
Abstract
Abstract
In this work, Dehydroleucodine (DhL) was evaluated as a chemical activator of bovine oocytes and somatic cell nuclear transfer (SCNT) reconstituted embryos. Oocytes were activated with 5 μM Ionomycin (Io) and exposed for 3 h to 1 or 5 μM DhL alone (Io-Dhl1 or Io-DhL5) or combined with Cytochalasin B (Io-DhL1/CB; Io-DhL5/CB). Control groups were Io (Io), Io followed by 1.9 mM 6-Dimethylaminopurine (Io-6DMAP), and embryos produced by in vitro fertilization (IVF). Pronuclear formation and development to blastocysts of activated oocytes were evaluated. Embryos obtained by the DhL concentration that induced the highest blastocyst rates (1 μM) were karyotyped. An additional treatment based in Io-DhL1 plus lengthened (6-h) exposure to CB (Io-DhL1/long CB) was included to improve the proportion of diploid blastomeres. Finally, DhL combined with CB was employed to assist cloning by intracytoplasmic injection of whole cumulus cells. Results showed that DhL induces a pronuclear formation dynamic that was more similar to IVF-produced embryos than DMAP. Development to blastocyst stage was higher after activation with 1 μM DhL than with 5 μM DhL, either for groups combined or not with CB (19.15; 21.74 vs. 6.82; 0%, respectively) (p < 0.05). Io-DhL1 and Io-DhL1/CB treatments induced blastocyst-cleaved embryo ratios not statistically different from those of Io-DMAP (35.85%) and IVF (33.33%) groups (p > 0.05). Io-DhL1/long CB induced higher diploid blastomere rates than Io-Dhl1, Io-DhL1/CB and Io-DMAP (63.8 vs. 36.8; 40 and 31.6%, respectively) (p < 0.05). Moreover, all DhL treatments resulted in polyploidy rates that were lower than Io-DMAP (5.2, 12.0, 10.6, and 31.6%, respectively) (p < 0.05). Io-DhL1/CB and Io-DhL1/long CB induced cloned embryo blastocyst rates that were not significantly different from Io-DMAP (6.1, 9.4, and 18.3%, respectively) (p < 0.05). Our results indicate that Io-DhL1/long CB protocol could be useful for SCNT programs.
Introduction
Although cloning provides a powerful tool for biotechnological applications, production rates of viable offspring still remain low (Kato et al., 1998; Wilmut et al., 1997; reviewed by Kishigami et al., 2008), the activation of recipient oocytes being one of the factors involved in the success of somatic cell nuclear transfer (SCNT) protocols (Ross et al., 2009; Whitworth et al., 2009).
Oocyte activation induces exit from metaphase II arrest and triggers embryonic development. This arrest is sustained by high levels of maturation promoting factor (MPF) (Nurse, 1990), a heterodimer composed of cyclin B and p34cdc2 kinase (Arion et al., 1988; Draetta et al., 1989; Labbé et al., 1989; Murray et al., 1989). MPF activity oscillates during the cell cycle depending on the balance between permanent synthesis and degradation of cyclin B (Kubiak et al., 1993) and on its phosphorylation status (Gautier et al., 1989). MPF activity peaks at metaphase (Lohka, 1989), due to stabilization by the c-mos active component of the “cytostatic factor” (CSF), which inhibits cyclin B degradation (Kubiak et al., 1993; Masui, 1991; Masui and Markert, 1971; Nganvongpanit et al., 2006; Wu et al., 1997).
After normal fertilization, calcium oscillations induced by sperm penetration release the oocyte from metaphase II (MII) arrest by degrading cyclin B and CSF (Watanabe et al., 1991). MPF complex is inactivated by phosphorylation of cdc2 on Thr161 and Thr14/Tyr15. These inhibitory phosphorylations are probably catalysed by Myt1 protein kinase (Gavin et al., 1999; Inoue et al., 2005; Patra et al.,1999; Qian et al., 2001; Wells et al., 1999b). Calcium ionophores, like ionomycin (Io), can be employed to initiate artificial activation of mammalian oocytes through the induction of an intracellular calcium peak and the consequent inactivation of MPF (Liu et al., 1998a). To complete oocyte activation, MPF must remain inactive. Several other agents, such as the protein kinase inhibitor 6-dimethylaminopurine (DMAP), are used after Io exposure (Szöllösi et al., 1993). These agents induce high rates of pronuclear formation and blastocyst development in bovine oocytes (Liu et al., 1998b; Susko-Parrish et al., 1994). Several authors have reported offspring production in this species by activation of SCNT- derived embryos with Io-DMAP (Cibelli et al., 1998; Wells et al., 1999a). Most pregnancies failure (Cibelli et al., 1998) could, in part, be attributed to the high incidence of polyploidies observed in cloned embryos exposed to 6-DMAP (Alexander et al., 2006; Bhak et al., 2006).
