Materials and Methods
Culture of cyESCs
For the present study, we used a previously established cynomolgus monkey ES cell line Cyk-3 (Takehara et al., 2008). The cyESCs were maintained on culture dishes containing a layer of mitotically inactivated mouse embryonic feeder (MEF) cells in ES medium composed of Knockout-DMEM (Invitrogen Corporation, Carlsbad, CA, USA), 1 mM
Germ cell differentiation from the cyESCs
The cyESCs were dissociated into single cells by treating with TrypLE Express cell dissociation enzyme (Invitrogen) for 3 min, followed by the addition of fresh media to inhibit further digestion. Then, the cells were collected, resuspended in ES medium,and filtered through 40-μm meshes (BD Biosciences Discovery Labware, Bedford, MA, USA) to obtain a single-cell suspension. Dissociated cyESCs were seeded at a concentration of 3 × 104 cells/cm2 onto gelatin-coated dishes without feeder layers in the presence of Y-27632. Then the cells were cultured for 8 days in 20% KSR-DMEM with or without 1000 units/mL of recombinant Human Leukemia Inhibitory Factor (hLIF; Millipore, MA, USA). For the experiment of JAK–STAT signal inhibition, JAK inhibitor I (Calbiochem, La Jolla, CA, USA) 6 μg/mL were added to the culture medium.
ALP staining
For ALP staining, we used an ALP kit (Sigma-Aldrich) according to the instructions provided.
RT-PCR and real-time PCR
Total RNA was extracted using TRIzol (Invitrogen) according to the manufacturer's instructions. Single-strand cDNA was prepared from total RNA using a random primer under standard conditions with a High Capacity cDNA reverse transcription kit (Applied Biosystems, Foster City, CA, USA). The cDNA from each sample was diluted and used for a reverse transcription polymerase chain reaction (RT-PCR)-based assay for VASA, OCT-4, BLIMP-1, DMC-1, SCP-1, and β-Actin. RT-PCR with total cDNA was performed using platinum Taq PCRx DNA polymerase (Invitrogen) and following primer sets (listed 5′ to 3′ in the order of forward and reverse); DMC-1, GATTCAATAATGGCAGTTTTTCGA, CATCTGGGCCAATTTTTGCT; SCP-1, ACAGCGAAAAGCCATTCAGG, TCTGGAATTCTCAGCTTGCA. PCR conditions were as follows: denaturation at 94°C for 30 sec, followed by annealing at 60°C for 30 sec and extension at 72°C for 30 sec. The cDNA prepared from testis and fibroblast cells of cynomolgus monkey were used as positive control and negative control for the RT-PCR assays. PCR products were subjected to electrophoresis in 2% agarose gel and visualized by ethidium bromide staining.
Quantitative real-time PCR with total cDNA was performed using Perfect Real-time SYBR II Premix Ex-Taq™ (Takara Bio, Inc., Shiga, Japan) and following primer sets (listed 5′ to 3′ in the order of forward and reverse): VASA, AAGTTAATTTCTTGCCCAGGAATG, TGCAGCCAACCTTTGAATTC; OCT-4, ATTTGGGTAGGTGTTCAGCC, CGCCGCTTACTCATGTTCTT; BLIMP-1, TTTGCAGAAAGGCTTCACTAC, AGTGCTCGGCTGCTTGAG; β-Actin, AAATCGTGCGTGACATTAAGGA, GCCGCTGTGGCCATCTC. PCR amplifications were performed with the 7700 real-time PCR System (Applied Biosystems) at 95°C for 20 sec followed by 40 cycles of 95°C, 5 sec; 60°C, 30 sec. Reactions were replicated three times for two different samples that were independently differentiated and prepared. All experiments included negative controls consisting of no cDNA for each primer pair. Fold changes were calculated relative to control treatment using the 2−ΔΔCt method (Dussault and Pouliot., 2006). β-Actin expression was used as a reference standard to obtain the ΔCt value, and ΔΔCt values were calculated by subtracting the ΔCt of undifferentiated cyESCs as a calibrator.
Observation of the colony-specific expression of the germ cell marker genes
To determine whether the germ cell marker-gene expressions were specific to the colony-forming regions, we picked up the colony regions and the monolayer cell regions that were located around the colonies using glass capillary and treated with TRIzol reagent. The cDNA were prepared as above protocol. Comparison between the colonies and other regions were performed by real-time PCR analysis for VASA.
