
Abstract
Abstract
Universal recipients in the G2 phase of mitotic cell cycle (preactivated oocytes, zygotes, blastomeres) accept embryonic nuclei in all the stages of their cell cycle. To test if recipients in the G2 of meiotic cycle (immature oocytes) are universal recipients, mouse germinal vesicle (GV) oocytes were enucleated and reconstructed with blastomere nuclei in the G1, S, or G2 stages. Analysis of their maturation has shown that about 30% of the G1 nuclei and 60% of G2 nuclei allow for normal metaphase II (MII), both in the oocytes with and without the first polar body (1st PB). Among oocytes reconstructed with the S phase nuclei, only 8% or less have normal MII, although 75% of them extrude 1st PB. No phase of donor cell cycle prevented the abnormal acceleration of 1st PB extrusion, found in reconstructed GV oocytes. In conclusion, enucleated GV oocytes are not universal recipients of embryonic nuclei, because they do not accept the S donors. However, both the G1 and G2 donor nuclei can be reprogrammed in the GV oocyte cytoplasm.
Introduction
It has been proven that an ovine or bovine preactivated oocyte (i.e., an interphase recipient) can harbor donor embryonic nuclei at all stages of the cell cycle (a “universal cytoplast,” Campbell et al., 1993; “The Universal Recipient,” Campbell et al., 1994; for review, see Campbell et al., 1996; Modlinski et al., 2001). However, when other interphase recipients like mouse zygotes or blastomeres from the two-cell stage in the G1 or G2 were used, their G2 phase of the cell cycle supported the development of reconstituted cybrids receiving embryonic nuclei better than their G1 phase, as can be inferred from the cleavage rate (Smith et al., 1990) and blastocyst formation. When transferred to late (G2) zygotes, late (G2) two-cell embryonic nuclei yielded 63% of blastocysts, mid (S) two-cell nuclei 49%, and early (G1) two-cell nuclei 66% (Smith et al., 1988). In early (G1) zygotes, only 2.3, 18.9, and 20.0% of embryos reconstituted with G1, S, or G2 blastomere nuclei, respectively, developed into blastocysts (Smith et al., 1988).
The recipients routinely used in the majority of protocols for cloning mammals by nuclear transfer are oocytes in metaphase of the second meiotic division (MII oocytes). However, immature mouse oocytes in meiotic prophase (germinal vesicle oocytes, GV oocytes) have been tested as alternative recipients of embryonic (Grabarek et al., 2004) or somatic nuclei (Bui et al., 2008; Chang et al., 2004; Mohammed et al., 2008; Nan et al., 2007; Polanski et al., 2005), and their developmental potential as recipients of embryonic nuclei has been confirmed (Mohammed et al., 2008). The GV of an immature oocyte is an interphase nucleus that has been arrested during the diploten (dictyate) stage of meiotic prophase, and its cell cycle stage is G2 (Lima-de-Faria and Borum, 1962; Peters et al., 1962). As a recipient in the G2, the GV oocyte might be thought a universal recipient. Both embryonic and somatic donors in the G2 phase of the cell cycle were often used to reconstruct enucleated GV oocytes (Chang et al., 2004; Grabarek et al., 2004; Mohammed et al., 2008; Polanski et al., 2005). The G0/G1 cumulus cell nuclei were injected into enucleated GV oocytes to test their reprogramming ability (Bui et al., 2008). When the G0/G1 cumulus cell nuclei were compared with the G2 spermatocyte nuclei, both of them introduced into the GV oocytes 1 to 2 h before enucleation, the conclusion has been drawn that the G2 nuclei only slightly improve maturation of the GV cytoplasts compared with G0/G1 nuclei (Polanski et al., 2005). However, it is difficult to conclude how the cell cycle stage of donor nucleus may affect the meiotic maturation of reconstructed oocytes, because the reports differ regarding the type of donor cell, its cell cycle stage, as well as the experimental protocol used.
Given these data and those on the late G2 zygotes reconstructed with blastomeres (Smith et al., 1988), we have aimed at finding if the GV ooplast is the universal recipient. To achieve this, the blastomere nuclei in the G1, S, or G2 stages of the cell cycle were introduced into enucleated GV oocytes and their maturation was compared in detail, within the same experimental system.
Materials and Methods
Reagents and culture media
All inorganic and organic compounds used in this study were purchased from Sigma (Sigma Chemical Co., St. Louis, MO, USA), unless otherwise stated. All media were prepared fresh and sterilized by filtering through a 0.22-μm filter (Acrodisc; Pall Gelman Laboratory, Ann Arbor, MI, USA).
Animals
Mice originated from the mouse colony, which is bred in the Department of Experimental Embryology. Mice were kept under a 12-h light/12-h dark cycle starting at 7 a.m. Food (Labofeed H, Poland) and water were available ad libitum. Donor females were killed by cervical dislocation.
Recipient cells
For collection of germinal vesicle (GV) oocytes, 6- to 8-week-old F1 (C57Bl10xCBA/H) females were injected with 7.5 IU of pregnant mare's serum gonadotrophin (PMSG; Folligon, Intervet, Holland). Ovaries were removed from the donor females 44–48 h after PMSG injection. Antral follicles were punctured by 30-gauge needles, and cumulus–GV oocyte complexes were released into HEPES-buffered manipulation M2 medium (Fulton and Whittingham, 1978; Whittingham, 1971) containing 0.2 mM dibutyryl cyclic adenosine 3′,5′-monophosphate (dbcAMP) to inhibit GV breakdown. The oocytes were stripped of cumulus cells in the above medium by repeated aspiration into a glass pipette, the tip diameter of which was slightly larger than the diameter of an oocyte and cumulus-free GV oocytes were selected for enucleation or culture.
Collection of cleaving embryos
Embryos were collected from 3-month-old MIZ (Swiss albino) female mice. Females were superovulated by injection of 7.5 IU of PMSG (Folligon) followed by 7.5 IU of human chorionic gonadotrophin (hCG, Chorulon, Intervet, Holland) 48 h later and were mated with F1 (C57Bl10xCBA/H) males. The embryos that served as a source of the G1 and S blastomeres were collected from the oviducts 29–30, 48–49, and 59–60 h after hCG injection, as late one-cell, two-cell, and four-cell stage embryos, respectively. For G2 blastomeres, two-cell, four-cell, and eight-cell embryos were flushed from the oviducts 44–46, 56–58, and 68–70 h after hCG injection, respectively (Grabarek et al., 2004).