The aim of this work was to employ the new compound, Dehydroleucodine (DhL), to induce chemical activation of bovine oocytes. DhL is a sesquiterpenic lactone reported to selectively induce a transient arrest of the cell cycle in G2 stage of vegetal (Lopez et al., 2002) and animal (Cruzado et al., 2005) somatic cells. As well, DhL has been shown to inhibit maturation of Bufo arenarum oocytes (Sánchez Toranzo et al., 2007). A recent work has proposed a protein kinase, Myt-1 kinase, as a possible target of DhL responsible for MPF inactivation (Sánchez Toranzo et al., 2009). In the present study, DhL was used alone or in combination with Cytochalasin B (CB) to induce chemical activation of bovine oocytes previously treated with Io. Pronuclear (PN) dynamics, blastocyst development, and ploidy of parthenogenetic embryos were evaluated and, for the first time, DhL was combined with CB to induce in vitro development of SCNT embryos produced by intracytoplasmic cell injection (ICI).
Materials and Methods
Unless otherwise indicated, all chemicals were purchased from Sigma Chemical Company (St. Louis, MO, USA).
Cumulus–oocyte complexes (COCs) collection and in vitro maturation (IVM)
Cow ovaries were transported from a local slaughterhouse to the laboratory in a thermo container at 24 to 27°C. COCs were aspirated from follicles with a diameter of 2–8 mm into Dulbecco's phosphate-buffered saline (DPBS; 14287–072; Gibco BRL, Grand Island, NY, USA) containing 10% v/v fetal bovine serum (FBS; 10499–044; Gibco BRL) and 1% v/v antibiotic–antimycotic (ATB; 15240–096; Gibco BRL). Oocytes covered with at least three layers of granulosa cells were selected for IVM. The maturation medium was bicarbonate-buffered TCM-199 (31100–035; Gibco BRL), containing 2 mM glutamine (G-8540), 10% v/v FBS, 2 μg mL−1 follicle-stimulating hormone (NIH-FSH-P1; Folltropin; Bioniche, Belleville, Ontario, Canada), 0.3 mM sodium pyruvate (P2256), 100 μM cysteamine (M9768), and 1% v/v ATB. Groups of 25–30 COCs were in vitro matured in 100 μL droplets of maturation medium covered with mineral oil (M8410), at 39°C in a humidified atmosphere of 6% CO2 in air. After 21–24 h of IVM, cumulus cells were removed from COCs by vortexing for 2–3 min in hyaluronidase solution (H-4272; 1 mg mL−1 HEPES-TALP) and washed three times in HEPES-TALP. Oocytes with an extruded first polar body (PB) were selected and immediately activated or used for enucleation and SCNT.
Chemical activation
After IVM, selected oocytes were randomly assigned to the following activation treatments: 5 μM ionomycin (I24222; Invitrogen, Carlsbad, CA, USA) in HEPES-TALP for 4 min, either alone (Io) or followed by incubation for 3 h in either 1 μM (Io-DhL1) or 5 μM (Io-DhL5) DhL (Instituto de Embriologia e Histologia, Mendoza, Argentina) alone or combined with 5 μg mL−1 CB (C6762) (Io-DhL1/CB and Io-DhL5/CB, respectively) and 1.9 mM 6-DMAP, (D2629) for 3 h (Io-DMAP). CB was added to the activation medium to inhibit second PB extrusion. DhL and 6-DMAP were diluted in TCM-199 medium. After single or combined treatments, oocytes were thoroughly washed in HEPES-TALP and cultured as described below.