Immunofluorescent observation for differentiated cells
The differentiated cell cultures at day 8 were fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS) (4% PFA; pH 7.4). For coimmunofluorescence for VASA and SSEA-1, fixed cells were permiabilized in 0.2% Triton-X (Sigma-Aldrich) diluted in PBS (0.2% PBT) for 5 min, blocked by incubation in 5% skim milk (Sigma-Aldrich), diluted in PBS for 1 h, and then incubated with primary antibody for 1 h at room temperature. The primary antibody sets were anti-SSEA-1 mouse IgM monoclonal antibody (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and antihuman VASA goat polyclonal antibody (R&D Systems, Minneapolis, MN, USA) diluted 1/100 in 0.1% PBT. For immunofluorescence microscopy, the SSEA-1 and VASA antibody-treated samples were incubated for 1 h at room temperature with Texas red-conjugated antimouse IgM antibody (Santa Cruz Biotechnology) and FITC-conjugated antigoat IgG antibody (Santa Cruz Biotechnology) diluted 1/1000 in 0.1% PBT. Samples were then washed three times for 10 min each in PBS, and incubated in DAPI (Vector Laboratories, Ltd, Peterborough, UK) for detection of nuclear localization.
Western blot analysis
Cultured cells were collected by scraping and homogenized in SDS buffer (4% SDS, 125 mM Tis–glycine, 10% β-mercaptoethanol, 2% bromophenol blue in 30% glycerol). Aliquots were subjected to polyacrylamide gel electrophoresis in the presence of SDS (SDS–PAGE) followed by electrotransfer onto PVDF membrane (Hybond-P; Amersham Pharmacia Biotech, Buckinghamshire, UK). Molecular size was calibrated with Precision plus protein™ all blue standards (Bio-Rad; Mississauga, ON, Canada). The blotted membranes were blocked overnight with Block ace (Dainippon Sumitomo Pharma, Osaka, Japan), then reacted with objective primary antibody for overnight at 4°C. The antibodies were anti-OCT-4 rabbit polyclonal antibody (Santa Cruz Biotechnology), antihuman VASA goat polyclonal antibody (R&D Sstems), anti-STAT3 mouse monoclonal antibody (BD Biosciences) and antiphospho-STAT3 (Tyr705) rabbit polyclonal antibody (Cell Signaling Technology, Danverse, MA, USA). Antibody incubations and washes were performed in 0.1% Tween-20 in TBS throughout. Detection was realized by enhanced chemiluminescence with an ECL plus Western blotting detection system (Amersham Pharmacia Biotech) and horseradish peroxidase (HRP)-conjugated secondary antibodies (Santa Cruz Biotechnology). The lumino-labeled membranes were developed using X-Ray Film Processor. As a positive control, homogenized and SDS-treated cynomolgus monkey testis was used. For detection of OCT-4, the control sample was diluted at 1:5 and used for the blotting. For detection of VASA, the control sample was diluted at 1:1000 and used for the blotting. As a negative control, fibroblast cell of cynomolgus monkey was used.
Propidium iodide (PI) staining
Induced apoptosis was determined by PI staining. In brief, total cells were collected at days 2, 4, 6, and 8 of culture by TrypLE express treatment. Cells were washed twice and stained with PI/RNase Staining Buffer (BD Pharmingen, San Diego, CA, USA) for 15 min. Stained cells were analyzed with a FACS Caliber (Becton Dickinson, Mountain View, CA, USA). Fluorescent intensities were observed and the data were analyzed using Cell Quest (Becton Dickinson).
Statistical analysis of the data
Significant differences were detected by one-way analysis of variance (ANOVA) followed by Tukey-Kramer HSD tests or Student's t-test. A p-value of less than 0.05 was considered significant.