Staging the cell cycle
Late one-cell, two-cell, and four-cell stage embryos were cultured in KSOM medium (Specialty Media, Phillipsburg, NJ) at 37°C in the atmosphere of 5% CO2 in air and were monitored every 15 min for cleavage. The embryos were either used immediately after cleavage, as a source of G1 phase blastomeres, or after next 3–4 h of culture, as a source of S phase blastomeres. Because nuclear transfer procedure took 1.5–2 h, then the introduced nuclei were likely to be in G1 and S phases of the second, third, and fourth cell cycle (Bolton et al., 1984; Sawicki et al., 1978; Smith and Johnson, 1986). To obtain G2 blastomeres, two-cell, four-cell, and eight-cell embryos were used immediately after being flushed out.
Isolation of blastomeres
Zonae pellucidae were removed from cleaving embryos by treatment with 0.5% pronase in PBS for 3–5 min. After rinsing the embryos in M2 medium they were transferred to Dulbecco's salt solution without Ca and Mg ions (Dulbecco and Vogt, 1954) for 15 min. After this treatment, the embryos were placed in M2 medium and were disaggregated into single blastomeres by repeated pipetting with a flame-polished narrow-bore pipette.
Micromanipulation tools and equipment
All manipulations were performed under inverted Leitz Fluowert microscope (Leitz, Germany) equipped with Nomarski differential interference contrast (DIC) and connected with Leitz (Germany) mechanical micromanipulators. Beaudouin Alcatel (France) and Cell Tram Air (Eppendorf, Germany) micropumps were connected with micromanipulation and holding pipettes, respectively. Pipettes were prepared from thin-walled borosilicate glass capillaries of an external diameter 1 mm (GC 100T-15, Harvard Apparatus Ltd, Kent, UK). Enucleation and nuclear injection pipettes were prepared using PB-7 vertical puller (Narishige, Japan). After pulling, the ends of pipettes were broken at the appropriate point using a MF-79 microforge (Narishige, Japan) and bevelled on an EG-4 grinding wheel (Narishige, Japan) to obtain a 45° cut. To facilitate penetration of the zona pellucida and to minimalize the damage of the oolemma, a spike was formed (by means of the MF-79 Narishige microforge) at the tip of enucleation and injection pipettes, and the tools were bent with a microforge (Alcatel, France).
Enucleation of oocytes
Enucleation of GV stage oocytes was performed as described by Grabarek et al. (2004). Prior to enucleation oocytes were incubated at 35°C in enucleation solution (M2 medium supplemented with 0.2 mM dbcAMP, 10 μg/mL of cytochalasin B (CB), and 0.25 μg/mL of nocodazole) for 30 min. Incubation of GV oocytes in enucleation solution resulted in deformations in their cell shape: one to five depressions in the cell surface were observed. The identations broadened the perivitelline space, which is otherwise too tight to place a pipette tip inside. Therefore, only the oocytes with indentations were used for experiments. After placing the oocytes in the micromanipulation chamber they were secured with the holding pipette and an enucleation pipette was gently introduced through the zona pellucida into the perivitelline space in the area of depression. The GV with a smallest possible amount of surrounding cytoplasm was removed by smooth suction.
Reconstruction of cytoplasts with karyoplasts or blastomeres
Blastomeres isolated from two-, four-, and eight-cell stages were preincubated in M2 medium supplemented with CB (5 μg/mL) for 20–30 min before being placed in a micromanipulation chamber filled with the enucleation medium. When blastomeres from the two- or four-cell stage were used, the blastomere was partially aspirated into the micropipette with its nucleus located inside the pipette. The reduction of the blastomere (cytoplasm) volume was achieved by “cutting off” the nonaspirated part of the blastomere during penetration of the pipette through the zona pellucida. In the case of blastomeres from the eight-cell stage, the entire cell was introduced under the zona pellucida. Donor karyoplasts or blastomeres were introduced into the perivitelline space of ooplasts through the slit in the zona pellucida made during enucleation. After insertion of a donor cell into the perivitelline space the pairs were washed out of enucleation solution, placed in M2 with dbcAMP to inhibit maturation, and incubated until electrofusion. Then, the complexes were washed twice in electrofusion solution (0.3 M mannitol supplemented with 0.1 mM CaCl2, and 0.1 mM MgSO4) and were placed between two parallel platinum electrodes in an electrofusion chamber filled with the same solution. Complexes were exposed to two DC pulses (50–60 μsec each, 1.8–2.5 kV · cm−1) generated by the BTX 2001 ElectroCell Manipulator (Genetronics, Inc., San Diego, CA, USA). Fusion usually occurred within 30 min, but if it failed, the application of electrical pulses was repeated. Fused pairs were then washed in M2 and were cultured in maturation medium for up to 17 h.
Heterokaryons
In a separate experiment intact GV oocytes were fused with blastomeres from the two-cell stage in the G2 phase of the cell cycle, and just after cell fusion heterokaryons were treated with dbcAMP for 3 h, to synchronize meiotic and mitotic nuclei in the G2 by preventing germinal vesicle breakdown. For cell fusion, the oocytes were devoid of zona pellucida with pronase, and oocyte/blastomere pairs were agglutinated with phytohemagglutinin (300 μg/mL PBS). Heterokaryons were cultured in M2 medium throughout maturation.
In vitro maturation of oocytes
Control nonmanipulated oocytes as well as enucleated oocytes reconstructed with the blastomere nuclei were matured in vitro in TCM199 medium supplemented with 10% (v/v) fetal calf serum (FCS), 75 (μg/mL penicillin G potassium salt, and 50 (μg/mL streptomycin sulfate.
Cytological studies during maturation
To examine the nuclear morphology at the time of expected MI (5 h postfusion) and after reaching MII (17 h postfusion), the oocytes were fixed in 4% paraformaldehyde and stained for DNA with 10 μg/mL Hoechst (33342). Nuclear morphology of the MI and MII stages as well as presence of the first polar body (1st PB) were analyzed under an epifluorescence microscope. Whole-mount preparations were made of some reconstructed oocytes (Tarkowski and Wróblewska, 1967) and were viewed and photographed under bright field.
Metaphase I and II were scored normal, when chromosomes were visible at the equatorial plate of the spindle. Abnormal nuclear progression was when either the donor nucleus stayed intact or the chromosomes were scattered throughout the spindle.
