
Abstract
Abstract
Human embryonic stem cells (hESCs) are usually maintained in an undifferentiated state by coculture with mitomycin C-treated mouse embryonic fibroblasts (MEFs) as feeder cells in the presence of animal sera such as fetal bovine serum (FBS). Here, we use primary human amnion epithelial cells (hAECs) as feeder cells and human umbilical cord blood serum (CBS) as a replacement for FBS to support undifferentiated growth of hESCs. The 5∼10-fold higher expression levels of ES cell markers including FGF, Oct-4, Nanog, Sox-2, Rex, and TERT were found in hESCs grown on hAECs compared with that on MEFs as measured by quantitative real-time polymerase chain reaction (PCR). By immunofluorescence, the expresisons of Oct-4 and Nanog is also higher in cells grown on hAECs than those on MEFs. Importantly, the ES cells grown on hAECs exhibit normal karyotypes on passage 25, thus ruling out the possibility of transformation. Using flow cytometry analysis, we show that both the ES cells grown on hAECs and MEFs have the same cell cycle distribution pattern. Further, hESCs cultured on hAECs for at least 20 passages could differentiate into three germ layers via teratoma formation. In addition, chromatin immunoprecipitation assay revealed that histone H3 is highly acetylated, and H3 lysine (K) 4 is hypermethylated at the Nanog locus and the Oct-4 locus in hESCs grown on hAECs. Conversely, hESCs grown on MEFs show histone deacetylation and H3-K4 demethylation. Taken together, these results suggest that hAECs supplemented with 10% CBS are suitable for hESC culture, and that this method may prove to be valuable for use in future regenerative therapies.
Introduction
Here, we developed a clinically suitable protocol to use hAECs supplemented with human umbilical cord blood serum (CBS) to support undifferentiated growth of hESCs, and first provided evidence of histone modifications such as histone acetylation and H3-K4 methylation of the Nanog and Oct-4 gene in human ES cells grown on hAECs.
Materials and Methods
Preparation of hAECs
Human placentas and cord blood were collected with written and informed consent from healthy mothers with approval of the institutional ethics committee, after uncomplicated selective Cesarean section. Amnion membranes were prepared according to the methods previously described (Akle et al., 1981). Briefly, the membrane was mechanically peeled from chorines of placentas and washed with 1× phosphate-buffered saline (PBS). To relase hAECs, the amnion membrane were cut to yield 0.5–1.0 cm2 segments and digested with 0.25% trypsin/EDTA at 37°C for 45 min. The resulting cell suspension was seeded in a six-well plate in RPMI 1640 medium supplemented with 10% KO serum replacement (Gibco, Grand Island, NY, USA), streptomycin (100 U/mL), penicillin (100 U/mL), and glutamine (0.3 mg/mL), and incubated in a humidified tissue culture incubator containing 5% CO2 at 37°C. The hAECs grown to a density of 70–80% confluence were used as feeder layers for hESC culture.
Preparation of human cord blood serum
The cord blood was allowed to clot for 8–16 h at room temperature, centrifuged at 1000 × g for 30 min, and the cleared serum was collected. Sera from three donors were pooled to eliminate batch variations due to donor characteristics. Serum was filter sterilized by passing through a 0.22-μm filter. Complement was inactivated by keeping the serum at 56°C for 30 min.
Preparation of MEF feeder cells
These were done by Evans and Kaufman (1981).
Human ES cell culture
This study was approved by the International Peace Maternity and Child Health Hospital of Shanghai Jiaotong University committees on human subject research and ethics. The human ES cell line hHES1 (Wu et al., 2005) (gift from Dr. Wenjie Zhang) was passaged on mitotically inactivated MEFs. hESC culture medium consisted of KO-DMEM containing 10% KO serum replacement (or ES cell-qualified FBS), 2 mM L-glutamine, 0.1 mM nonessential amino acids, 0.1 mM 2-beta-mercaptoethanol, 10 ng/mL bFGF, 12 ng/mL hLIF, and penicillin (25 U/mL)–streptomycin (925 μg/mL) mixture. All reagents were purchased from Gibco. hESCs maintained on MEFs were dissociated by treatment of 0.05% trypsin–EDTA solution and plated onto MEFs or hAECs grown in hESC culture medium containing 10% human CBS instead of KO serum replacement or FBS. After small colonies emerged in the culture dish, single colonies were picked up with pipetteman (it is convenient to separate the colonies from the feeder cells) and cultured until they grew to large colonies.
Growth curve assay
The hAECs were cultured with CBS or KO serum replacement and 4000 cells per well were plated in 48-well culture dishes in 500 μL of grown medium; 50 μL of medium per well were added every 2 days. The number of cells for each well was evaluated after 7 days of culture.
Cell cycle distribution analysis
These were carried out by Lai et al. (2009).