Results
The cyESCs rapidly differentiated when depriving bFGF from the culture medium. On day 3 to day 4 after differentiation induction, the cyESCs showed clear morphological change from the usual flat and compact colonies to elongated forms (Fig. 1A). These differentiated cells expressed early endodermal lineage markers Alpha fetoprotein (AFP) or early ectodermal markers NESTIN; on the other hand, pluripotent marker NANOG expression dramatically decreased (data not shown). When the differentiation is induced in LIF added condition, the cyESCs formed compact dome-like colonies at day 8 (Fig. 1A). Although the colony formations were also observed in the culture without LIF treatment, significantly higher number (about threefold) of colonies was observed in the LIF added condition (Fig. 1B). In mouse ESCs, LIF binds to cell surface receptor complex composed of LIF receptor and gp130, through which several signaling molecules including ERK and STAT3 are activated. The Stat activation is dependent upon tyrosine phosphorylation and the activated STAT3 can be observed by detecting phosphorylation at Tyr705 (Kristensen et al., 2005). Also in this study, the phosphorylation of STAT3 at Tyr705 residue was observed by LIF addition (Fig. 1C). To examine the effect of LIF addition to germ cell induction, we performed real-time PCR for primordial germ cell markers VASA, BLIMP-1, and an early germ cell/pluripotent cell