Results
The 1st PB extrusion
Once the 1st PB is extruded and metaphase II is reached, mammalian oocytes are blocked at this stage until fertilization. Therefore, one can fix metaphase oocytes at 17 h postfusion, although metaphase II is reached few hours earlier (about 12 h postfusion). When the reconstructed oocytes were cultured for 17 h and examined for the 1st PB extrusion, striking efficiency of the S phase donors in stimulating cell division was observed. About three-quarters of the oocytes reconstructed with the S phase karyoplasts or blastomeres underwent their first meiotic division in vitro (Fig. 1). From 55.9 to 62.5% of the cytoplasts reconstituted with the G2 blastomeres divided. After synchronization of the G2 meiotic and mitotic nuclei in heterokaryons (Supplementary Fig. 3), the 1st PB extrusion was 10% less frequent than in the respective cybrids (the G2 donors from two-cell stage) (Fig. 1). The G1 phase blastomeres from the two- and four-cell stages supported cytokinesis only in 22.2 and 25.6% of oocytes, respectively. The G1 donors from eight-cell stage induced cell division almost twice as effectively (43.3%) as donors from four-cell and two-cell embryos.
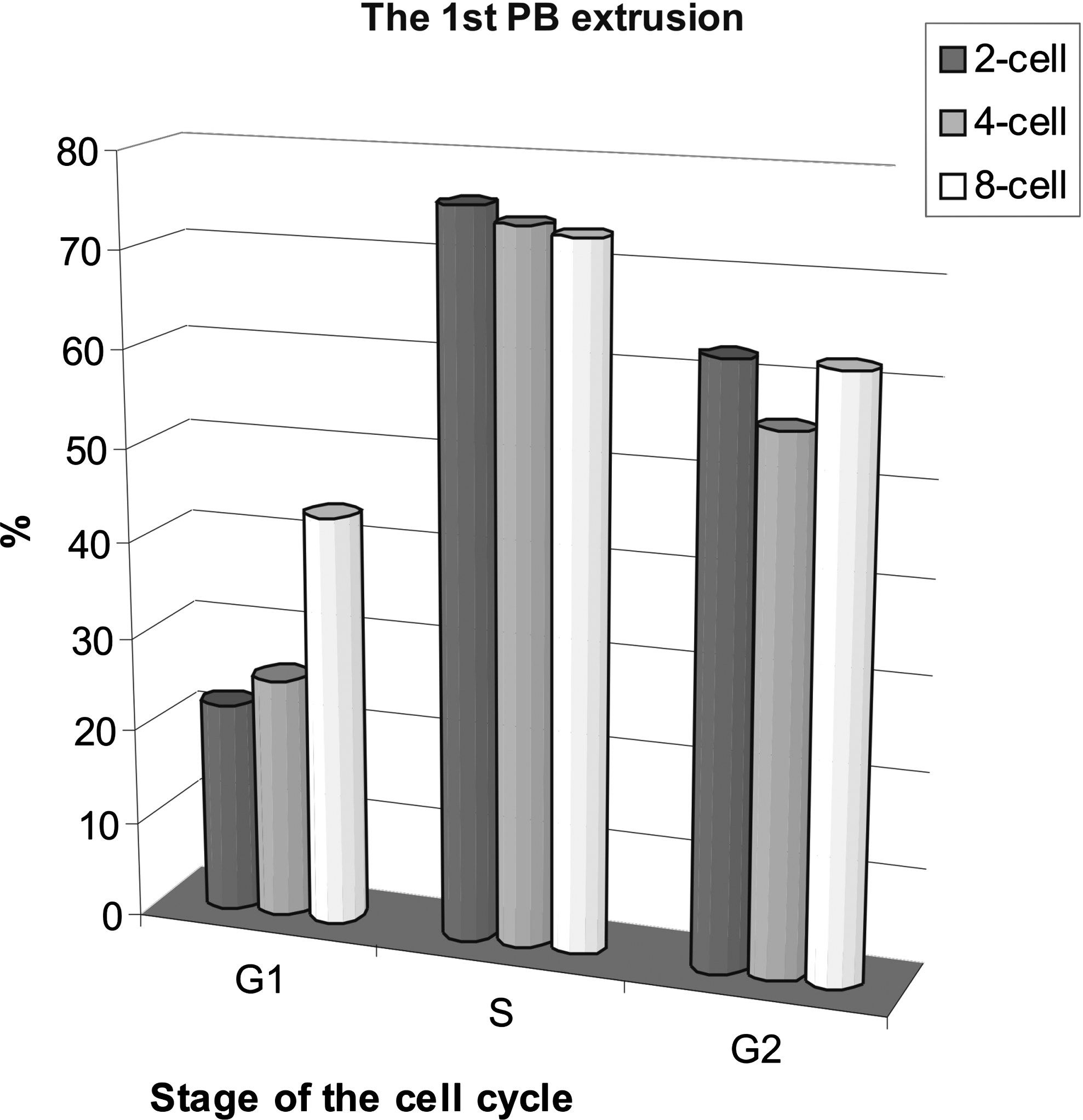
Effects of the cell cycle stage on the 1st PB extrusion in enucleated GV oocytes reconstructed with blastomere nuclei from two-, four-, or eight-cell stage. Each column shows the results from 27–45 oocytes. The differences between G1, S, and G2 stages are statistically significant (paired t-test; G1–S: p = 0.025, S–G2: p = 0.022, G1–G2: p = 0.04). Control oocytes (n = 30) extruded the 1st PB in 83.3%.
PB size
Abnormal, large PBs were extruded least frequently by cytoplasts reconstituted with the G1 blastomeres (Fig. 2). The G1 donors from the two-cell stage best stabilized asymmetric division; there was a tendency to slightly loosen the control with later stages (four cell, eight cell). However, it cannot be attributed to decreasing cytoplasmic volume, because only small karyoplasts were taken from the two-cell stage blastomeres to be fused with ooplasts, comparable to four-cell stage and eight-cell stage blastomeres.