RNA extraction and real-time qPCR analysis
Total RNAs were isolated from hESCs grown on hAECs or MEFs using RNeasy Mini Kit (Qiagen, Chataworth, CA, USA). Five hundred nanograms of total RNA from each sample was used in reverse transcription (RT) using iScript cDNA synthesis kit (Bio-Rad, Hercules, CA, USA). Real-time RT-qPCR was carried out on cDNA using IQ SYBR Green (Bio-Rad) using Mastercycler ep realplex (Germany). All reactions were performed in a 25-μL volume. Primers sequences were listed in Table 1. PCR was performed as previously reported (Lai et al., 2009).
Western blot analyses
The hESCs clones were pick up from MEFs or hAECs and pooled and homogenized in the sample buffer. Total proteins were measured using the BCA kit (Pierce, Gaithersburg, MD, USA) according to the manufacturer's protocol. Twenty micrograms of protein was separated by SDS–PAGE and transferred to nitrocellulose membrane. Then the membrane was incubated with the primary antibody against LIF or β-actin (Rabbit antihuman/mouse, 1:200, BoShiDe, Wuhan, China; or Rabbit antihuman/mouse, 1:1000, Cell Signaling, Danvers, MA, USA) at room temperature overnight. This was followed by incubation with peroxidase-linked goat antirabbit-IgG (1:1000, Santa Cruz Biotechnology, Santa Cruz, CA, USA) at room temperature for 1 h; then it was developed with a chemiluminescence reagent (Perkin-Elmer Life Science, Norwalk, CT, USA) and analyzed using the ChemiImager Imaging System (G: BOX SYNGENE, Gene Company Limited, Hong Kong). The MEFs and hAECs were used as positive controls.
Karyotype analysis
Chromosome analysis of hESCs grown on MEF or hAECs were performed using the G-band methods.
Immunofluorescence staining
hESCs cultured on hAECs or MEFs were fixed with 4% paraformaldehyde for 15 to 20 min at room temperature, and then washed twice (10 min each) with 1 × PBS. Cells were permeabilized with 0.1% Triton X-100/PBS for 10 min at room temperature, and then washed twice with 1 × PBS. The cells were then blocked with blocking solution for 30 min and incubated with the anti-Oct-4 (rabbit antihuman 1:200, Chemicon, Temecula, CA, USA) or anti-Nanog antibody (rabbit antihuman 1:200, Chemicon) for 1 h at room temperature. Cells were then washed three times with 1 × PBS, and probed with fluorescein isothiocyanate (FITC)-tagged antirabbit (1:200, Jackson Immunoresearch, West Grover, PA, USA) antibody. Fluorescence images were taken using a Leica DMI3000 microscope.
Teratoma formation
All animal procedures were conducted following experimental protocals previously approved by Shanghai Jiaotong University with Institutional Animal Care. After 20 passages on hAECs, 1 × 106 hESCs were injected into the hind leg of severe combined immunodeficient (SCID) mice. After 7–8 weeks, tumors were processed in paraffin and analyzed after hematoxylin and eosin staining. The same numbers of hAECs were injected into the hind leg of SCID mice as negative controls for tumor formation.
Chromatin immunoprecipitation (ChIP) assays
The ChIP assays were performed using antiacetylated histone H3 antibodies (Catalog no. 06599; Upstate, Waltham, MA, USA) and antitrimethylated H3-K4 (UniProt Number: Q16695; Upstate) according to the manufacturer's protocals. Normal rabbit IgG (Catalog no. 12-370; Upstate) was used as a negative control to verify immunoprecipitation specificity. In brief, cells were washed twice with PBS and treated with 1% formaldehyde for 10 min at room temperature to form crosslinked chromatin. Samples were sonicated on ice until fragments became 200–1000 bp in size and incubated with antibodies at 4°C overnight. The PCR amplification was performed under the following conditions: 95°C for 10 min, 32 cycles of 94°C for 30 sec, 60°C for 30 sec, and 72°C for 30 sec, final extension 72°C for 10 min. Primers were designed as follows: Nanog forward, 5′-GTTCTGTTGCTCGGTTTTCT-3′; reverse, 5′- TCCCGTCTACCAGTCTCACC-3′ (Sequence coverage relative to TSS, −173 to −79), Oct-4 forward, 5′-GAGGATGGCAAGCTGAGAAA −3′; Oct-4 reverse, 5′- CTCAATCCCCAGGACAGAAC-3′ (Sequence coverage relative to TSS, −2546 to −2379) (Hattori et al., 2007). The amount of each product was evaluated with an ethidium bromide stained gel-image using BandScan software 4.30.
Statistical analysis
Assays were performed three times each in triplicate and results are reported as mean ± SD. The significance of differences in mean values was determined using Student's t-test.