marker OCT-4. The germ cell-specific gene expressions were noticeably upregulated in LIF+ condition. The increase in the gene expressions occurred between day 4 and day 8, approximately 50-fold increase in VASA expression and 3.5-fold increase in BLIMP-1 expression from undifferentiated cyESCs were observed at the day 8 time point. At day 8, VASA and OCT-4 proteins were detected in both LIF− and LIF+ conditions, and the same tendencies as in the mRNA expressions that VASA expression was observed approximately six times higher and OCT-4 expression was observed approximately 2.5 times higher in the cells induced from LIF+ conditions than that of the cells induced from LIF− conditions (Fig. 2B). Then we examined that the germ cell marker expression was specific in the colony structures, by comparing the expression of VASA between the colony regions and the monolayer regions. VASA expression in the colonies was approximately 30 times higher than that of the monolayer regions; it was determined that the putative germ cells were located in the colonies (Fig. 2C). Immunofluorescent staining showed germ cell-like characteristics of the colonies more clearly. VASA and SSEA-1 coexpressed cells were localized in the colonies (Fig. 3).
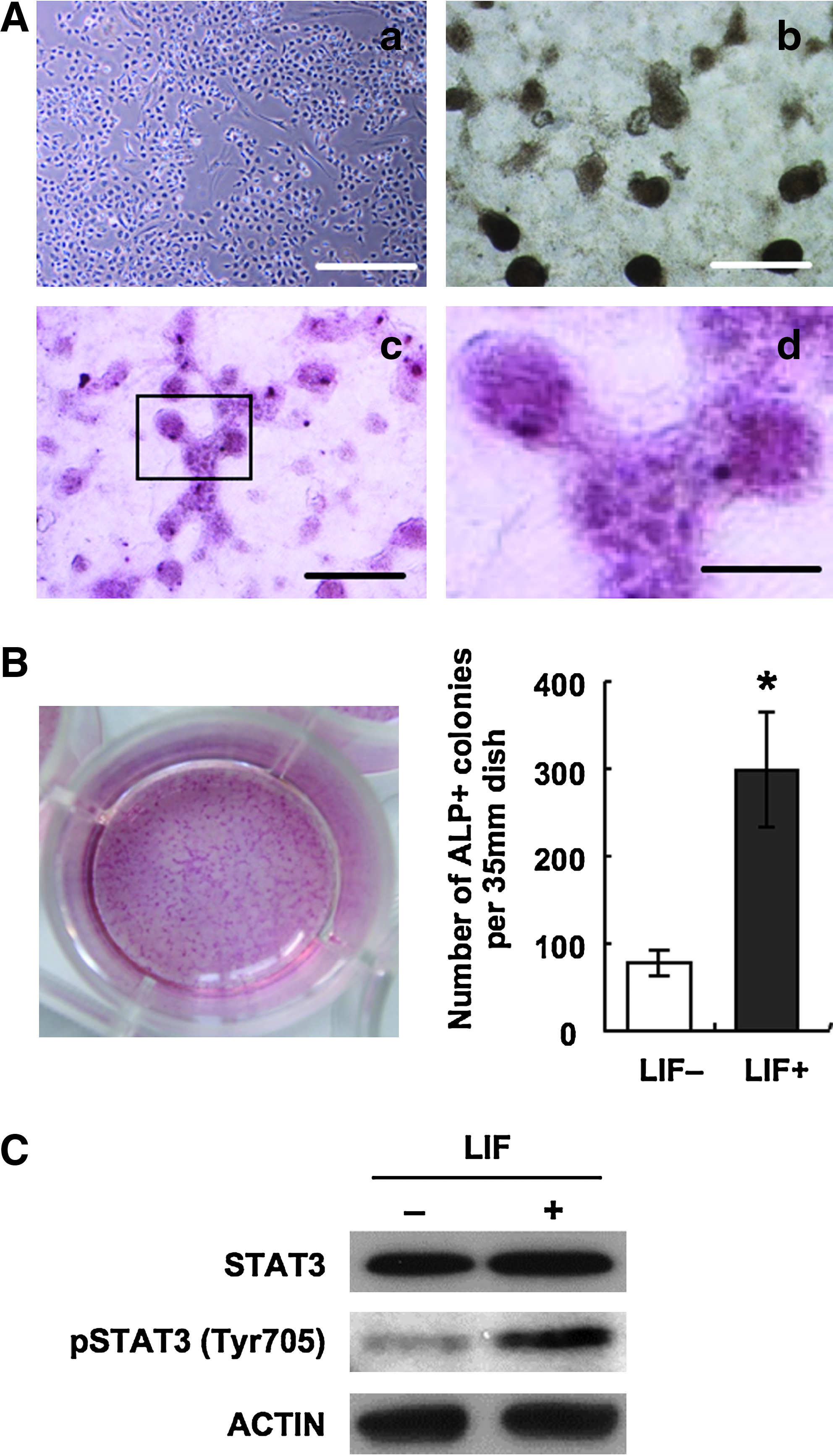
Colony formation in differentiation culture supplemented with LIF. (
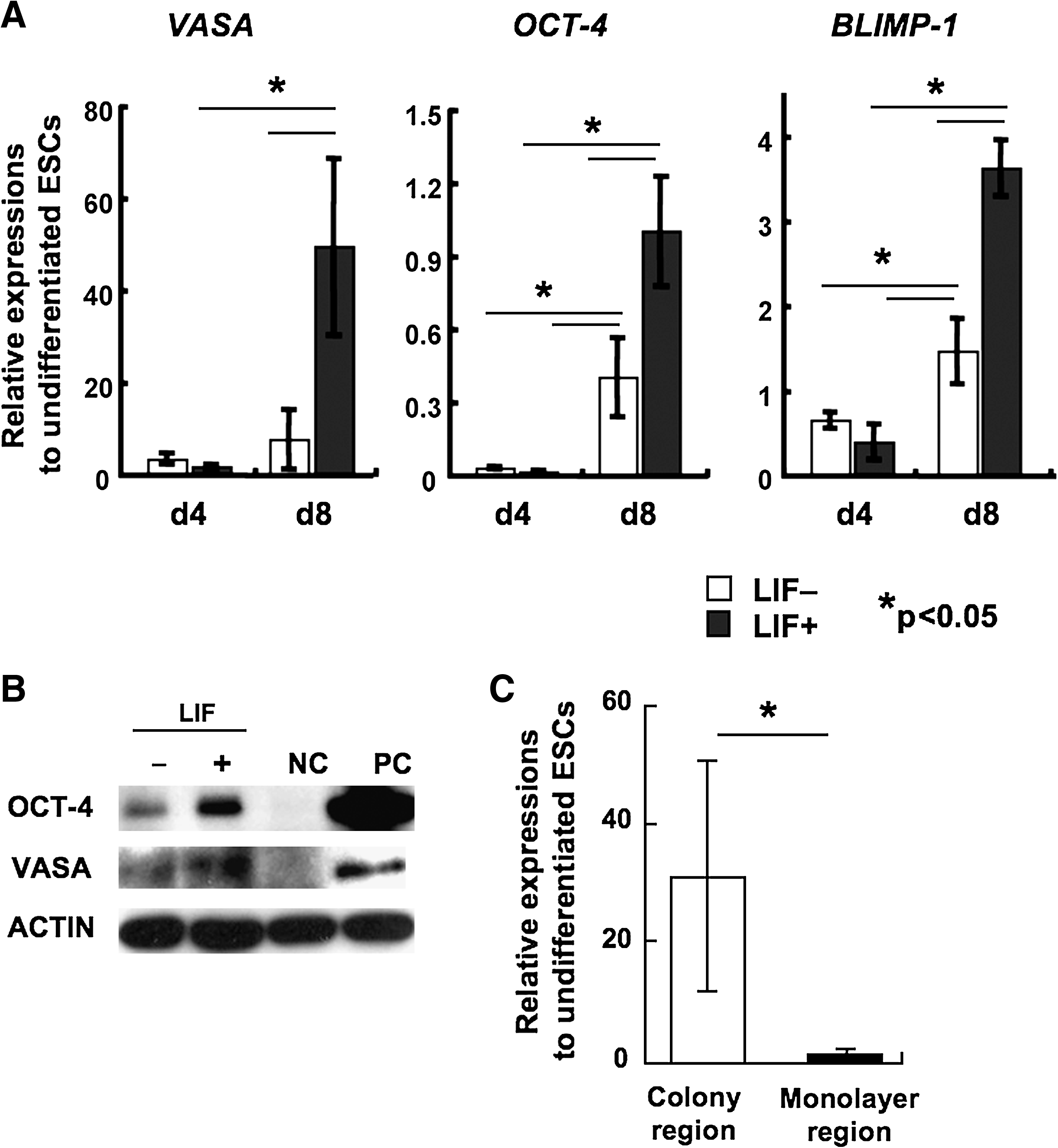