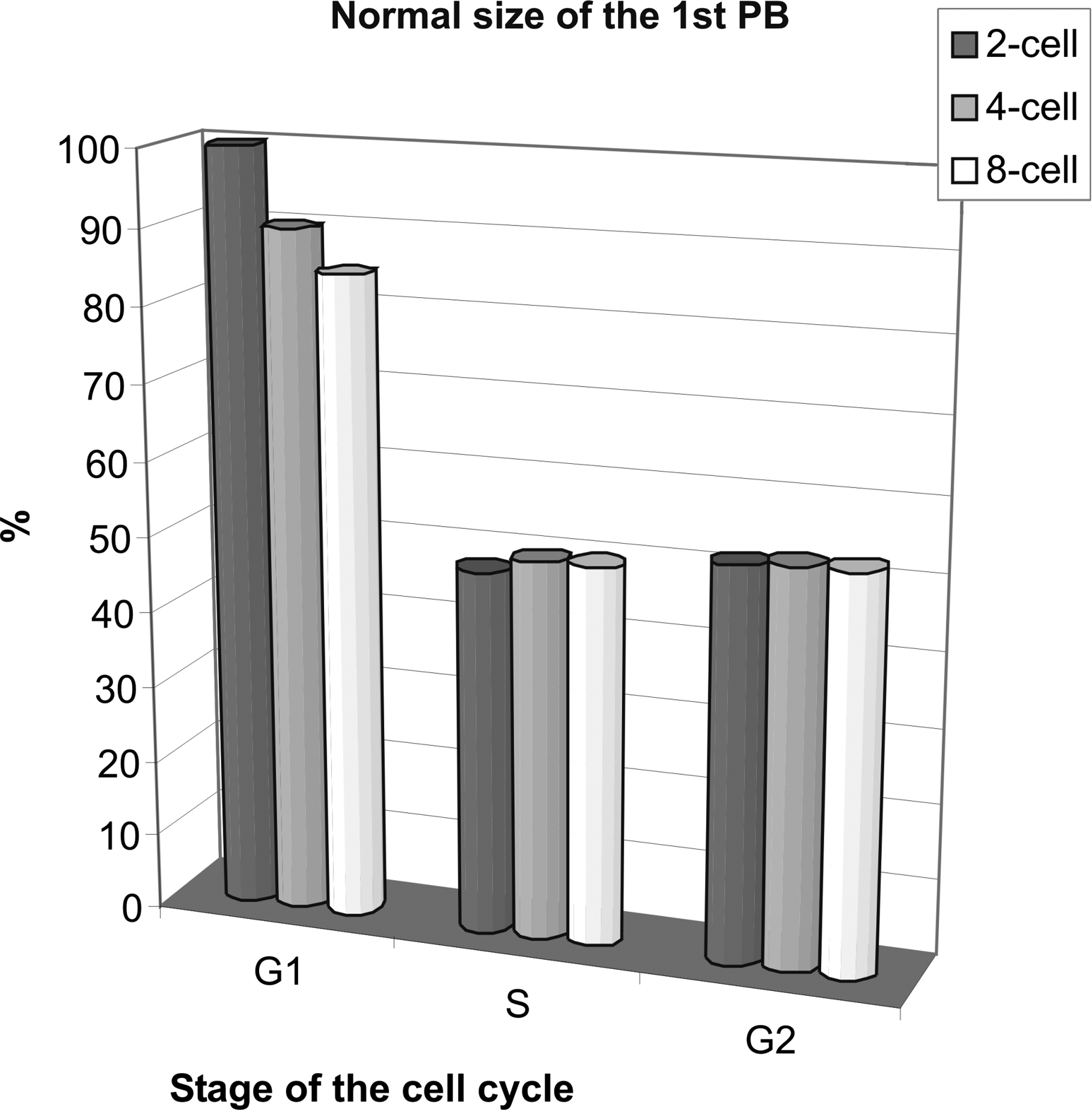
Effects of the cell cycle stage on the size of the 1st PB in enucleated GV oocytes reconstructed with blastomere nuclei from two-, four-, or eight-cell stage. Each column shows the results from 27–45 oocytes. The differences between G1, S, and G2 stages are statistically significant (paired t-test; G1–S: p = 0.014, S–G2: p = 0.022, G1–G2: p = 0.014).Control oocytes (n = 30) had normal (small) 1st PBs in 100%.
Blastomeres in the S phase and G2 phase induced large PB formation in about 50% of reconstituted cytoplasts. The frequency of normal 1st PB was almost twice as high after synchronization of the G2 meiotic and mitotic nuclei in heterokaryons (Supplementary Fig. 3), as in the respective cybrids (cytoplasts reconstructed with two-cell donors in the G2) (Fig. 2). Asymmetric division in the oocytes reconstructed with the S and G2 donors was independent of their developmental stage.
Metaphase I (MI)
When examined at the expected time of metaphase I (5 h postfusion), about 30% of oocytes carrying the G1 donors had normal metaphase I independent from the developmental stage of the donor (Supplementary Fig. 4). Normal metaphase I was induced twice as often when using the G2 donors: from two-cell stage, 64%; four-cell stage, 55%; and eight-cell stage, 71.4% (Fig. 3). The frequency was comparable with, but slightly higher than their 1st PB extrusion (Fig. 1). The nuclei in the S phase drove normal MI in up to 27% of oocytes. This is remarkably low, given their efficiency of 1st PB extrusion, which was about 75% (Fig. 1).
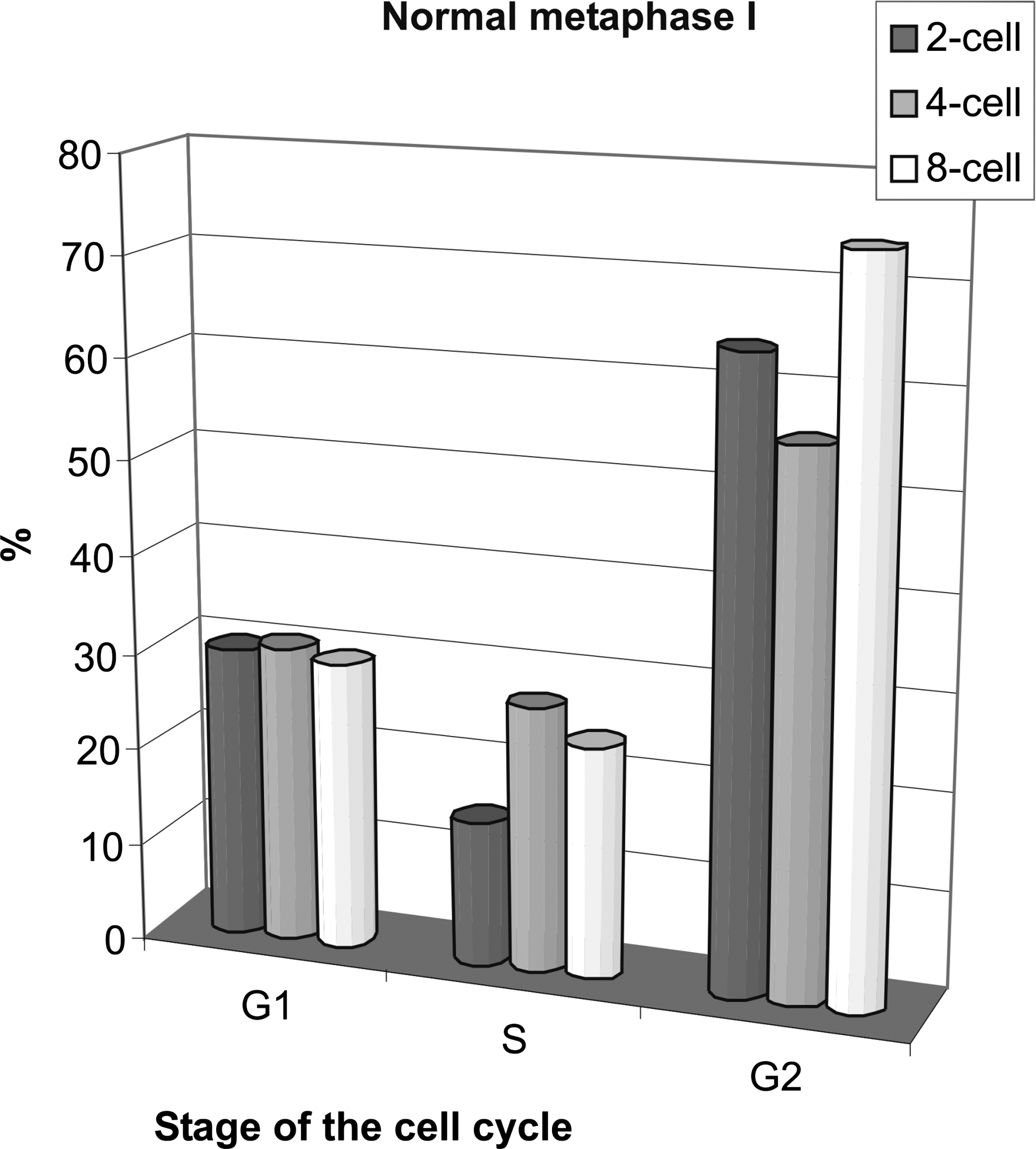
Effects of the cell cycle stage on the frequency of normal metaphase I in enucleated GV oocytes reconstructed with blastomere nuclei from two-, four-, or eight-cell stage 5 h after fusion. Each column shows the results from 20–27 oocytes. The differences between G1 and G2 as well as between S and G2 stages are statistically significant (paired t-test; S–G2: p = 0.027, G1–G2: p = 0.026). Control oocytes (n = 30) had normal MI in 100%.
Very high numbers of reconstructed oocytes showed an abnormal pattern of nuclear progression. The presence of interphase-like nuclei was an indication of inefficient progression of donor nucleus to metaphase. The frequency of interphase nuclei was 40% of oocytes reconstructed with the G1 donors from two-cell stage, and 35%, with the S donors (Supplementary Fig. 1; see online supplementary material at www.liebertonline.com). Among both the G1 and S donors, there was a tendency that interphase nuclei were the less frequent, the later the developmental stage of the donor. The G2 donors induced interphase nuclei only when they originated from four-cell stage blastomeres. The