Results
Evaluation of hAECs as feeder layers for hESC culture
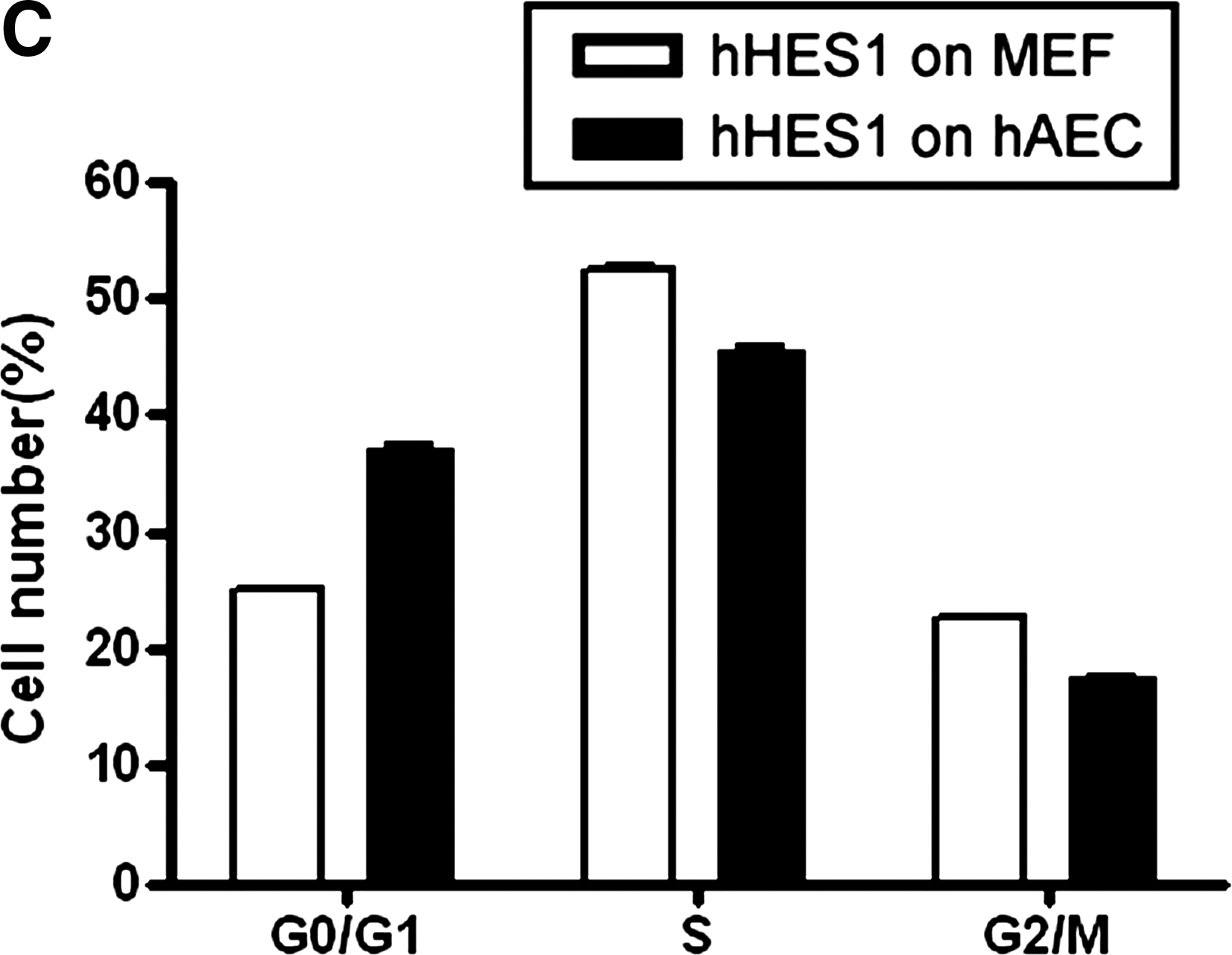
To examine whether hAECs have the potential to support the growth of hESCs, we collected primary hAECs from human placenta and cultured them in the hESC culture medium supplemented with 10% CBS instead of KO replacement serum or FBS. These hAECs can grow in CBS or KO replacement serum, but they did not proliferate under these culture conditions (Fig. 1A and E, p < 0.01), although they were not treated with mitomycin C or with gamma irradiation (Lai et al., 2009). The MEF cells were treated with mitomycin C (Fig. 1C). hESCs were grown on hAECs or MEFs, and the morphology of ES colonies were compared. As shown in Figure 1, the hESC colonies maintained on hAECs (Fig. 1B) are compact multicellular round colonies, similar to those grown on MEFs (Fig. 1D). hESCs grown on hAECs and MEFs also exhibit similar cell cycle distribution. As shown in Figure 2AB, the cell cycle profile of the hESCs on hAECs is the same as those grown on MEFs (Fig. 2C, p > 0.05). To exclude the feeder's contamination, we examined the LIF expression in hESCs on MEF or hAECs. As shown in Figure 3, the LIF expression level in hESCs on MEF or hAECs is almost negligible. To rule out the possibility of transformation of ES cells, we performed karyotype analyses. Results show that the hESCs grown on hAECs (Fig. 4A) or MEFs (Fig. 4B) have a normal 46XY karyotype on passage 25.