Real-time PCR based assays for germ cell-marker genes in the differentiated cells. (

Immunofluorescent staining for differentiated cells in LIF+ condition at day 8. (
In the next series of the experiment, we determined whether the upregulation of germ cell-marker gene expressions were results of activation of LIF signaling pathway using a JAK inhibitor, which interrupts JAK–STAT signal transduction. By adding the JAK inhibitor, colony formation was dramatically inhibited (Fig. 4A). The JAK–STAT interruption resulted in complete repression of VASA and BLIMP-1 expressions (Fig. 4B). The repressions of phosphorylation of STAT and the germ cell marker gene expressions were evidenced by Western blot analysis (Fig. 4C). These results demonstrate, at least in the present study, that LIF–STAT signal affects the germ cell-related gene expressions. Unexpectedly, OCT-4 mRNA expression was detected in the JAK inhibitor added cultures. Although detailed reason and mechanisms for the expression are unclear, the result that OCT-4 protein could not be detected at all suggests that the character did not represent functional expression of OCT-4, such as pluripotent cell remaining or germ cell differentiation.
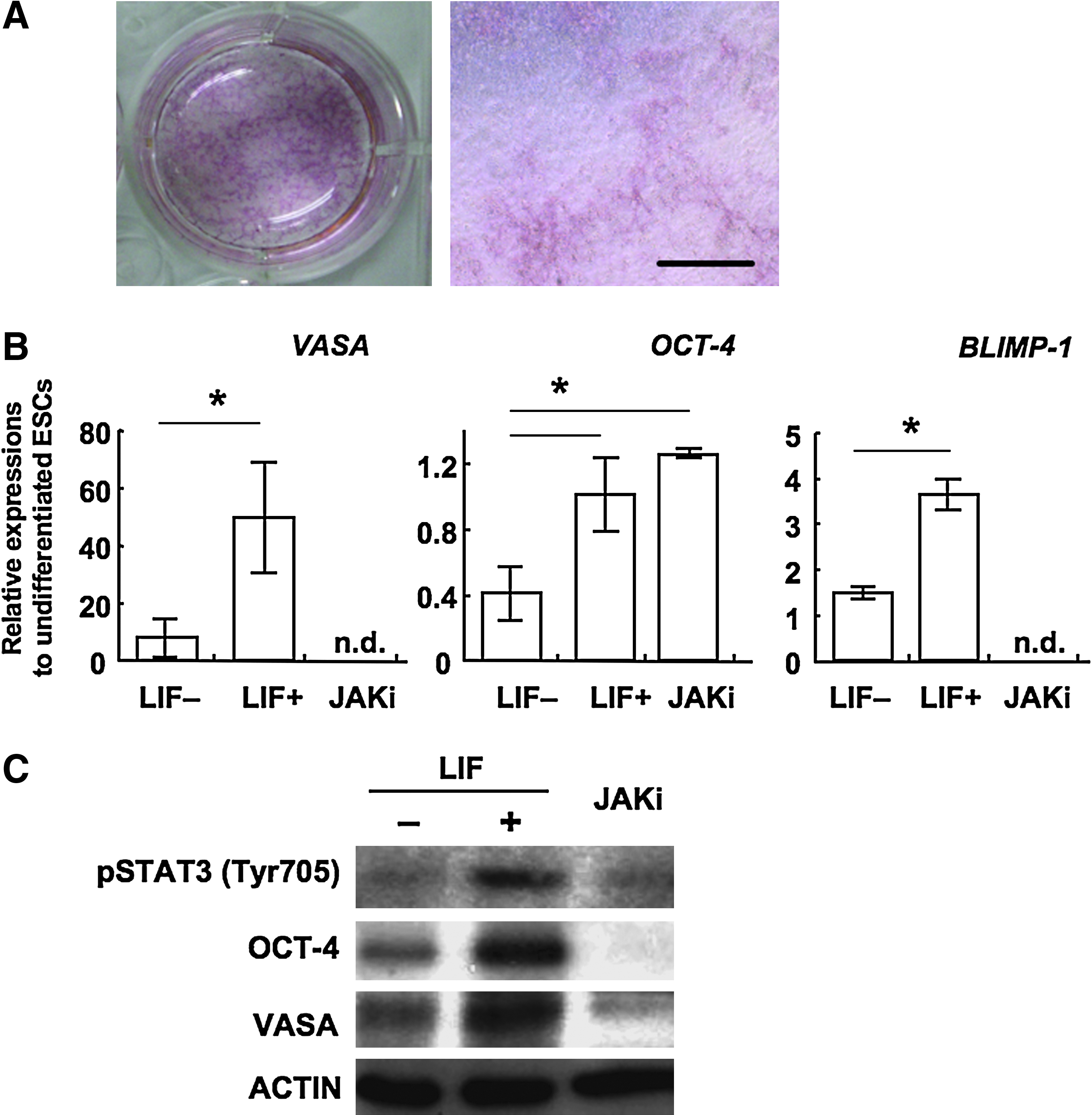
Inhibition experiment of LIF-STAT3 pathway in the differentiation culture. (