other abnormal nuclear pattern comprised metaphase plates with chromosomes scattered throughout, which represented improper meiotic progression. The frequency of scattered chromosomes was proportional to the developmental stage of the G1 donors; among the S and G2 donors this proportionality was disturbed by an overrepresentation of scattered chromosomes when two-cell embryos were used. The S donors induced scattering of chromosomes in up to 52% of oocytes.
Normal metaphase II
The nuclear configurations of the reconstructed oocytes were examined a few hours after the expected time of MII stage (about 12 h postfusion), after an overnight culture, at 17 h postfusion. Properly condensed chromosomes were present in all control oocytes and no abnormal configurations were found (n = 30) (Supplementary Fig. 5).
Cytoplasts fused to blastomeres from eight-cell stage in the G2 phase of their cell cycle exhibited the highest frequency of normal metaphase II (68%) (Fig. 4A) among all reconstructed oocytes. Normal metaphase II was found in 52% of oocytes reconstructed with the two-cell- and four-cell blastomere nuclei in the G2 (Fig. 4A). The frequency of normal metaphase II was 10% higher in the G2-synchronized heterokaryons (Supplementary Fig. 3), than in the respective cybrids (cytoplasts reconstructed with two-cell donors in the G2) (Fig. 4A). Unexpectedly, normal MII chromosomes were also found in the G2-reconstructed oocytes without 1st PB (Fig. 4B). They were as frequent among nonextruding oocytes as among the extruding oocytes, decreasing only in those receiving the four-cell stage blastomere nuclei.
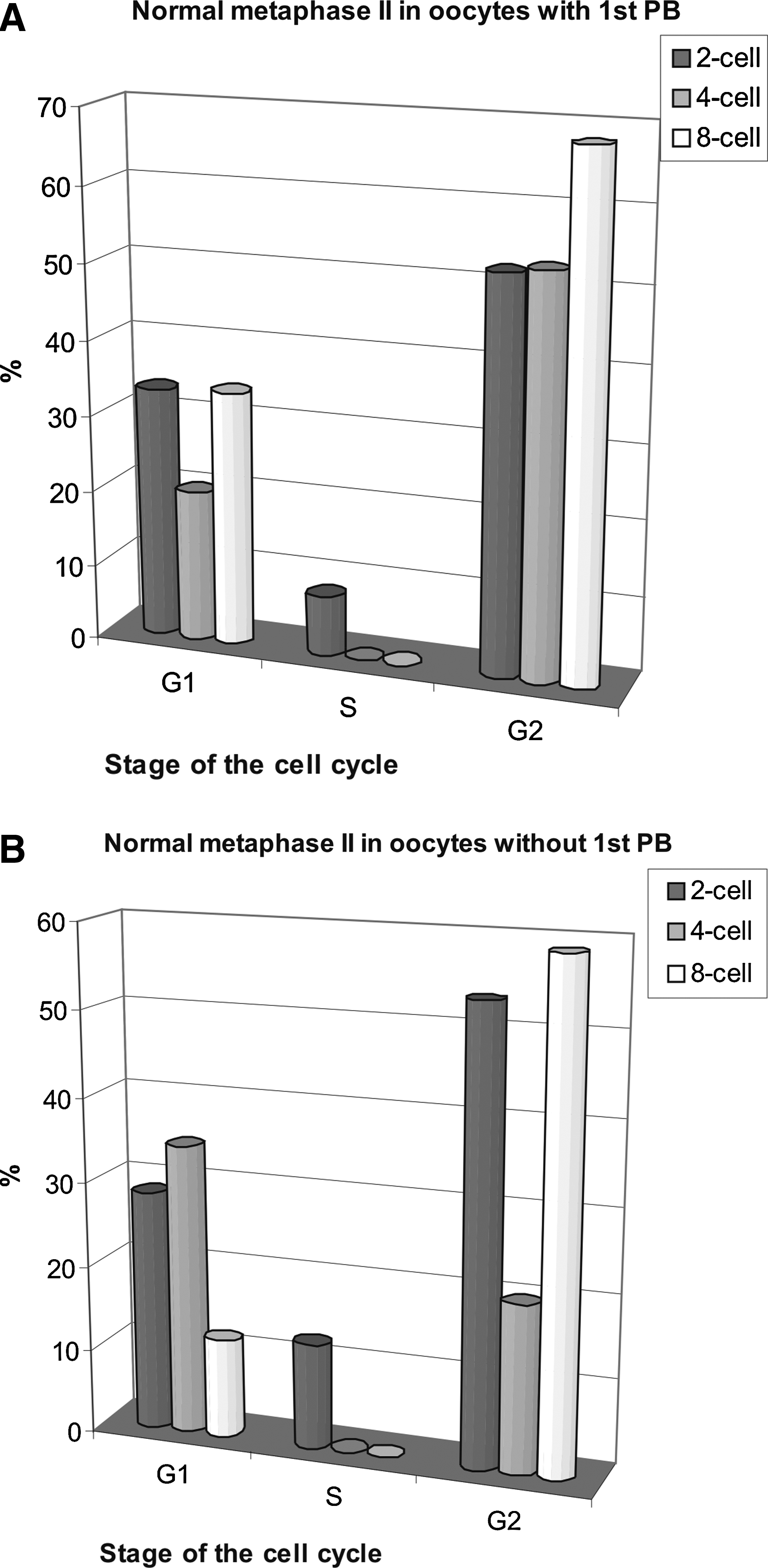
Effects of the cell cycle stage on the frequency of normal metaphase II in enucleated GV oocytes reconstructed with blastomere nuclei from two-, four-, or eight-cell stage; 17 h after fusion. (
When nuclei from two-cell and eight-cell blastomeres in the G1 stage were used, normal metaphase II was found in 33% of oocytes extruding 1st PB. However, the G1 nuclei from four-cell stage supported MII only in 20% of reconstructed oocytes. The frequency of normal MII among nonextruding oocytes carrying nuclei from two-cell stage was similar to that among the extruding oocytes. When four-cell stage blastomere nuclei in the G1 were the donors, normal MII was found in 34% of nonextruding oocytes. On the other hand, with eight-cell stage blastomere nuclei in the G1, normal metaphase II was only found in 12% of nonextruding recipients.
Normal MII chromosomes were present in about 10% of GV-cytoplasts reconstituted with the S phase two-cell blastomere nuclei and in no cytoplasts reconstructed with the S phase four- and eight-cell nuclei (Fig. 4A and B).
Abnormal metaphase II