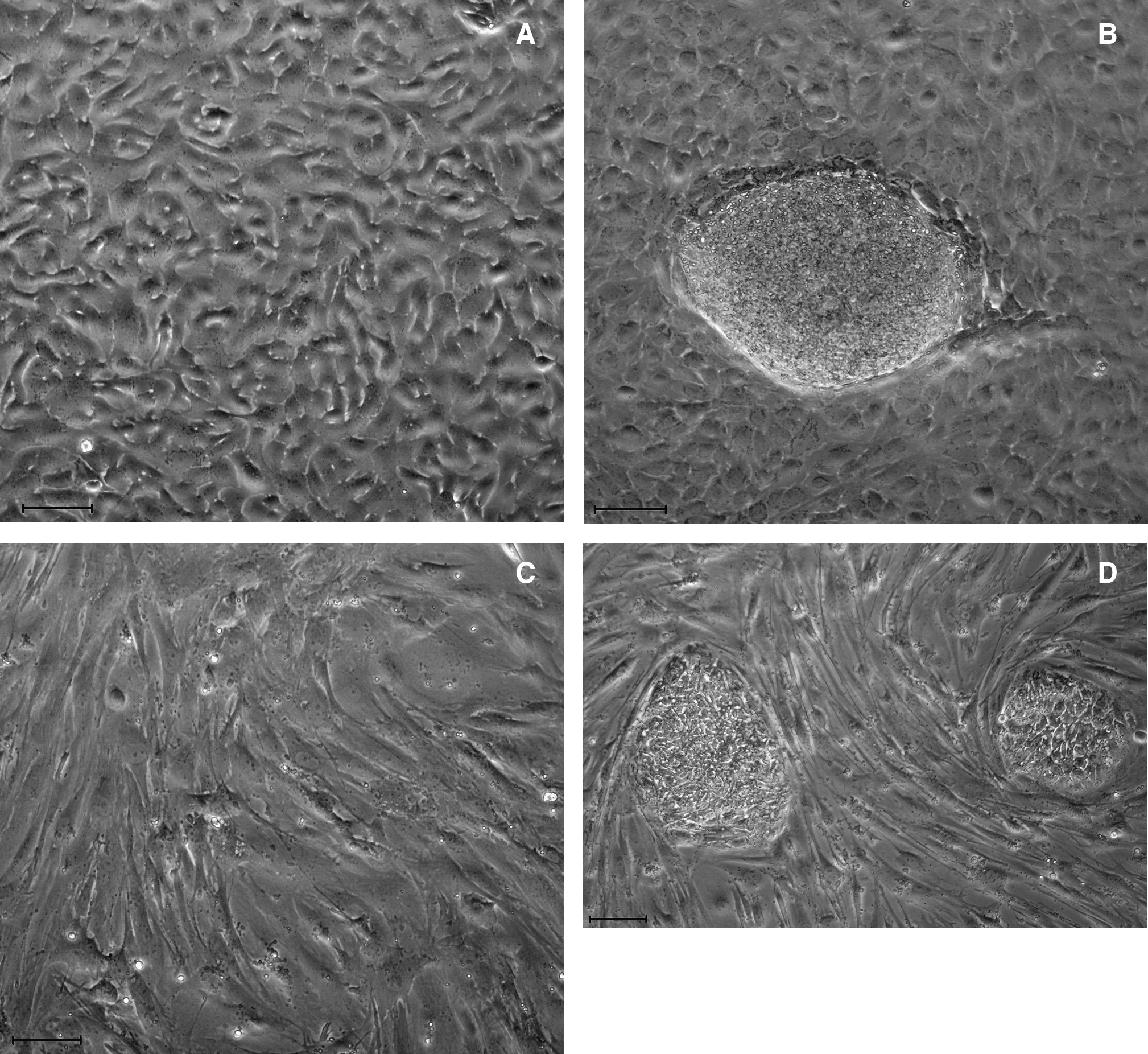
Morphology of hESCs. (

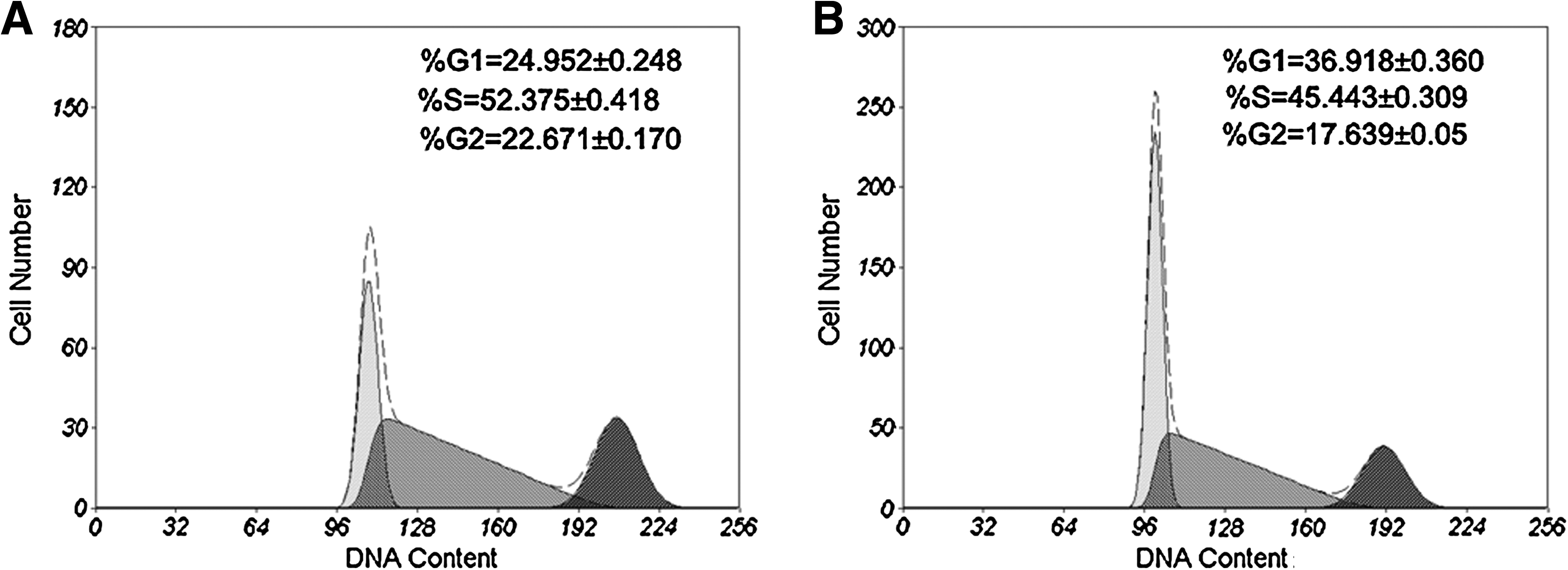
Cell cycle analysis of hESCs. (
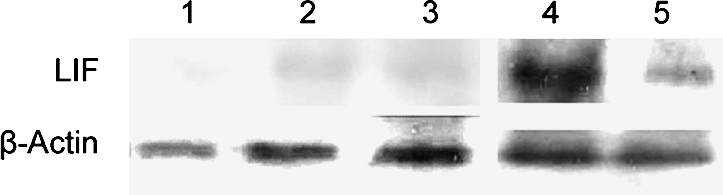
LIF expression in hESCs grown on hAECs or MEFs by Western blot analysis. The LIF expression levels in MEF and hAECs are as positive controls, LIF expression in hESCs on MEF or hAECs is almost negligible. (1: hESCs without feeder. 2: hESCs on MEFs. 3: hESCs on hAECs. 4: hAECs. 5: MEFs.)
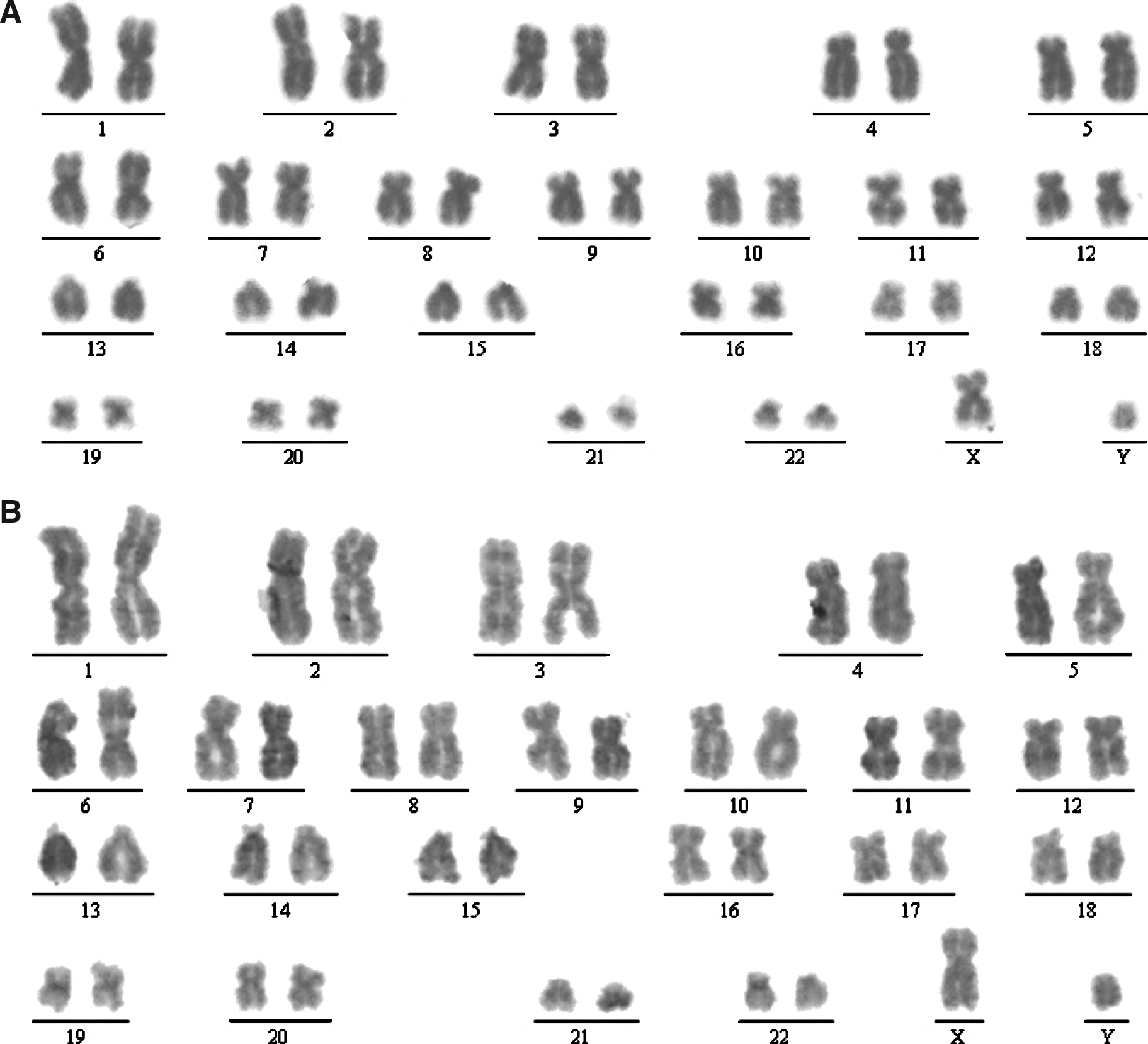
Karyotype analysis of hESCs grown on hAECs (
ES marker expression of hESCs grown on hAECs
To further evaluate the growth of hESCs on hAECs, we analyzed the expression of ES cell marker genes. Results from our qPCR experiments show that these cells not only maintain the expression of the markers tested, but the expression levels of these markers including FGF, Nanog, Rex, Tert, Oct-4, and Sox-2, are 6- to 10-fold higher than those grown on MEFs (Fig. 5). In