Koshimizu et al. (1996) described that the LIF prevents primordial germ cell death, and De Felici et al. (1992) also reported that the LIF promotes the survival rate of PGCs in in vitro culture. To examine the effects of LIF to the cell survival, we observed the rate of dead cells by FACS followed by PI staining of total cells. In the present study, LIF addition may not affect the rate of cell survival in the present systems (Fig. 5A). On the other hand, the percentage of ALP positive putative germ cells significantly increased in LIF added condition at day 6 and day 8 (Fig. 5B). From these results, it was suggested that LIF is not an inhibitor of cell death but is a promoting factor for the rate of germ cell-like cells, at least in vitro germ cell differentiation from ESCs. The previous two reports described that LIF inhibited spontaneous progression of meiosis of mouse PGCs, which occurred following isolation and in vitro culture (Chuma and Nakatsuji, 2001; Farini et al., 2005). Importantly, also in the present study, the repressions of meiosis specific gene expressions were observed. Meiosis related genes Dmc-1 and Scp-1 (Chuma and Nakatsuji., 2001) were detected at day 8 cells cultured in the LIF− condition, but no bands were detected in the cells cultured in the LIF+ condition (Fig. 5C). These results suggest the possibility that the major effect of LIF in cyESCs was to inhibit meiosis progression in germ cells or candidate of the cells, which may lead to increasing the rate of the cells expressing early germ cell-like phenotype.