In reconstructed oocytes two types of anomalies were found, similar to those present at the time of metaphase I. Interphase nuclei were only present in oocytes without the 1st PB (with one exception). In nonextruding oocytes interphase nuclei could be found in up to 38% of oocytes (Supplementary Fig. 2B). The frequency of interphase nuclei after the G1 stage reconstruction was inversely proportional to the developmental stage of the nuclear donor.
Scattered chromosomes was the second anomaly found. Cytoplasts fused to blastomeres at the eight-cell stage in the G2 phase of their cell cycle exhibited the fewest anomalies of any group of oocytes extruding 1st PB: 32% of scattered chromosomes (Supplementary Fig. 2A). Oocytes extruding 1st PB after reconstruction with the G1 nuclei revealed scattered chromosomes in 66.6% of those receiving two-cell stage blastomere nuclei, but as much as 80% of those carrying four-cell stage nuclei (Supplementary Fig. 2A). Together with intact nuclei the anomalies amounted 88% of the G1 recipients (nonextruding the 1st PB and carrying eight-cell stage blastomere donors) (Supplementary Fig. 2B).
All oocytes reconstructed with the S nuclei from four-cell and eight-cell stage blastomeres revealed scattering of chromosomes, independent of extruding the 1st PB (Supplementary Fig. 2A and B). Almost 90% of those reconstructed with two-cell stage nuclei in the S phase were abnormal, comprising 37.5% of interphase nuclei and 50% of scattered chromosomes.
Timing of maturation
Germinal vesicle breakdown (GVBD) occurred 2 h after the onset of the culture in about 60% of the control oocytes (n = 30), whereas NEBD began in the third hour in the reconstructed cytoplasts. Control GV oocytes placed in maturation medium immediately after being collected from the ovaries extruded the first PB 9–12 h after the onset of maturation (Fig. 5, control). All GV-cytoplasts reconstructed with the S nuclei extruded their PBs 2 h earlier than the control, that is, between 7 h and 9 h postfusion (Fig. 5A–C). After reconstruction with the G1 nuclei, the first meiotic division was even more accelerated and commenced 7–8 h postfusion. Notably, all oocytes in this group divided within 1 h, either between 6 and 7 h, like those receiving nuclei from four-cell stage blastomeres (Fig. 5B), or between 7 and 8 h, like those carrying two-cell stage nuclei (Fig. 5A) and eight-cell stage nuclei (Fig. 5C). There was a tendency that among the recipients of the G2 nuclei the extent of acceleration was proportional to the developmental stage of the donor blastomere: those carrying two-cell blastomere nuclei divided between 6 and 8 h, four-cell nuclei between 7 and 9 h, and eight-cell nuclei between 7 and 10 h.