addition, we examined the expression of Oct-4 and Nanog by immunofluorescence. Results show that hESCs grown on hAECs have stronger staining patterns for Oct-4 (Fig. 6, compare A to B) and Nanog (Fig. 6, compare C to D) compared to those on MEFs.

Expression of FGF, Nanog, Rex, Tert, Oct-4, and Sox-2 in hESCs grown on hAECs or MEFs by QRT-PCR (p < 0.01).

Immunostaining of hESCs cultured on hAECs (
The developmental potential of hESCs maintained on hAECs
To investigate the pluripotent potential of hESCs grown on hAECs, we injected hESCs into hind legs of SCID mice. The hESCs grown on hAECs results in the formation of typical immature teratomas containing all three main germ layers, including immature tissues of neural tubes (Fig. 7A), muscles (Fig. 7B) and glands (Fig. 7C), as identified by histologic analysis. As expected, injection of hAECs alone did not produce tumors (data not shown).

Histology of hESCs that developed into tumors when transplanted into SCID mice. Immature teratoma containing immature tissues of neural tubes (
Status of histone H3 acetylation and H3-K4 methylation of Nanog and Oct-4 in hESCs grown on hAECs
To test the distinct epigenetic status between Nanog and Oct-4, we compared the status of histone H3 acetylation and H3-K4 methylation of Nanog locus and Oct-4 locus in human ES cell line hHES1 grown on hAECs and MEFs by ChIP assay (Fig. 8). In human ES cells grown on hAECs, histones H3 of the Nanog gene and Oct-4 gene were hyperacetylated, whereas acetylation of H3 in ES cells grown on MEFs was minimal (Fig. 8A). We also investigated H3-K4 tri-methylation status. Similarly, H3-K4 trimethylation was high at the Nanog gene and Oct-4 gene in ES cells grown on hAECs, but was low in ES cells grown on MEFs (Fig. 8B). Thus, in human ES cells grown on hAECs, histone modifications such as histone acetylation and H3-K4 methylation of Nanog and Oct-4 gene are distinct from ES cells grown on MEFs.