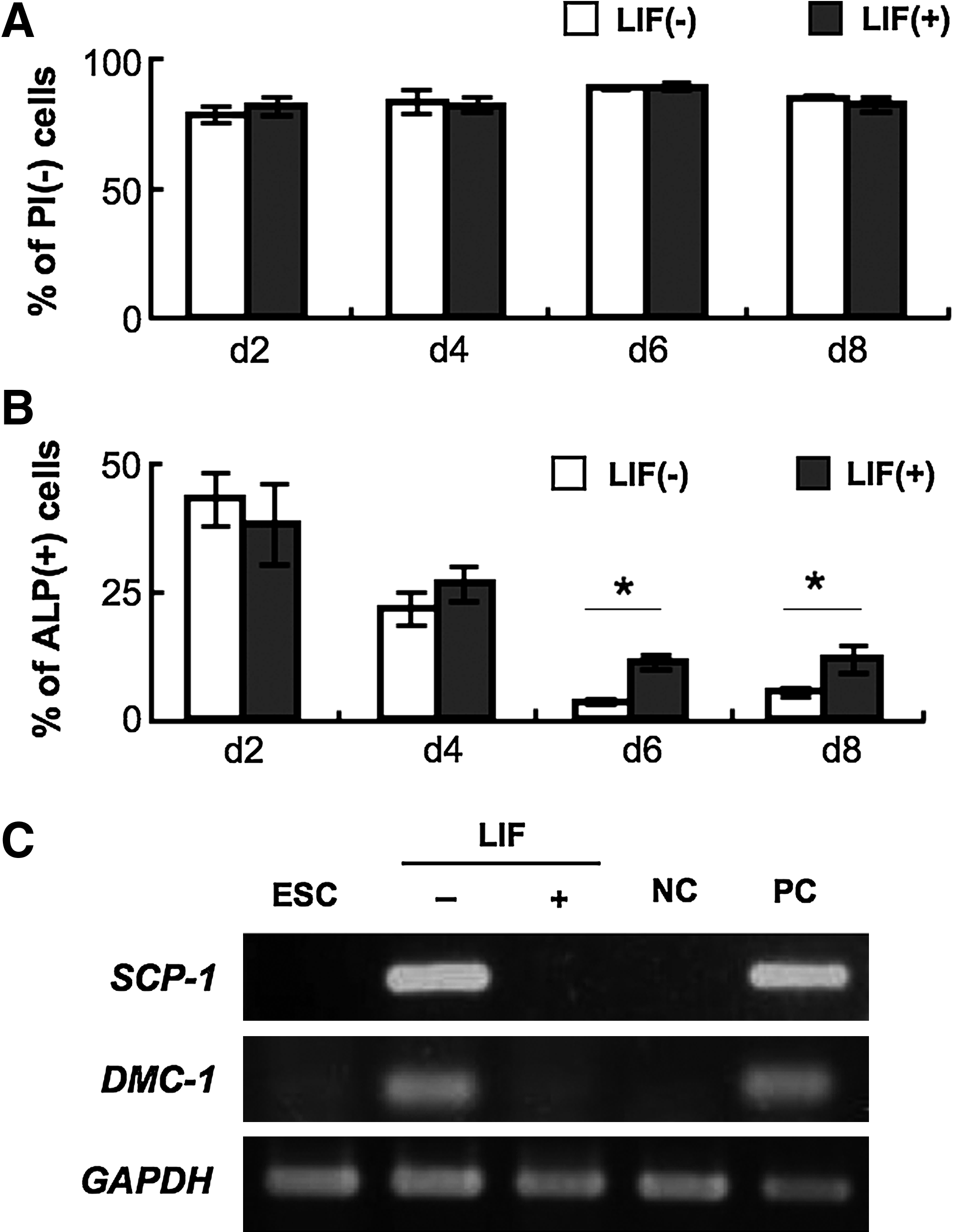
Effect of LIF in the induction of the germ cells. (
Discussion
To date, two groups reported germ cell differentiation from primate ESCs (Teramura et al., 2007; Yamauchi et al., 2009). However, the efficiency was so low and major improvements have been essential. We adopted the LIF as a factor for improving the germ cell differentiation, and examined the effects of LIF using cyESCs. Some functions of LIF to germ cells have already been reported in mouse PGCs, such as promotion of survival and cell growth, inhibition of meiosis progression, and inhibition of differentiation to mesodermal cells (Ohinata et al., 2009). If the same effects as above are determined in PGCs derived from cyESCs, it may directly lead to improvement of the protocols of germ cell induction from primate ESCs.