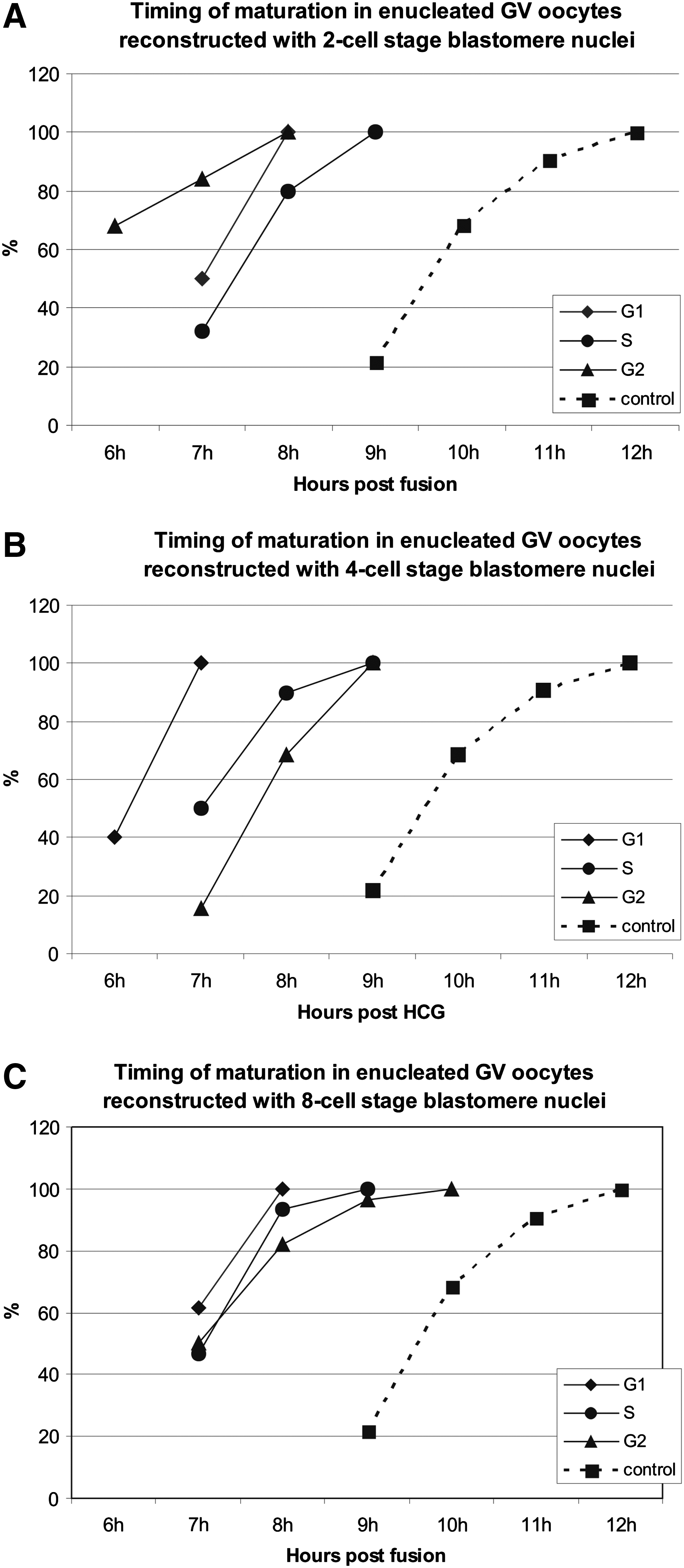
Timing of the 1st PB extrusion in reconstructed oocytes. Control = intact oocytes (n = 32). (
Discussion
The effects imposed by embryonic donor nuclei on oocyte maturation
The cell cycle stage of the donor nucleus strongly affects maturation after nuclear transfer of embryonic nuclei into enucleated GV oocytes, contrary to the earlier suggestions (Polanski et al., 2005). Nuclear donors in the S phase of the cell cycle induce almost 75% of reconstructed oocytes to divide, despite their omnipresent chromosomal anomalies in MII. The G1 donors bring about strict control of the 1st PB size and repeatedly support normal MI and MII, as well as 1st PB extrusion, but only in about one-third of the reconstructed oocytes. The G2 donors allow for the normal course of maturation (1st PB extrusion, size of 1st PB, normal MI, and MII) in about half to two-thirds of reconstructed oocytes. Donor–recipient incompatibility of the cell cycle stage could well explain very high metaphase II failure under the S donors and a moderate efficiency of MII under the G1 donors. Synchronization of the meiotic and mitotic G2 stage in heterokaryons slightly improved normal MII compared to cybrids.
Among the recipients of the G1 and G2 nuclei, those extruding the 1st PB often show similar frequencies of normal MII as the nonextruding oocytes. This suggests loosening of normal correlation between the meiotic nuclear progression and cytokinesis. Neither the extrusion of the 1st PB by oocytes reconstructed with the S phase donors is evidence of chromosomal maturation, nor the absence of the 1st PB in oocytes receiving the G1 and G2 nuclei indicates improper chromosomal maturation.
There are some effects imposed on the oocyte maturation by the developmental stage of the donor. Recipients receiving the G1 donors extrude 1st PB twice as often when harboring the eight-cell blastomere nuclei, as with two-cell and four-cell nuclei. The G2 blastomeres from four-cell stage confer decreased frequency of 1st PB extrusion, as well as of normal MI and MII in oocytes extruding the 1st PB, in comparison with the two- and eight-cell stage. The G2 blastomeres from the eight-cell stage yield higher percentages of normal MI and MII than those from two-cell and four-cell stages, independent of the 1st PB extrusion. Conferring this chromosomal normalcy renders the G2 blastomeres from eight-cell stage the best donors for the GV oocyte reconstruction of all tested in our experiments. Neither the above exceptions, nor the general tendency to equally support maturation of enucleated GV oocytes by the nuclei of blastomeres from two-, four-, and eight-cell stages, are in keeping with the idea of maternal proteins from those nuclei affecting maturation (as suggested by Polanski et al., 2005) due to gradual change from the meiotic to the mitotic program in the course of the first mitotic cell cycles of the dividing embryo (Ciemerych et al., 1999).
Meiotic ability of mitotic chromosomes throughout donor cell cycle
It should be borne in mind when comparing zygotes with GV oocytes as nuclear recipients that the former are in the mitotic cell cycle, whereas the latter have yet to undergo two meiotic divisions, the first of which is a reduction that begins by pairing homologous chromosomes and leads to their separation between the oocytes and the 1st PBs. To ensure balanced chromosome segregation during meiosis, each homologous pair at MI and sister chromatid pair at MII must be aligned on the metaphase plate of the spindle (Sluder and McCollum, 2000).
Interaction of the meiotic spindle with mitotic chromosomes has recently been analyzed in enucleated mouse GV oocytes fused to metaphase somatic cells (fibroblasts) (Chang et al., 2006). Complete and accurate alignment of chromosomes was not established in any reconstructed oocyte. However, bipolar spindles were built up and the 1st PBs were extruded. In mouse Sycp3−/−oocytes univalent chromosomes are present, which preserve meiotic kinetochore structure, but which are also bioriented in a mitotic manner (Kouznetsova et al., 2007). In such oocytes the time between germinal vesicle breakdown and PB extrusion is similar to wild-type oocytes. These and our results clearly show that neither misaligned chromosomes in reconstructed oocytes (Chang et al., 2006; this paper, the S phase donors) nor the presence of univalents in oocytes (Kouznetsova et al., 2007) induced the spindle checkpoint-related metaphase arrest. The meiotic spindle assembly checkpoint (SAC) depends on Mad2 (Homer et al., 2005; Tsurumi et al., 2004). Reducing Mad2 levels leads to an acceleration of meiosis I, not leaving enough time for proper attachment of the bipolar spindle to functional kinetochores in metaphase I. SAC function is affected, and therefore, oocytes with unattached kinetochores will eventually progress into meiosis II. Also, Mad2 overexpression interferes with chromosome segregation in meiosis I (Niault et al., 2007).