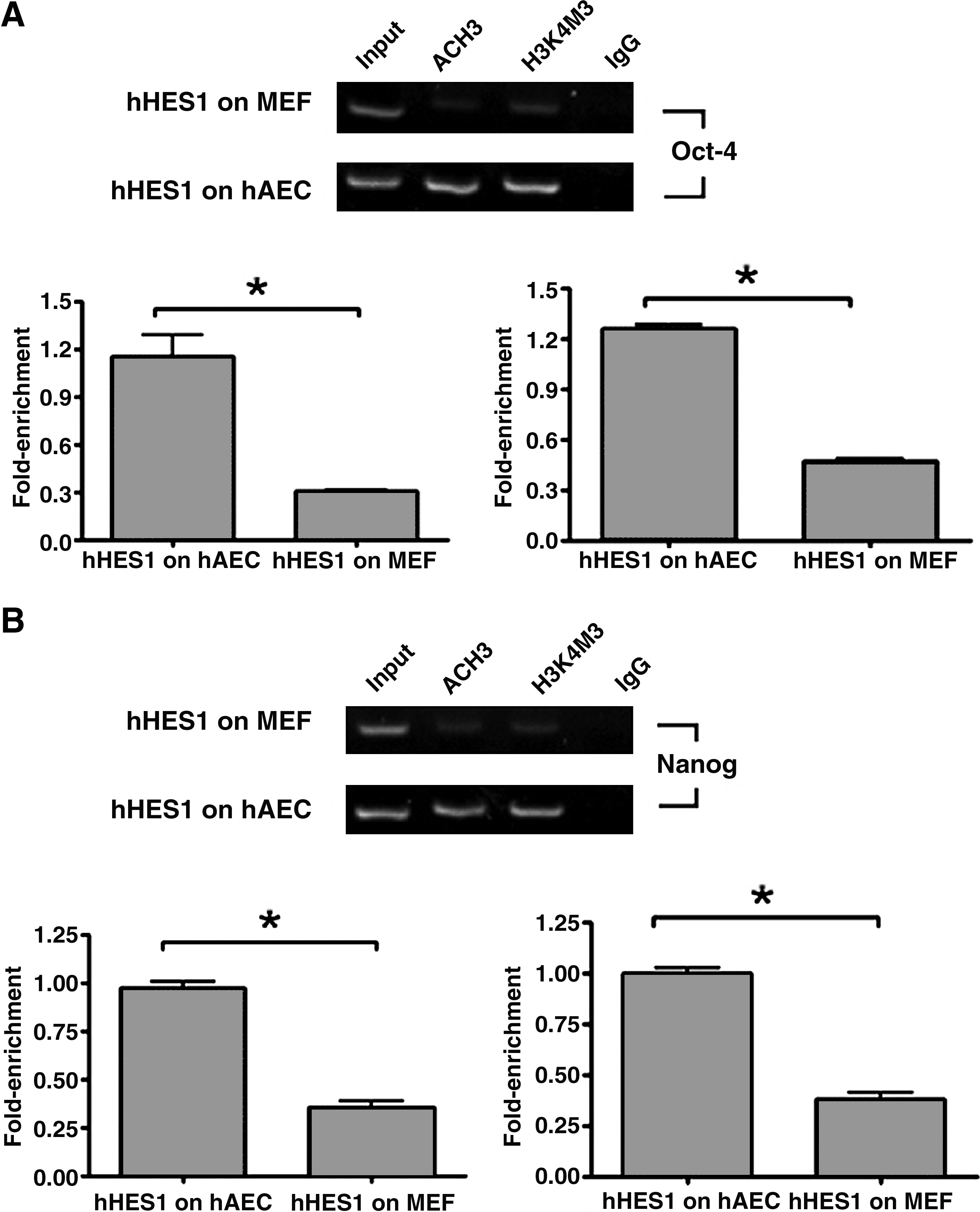
Histone H3 acetylation and methylation on Nanog and Oct-4 are different between hESCs grown on hAECs and MEFs. The human ES cell line hHES1 grown on hAECs and MEFs were subjected to ChIP assay using antibodies against acetylated histone H3 (AcH3) and antibodies against trimethylated H3-K4 (H3K4m3). Normal rabbit IgG (IgG) was used as a negative control for the specificity of the immunoprecipitation (IP). As a positive control, aliquots (0.56%) of chromatin fragments were also subjected to PCR without IP (input). Intensity of the bands for AcH3 and H3K4m3 were normalized by input, indicated as fold enrichment, and is shown by a histogram in the bottom panel. Data represent the mean ± SE in triplicate. *p < 0.05 (Student's t-test). (
Discussion
To require the clinical-grade human ES cells, it is important to derivate of human cell-based feeder and animal origin products to avoid the risks of zoonosis. Many types of human cells have been used as feeder layers. These include human foreskin, fetal muscles, fetal skin, adult fallopian tube epithelial, adult uterine endometrium, adult marrow stromal cells, but most of these are difficult to acquire due to ethical and practical limitations (Cheng et al., 2003; Hovatta et al., 2003; Inzunza et al., 2005; Lee et al., 2005; Richards et al., 2003). Although feeder-free culture of human ES cells are reported, the chromosomal instabilities of human ES cells happened during the long time culture (Catalina et al., 2008). In contrast, the human placental amnion is abundantly available as routinely discarded tissues, and there is no ethical complications. Thus, the hAECs could feasibly be used as an suitable source for generating feeder layers for hESC culture.