In this study, we observed ALP activity and germ cell marker genes VASA, BLIMP-1, OCT-4, and SSEA-1. ALP activity is a marker for premigratory to migratory PGCs (Lomelí et al., 2000). Vasa is an ATP-dependent RNA helicase and expressed in early germ cells at migratory to postmigratory stages (Anderson et al., 2007; Castrillon et al., 2000). Blimp-1 is a PR domain containing one with zinc-finger protein and an essential protein for PGC specification. It was also reported that Blimp-1-positive cells in postimplanted embryos were determined as germ cells (Saitou, 2009). Oct-4, which is a marker for pluripotent state of stem cells, is also a marker for early germ cell specification (Okamura et al., 2008). Lacham-Kaplan described that PGCs are the only cells expressing Oct-4 after postgastrulation in the mammalian development (Lacham-Kaplan, 2004). SSEA-1 is also a marker for early germ cells in primates (De Felici et al., 2004). These markers were differentially expressed by the stages of germ cells. Anderson et al. (2007) reported that, in human, expression of VASA is markedly increased between 9 and 14 weeks gestation when the gonocytes enter into meiosis in female. They also demonstrated that OCT-4 is not expressed in 14 to 19 weeks gestation (second trimester). On the other hand, SSEA-1 expression is first detected in PGCs before they migrate into the gonadal ridge, and continues until a postmigratory stage in males and is downregulated after the onset of meiosis in females. Thus, coexpression of VASA, SSEA-1, and OCT-4 was a guidepost for germ cells at the migratory stage to the postmigratory stage.
When inducing differentiation in the LIF+ condition, derivative cells of the cyESCs formed small dome-like colonies like the mouse ESCs at day 8. These colonies intensely expressed ALP, and expressed VASA, BLIMP-1, OCT-4, and SSEA-1. These results indicate that the induced cells from our study might correspond to migratory to postmigratory germ cells. Yamauchi et al. (2009) reported that spontaneously differentiated EBs derived from cyESCs contained germ cells at various developmental stages; SSEA-1 + /OCT-4 − , SSEA-1 + /OCT-4 + , VASA − /OCT-4 + , VASA + /OCT-4 + , and VASA + /OCT-4 − cells. In the present study, SSEA-1 + /OCT-4 + /VASA + cells were selectively expanded. Although significantly lower levels in comparison to the LIF+ condition, the germ cell-related gene expressions such as VASA, BLIMP-1, and OCT-4 mRNA were also detected in the differentiated cells in the LIF− condition. ESCs or the cells at early differentiation stages expressed LIF, and the endogenous JAK–STAT signal, due to either autocrine or paracrine LIF secretion, might affect the spontaneous differentiation of putative germ cells and result in the expression of germ cell-related genes.
LIF addition increased the rate of ALP-positive putative germ cells, although it did not affect the survival rate of total cells. In mice, it has been reported that the spontaneous progression to meiosis causes decreasing cell numbers of in vitro cultured PGCs (Ohkubo et al., 1996). Also in our study, meiosis-related gene expression was completely inhibited in LIF supplemented cultures. It might evidence to suggest that one important effect of LIF to improve germ cell differentiation may be inhibiting the germ cells to enter meiosis at abnormally early timing and increasing mitotic germ cells, at least in the germ cell formation from ESCs. Unfortunately, the presumptive germ cells induced in the LIF+ condition could not enter meiosis when they transfer to the LIF− condition or interrupt the LIF–STAT pathway by JAK inhibitor at day 8 (data not shown). One possible explanation is that the germ cells induced in the present study did not develop sufficiently to progress meiosis. Koubova et al. (2006) reported that retinoic acid is an essential factor for initiating meiosis, and SCF has also been demonstrated as an important factor for meiosis progression in female germ cells (Brieño et al., 2009; Koubova et al., 2006). Furthermore, Park et al. (2009) reported that coculture with fetal gonadal cells dramatically improved the number of the germ cells in the culture for inducing differentiation from human pluripotent cells. Addition of these factors at appropriate timing, the concentration might improve the stages of the putative germ cells.
Here, we elucidated that LIF can be an effective factor for germ cell differentiation from primate ESCs, although it was in the limited stages. Importantly, it has also become clearer that LIF supplementation alone may not be enough to obtain functional germ cells. We could not determine the target molecules of the LIF–STAT signaling in germ cell differentiation or could not elucidate of detailed mechanisms of LIF function in germ cell development in the present experiment. Specifications of these mechanisms and factors that can progress the stages may lead to establishing better culture systems for producing complete germ cells, and also help to enhance our knowledge in addressing infertility treatments.
Footnotes
Acknowledgments
We gratefully acknowledge Dr. Hirofumi Suemori, Institute for Frontier Medical Science, Kyoto University, for kindly providing primers to sequence the cynomolgus monkey BLIMP-1. We also thank Ms. Naomi Backes Kamimura, Department of Biology-Oriented Science and Technology, Kinki University, for English editing.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.