Our data show that the S phase of the blastomere cell cycle is not compatible with successful meiosis of reconstructed oocytes. Incomplete DNA synthesis and pulverization of condensing chromatin can be the reason of it. The possibility that the S phase nuclei could complete replication before NEBD can be ruled out, as DNA synthesis in embryonic nuclei does not stop the meiotic progression (GV breakdown) (Fulka et al., 1995). Both G1 and S phase donor nuclei are prone to DNA damage during chromosome condensation of 2C and hybrid 2C/4C DNA, respectively. Although DNA damage invariably arrests cell cycle progression of somatic cells in the late G2 phase (Maltzmann and Czyzyk, 1994)
Meiosis requires maintaining bivalents during condensation, setting up mechanisms to prevent sister-chromatid separation during first meiosis (these include suppressing biorientation of sister kinetochores) (Petronczki et al., 2003), and controlling the biphasic destruction of cohesins (Kitajima et al., 2004). It is likely that these mechanisms could more properly operate, when the G2 ooplasts are reconstructed with the G2 nuclei than with the G1 or S nuclei.
Timing of the first PB extrusion
The acceleration of the first PB extrusion has been observed in all experiments involving GV cytoplasts reconstructed with G1 and G2 donor nuclei (Bui et al., 2008; Chang et al., 2004; Grabarek et al., 2004; Mohammed et al., 2008; Nan et al., 2007; Polanski et al., 2005). In our experiments, abnormal acceleration of maturation could not be overcome by donors at any cell cycle stage (the S phase included) or at any developmental stage (blastomeres from two-cell, four-cell, or eight-cell embryos). Heterokaryons are slightly less accelerated than the respective cybrids (data not shown), indicating the role of the nucleus (GV) in the timing of maturation. However, when GV oocytes are selectively enucleated so that the liquid contents of GV remains in the cytoplasm, their maturation after reconstruction with fibroblasts' nuclei is even more accelerated than after complete enucleation (Mohammed et al., 2008), suggesting that the liquid contents of GV is not instrumental.
In all cell types where the SAC functions, its disruption results in the acceleration of anaphase/division, including mouse oocytes (e.g., Homer et al., 2005; Tsurumi et al., 2004) in which under such conditions 1st PB is formed 2–4 h earlier. The possibility that premature 1st PB formation in reconstructed GV oocytes also results from disruption of SAC has been indicated (Polanski et al., 2005). It seems that the analysis of the chromosome configuration at MI in reconstructed oocytes performed in the present study further support such hypothesis. Reconstructed oocytes frequently exhibit abnormal chromosome configuration at 5 h after fusion, yet 1–2 h later the majority of them has undergone division. This strongly suggests that in many reconstructed oocytes the SAC did not prevent anaphase and division despite chromosome anomalies, resulting in the acceleration of 1st PB formation. Finally, it was shown that nuclear transfer into MII bovine oocytes resulted in SAC disruption (Tani et al., 2007). This may be the effect of maternal/embryonic transition in transcription, resulting in paternally derived protein products being detectable at the late two-cell stage (for review, see Telford et al., 1990). That the G2 blastomeres from the two-cell stage (late two-cell stage) and the G1 blastomeres from the four-cell stage are the most accelerated in extruding 1st PB seems to confirm such hypothesis.
GV oocyte as a recipient of embryonic nuclei
Many reports (Bui et al., 2008; Chang et al., 2004, 2006; Garbarek et al., 2004; Mohammed et al., 2008; Nan et al., 2007; Polanski et al., 2005) have demonstrated that enucleated GV oocytes reconstructed with embryonic or somatic nuclei can mature, although under constraint of accelerated extrusion of 1st PB, abnormally large 1st PB, and abnormal MI and MII chromosomes. Our study confirms these observations and extends them to show how the donor cell cycle stage affects the anomalies.
A GV oocyte, harboring an interphase nucleus in the G2 phase of the cell cycle, could be considered an ideal universal recipient for nuclear transfer. However, chromosomal anomalies in maturation, induced by the embryonic S donors, exclude these donors. High frequency of normal 1st PB extrusion by the recipients of the G1 donors, together with up to 40% of normal MII in this group makes it an interesting possibility. Inhibition of the 1st PB could be applied to ooplasts reconstructed with the G1 blastomere nuclei, to prevent random segregation of their single chromatid chromosomes at MI. Also, about one-third of nonextruding oocytes reconstructed with the G1 blastomeres from two- and four-cell stage reaches normal MII (cf. Fig. 4B), supplying already “inhibited” oocytes.
The GV-cytoplasts achieved their greatest maturation efficiency and were least disturbed by abnormal meiosis when the G2 blastomeres from the eight-cell stage were used. The efficiency of proper GV-cytoplast reconstruction with such blastomeres was about 25% (maturation 62.5% × normal-sized 1st PB 52.6% × normal MII 67.9%). The respective efficiency with fibroblast nuclei was 19% (maturation 52.1% × normal-sized PB 66.1% × normal MII 55.2%) (as calculated from Chang et al., 2004). Also, above half of the nonextruding oocytes reconstructed with the G2 blastomeres from two- and eight-cell stages reaches normal MII (cf. Fig. 4B), supplying oocytes with a full complement of two-chromatid chromosomes, ready to be activated for development.
Comparison of reconstructed GV oocytes (this article) with the reconstructed zygotes (Smith et al., 1988) indicates that the former are not universal recipients, because they do not accept the S phase donors. On the other hand, when the G2 donors are used, then meiotic maturation of reconstructed oocytes, as measured with the frequency of normal MII, is equal to the frequency of blastocyst formation by reconstructed zygotes (Smith et al., 1988). The developmental potential of GV ooplasts reconstructed with the G2 embryonic nuclei is also worth studying because they can support preimplantation development, at least after selective enucleation followed by maturation, transfer of the G2 blastomere nucleus from the two-cell stage, artificial activation and inhibition of the 2nd PB extrusion (Mohammed et al., 2008).
The GV ooplasts harboring the G1 donors (this article) are twice less efficient in chromosomal maturation than are mitotic recipients (zygotes) (Smith et al., 1988) in driving blastocysts formation. However, enucleated GV oocytes are still worth considering as recipients of the G1 nuclei, because about 50% of reconstructed oocytes have been effective in remodeling donor G1 (somatic) nuclei toward meiotic nuclear morphology and normal MII (Bui et al., 2008). In donor nuclei, histone H3 demethylation, and its phosphorylation indicative of chromosome condensation were demonstrated (Bui et al., 2008).
Footnotes
Acknowledgments
The authors are grateful to Mrs Maria Górniewska for excellent technical assistance.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.