Another important reason for the hAECs as feeder cells is that human amnion epithelial cells may secrete some proteins into culture medium, or they may provide some membrane proteins for undifferentiated growth of ES cells. Koizumi et al. (2000) reported eight growth factors [epidermal growth factor, keratinocyte growth factor, hepatocyte growth factor, basic fibroblast growth factor, transforming growth factor (TGF)-α, TGF-β1, TGF-β2, and TGF-β3] and two growth receptors (keratinocyte growth factor receptor and hepatocyte growth factor receptor) in preserved human amniotic membrane (Koizumi et al., 2000). Recently, Miki et al. (2005) reported that the amniotic epithelial cells express Oct-4, Nanog, and retain the stem cell characteristics. Chen et al. (2007) reported that hAECs can be used as a human feeder layer equivalent for more effective ex vivo expansion of adult epithelial stem cells from the human limbus. Ellerström et al. (2006) developed a xeno-free medium supplemented with human adult serum to supports long-term culture of hESCs in an undifferentiated state. In our system, the CBS was used to avoid xeno-contamination. CBS is a rich source of different cytokines including a variety of growth factors like insulin-like growth factor (IGF), TGF-β, and vascular endothelial growth factor (VEGF). Lam et al. (2001) have used cord blood plasma for the expansion of cord blood HSC. The cord blood plasma has also been used to culture T cells for adoptive immunotherapy (Kim et al., 2005). Shetty et al. (2007) reported that human umbilical cord blood serum can replace fetal bovine serum in the culture of mesenchymal stem cells.
In this context, we report that the human amnion epithelial cells as a feeder layer supplemented with 10% CBS supports hESCs in an undifferentiated state and maintains the pluripotency of ES cells. We also found that this culture system could support the growth of other human ES cell lines, such as HUES-1 and HUES-2, in an undifferentiated state (data not shown). One interesting and useful observation from our studies is that human ES colonies grown on hAECs undergo a more compact multicellular state compared with ES colonies on MEFs. More importantly, both the real-time qPCR and immunofluorescence analysis showed that human ES colonies grown on hAECs express higher levels of stem cell genes, such as Nanog, Oct-4, FGF, Rex, Tert, and Sox-2. The Nanog and Oct-4 transcription factors are key molcules of self-renewal of ES cells. Oct-4 is a member of the POU family of transcription factors, and Nanog is the most recently described homedomain gene. It has been shown that Nanog function requires the continued presence of Oct-4, and together they maintain stem cell pluripotency and self-renewal (Deb-Rinker et al., 2005). Thus, these factors produced by amnion epithelial cells or cord blood may orchestrate elegant developmental steps in a regulated fashion and offer the appropriate microenvironment in regulating embryonic stem cell fate to proliferate and self-renew or to enter a multistep differentiation pathway.
Recently, the epigenetic reprogramming of histone acetylation and H3 lysine 4 (H3K4) methylation of the Nanog gene and Oct-4 gene in ES cells have been reported. Histones acetylation and of histone H3 lysine 4 (H3K4) methylation may affect gene activation by relaxing chromatin (Hayashi et al., 2005). These epigenetic marks in mouse ES cells were different from that in trophoblast stem (TS) cells, which histone H3 are highly acetylated, and H3 lysine (K) 4 is hypermethylated at the Nanog region and Oct-4 region in ES cells (Hattori et al., 2007). Epigenetic reprogramming of Oct-4 and Nanog regulatory locus has been elicited by exposure of 293T cells to human embryonal carcinoma (EC) cell extract. Lysines K4, K9, and K27 of histone H3 on Oct-4 and Nanog indicate that primary chromatin remodeling determinants are acetylation of H3K9 and demethylation of dimethylated H3K9. H3K4 remains di- and trimethylated (Freberg et al., 2007). We first examined histone modifications in the human ES cells in different feeder layers. In human ES cells grown on hAECs, histones H3 of the Nanog gene and Oct-4 gene were hyperacetylated, whereas acetylation of H3 in ES cells grown on MEFs was minimal. Similarly, H3-K4 trimethylation was high at the Nanog gene and Oct-4 gene in ES cells grown on hAECs, but was low in ES cells grown on MEFs. Thus, histone modifications such as histone acetylation and H3-K4 methylation of Nanog and Oct-4 gene are different between ES cells on hAECs and ES cells grown on MEFs. Our results provide evidence of reprogramming of DNA methylation and histone modification on the Nanog promoter and OCT4 gene in human ES cells grown on different feeder layers. Thus, the mechanisms underlying the hAECs mainting human ES cell pluripotency are composed of the distinct epigenetic reprogramming of Oct4 and Nanog genes DNA methylation and histone modifications.
In conclusion, we have developed an ideal strategy for propagating hESCs on hAECs supplemented with CBS. We are derivating new human ES cell lines that have very limited contact with animal products and thus are better candidates for future regenerative therapies.
Footnotes
Acknowledgments
We thank Dr. Cao yulin, Dr. Zhang Wenjie, and Tang Zhengya for helpful experimental suggestions concerning human ES cultures. We especially acknowledge the support of Melton Laboratory of Harvard University (HHMI). We thank Dr. Yingqun Huang for critically reviewing the manuscript. The funding for this work was supported by Science and Technology Commission of Shanghai Municipality 2007 Shanghai pujiang project to Dongmei Lai (07pj14090).
Author Disclosure Statement
No potential conflicts of interest exist.