In vitro fertilization (IVF)
The IVF procedure was previously described by Brackett and Oliphant (1975). Briefly, frozen semen was thawed in a 37°C water bath for 30 sec. The sperm were washed twice by centrifugation at 400 × g with Brackett's defined medium. Sperm concentration was adjusted to 15 × 106 mL−1 and sperm then coincubated for 5 h with COCs in Brackett's fertilization medium. Afterward, the oocytes were washed several times in HEPES-TALP and in vitro cultured as described below.
In virto culture and determination of blastocyst cell number
Activated/fertilized/reconstituted embryos were in vitro cultured in 50 μL droplets of synthetic oviductal fluid (SOF) (Tervit et al., 1972, modified by Holm et al., 1999) containing 2.5% v/v FBS under mineral oil, at 39°C in a humidified atmosphere of 6% CO2 in air. Cleavege, morula and blastocyst stages were evaluated at days 2, 5, and 8 of in vitro development, respectively. Percentages of morulae and blastocysts were calculated over cleaved embryos. Blastocyst cell numbers were determined at day 8 of in vitro development, by staining with 1 μg mL−1 of Hoechst Bisbenzimide 33342 (H33342: B-2261) for 10 min. Nuclei were visualized and counted using UV light under a fluorescence microscope.
Assessment of nuclear stage
Zero, 4, 11, and 17 h postactivation (h.p.a.), parthenogenetic and in vitro fertilized oocytes were stained with 1 μg mL−1 of H33342 for 10 min. IVF oocytes were denuded after sperm oocyte coincubation. For this group, evaluation hours were corrected for sperm penetration time (4 h after the beginning of sperm–oocyte coincubation). Nuclear stage was evaluated using a UV microscope. Extrusion of the second PB was evaluated at 11 and 17 h.p.a.
Chromosomal analysis
Forty-eight hours after activation, parthenogenetic embryos were cultured for 6 h in SOF medium containing 1 μg mL−1 colchicine (C3915) then transferred to a trisodium citrate hypotonic solution (F71497; 0.8% w/v in distilled water) for 15 min at 37°C. Subsequently, embryos were placed on a clean glass slide in a small volume of medium and a methanol:acetic acid solution (3:1, v:v) was applied. After air drying, fixed embryos were stained with 5% v/v Giemsa solution (Invitrogen) in distilled water for 10 min. Chromosome spreads were examined under a ×100 oil magnification objective and chromosomal complements were determined for each embryo which was classified as haploid (1n), diploid (2n), polyploid (≥3n), mixoploid (embryos with blastomeres of different ploidies), and aneuploid. Only those spreads which were clearly in metaphase were analyzed.
Enucleation and ICI
After 21 h of IVM, MII oocytes were denuded and stained with 1 μg mL−1 of H 33342 for 10 min. Immediately, oocytes were transferred into 50 μL microdroplets of Hepes-TALP supplemented with 0.3 g mL−1 BSA, under mineral oil, in 100 × 20 mm tissue culture dishes (430167; Corning, NY, USA) and mechanically enucleated using a Narishige hydraulic micromanipulator (Narishige Sci., Tokyo, Japan) mounted on a Nikon Eclipse E-300 microscope (Nikon, Melville, NY, USA). Enucleation was performed using a 20 μm internal diameter pipette. Metaphase chromosomes were visualized under ultraviolet light (<10 sec) and aspirated into the pipette with a minimal volume of oocyte cytoplasm. Chromosome removal was confirmed by the presence of stained MII chromosomes inside the pipette.
Cumulus cells obtained from in vitro matured COCs were used as nuclear donors and microinjected, as previously described by Wakayama et al. (1998). Briefly, COCs were treated with hyaluronidase solution (H-4272; 1 mg mL−1 HEPES-TALP) to disperse cumulus cells, placed in trisodium citrate solution (A661048, Merck KGaA, Darmstadt, Germany 2.8% w/v in Double distilled water) for 5 min on ice, and transferred to 4 μL droplets of 10% v/v polyvinylpyrolidone solution (PVP; 99219; Irvine Sci., Irvine, CA, USA) in HEPES-TALP for injection. The cell membrane was broken by gently aspiration in and out of the injection pipette (inner diameter of 8 μm) and a single cell was injected into the cytoplasm of each enucleated oocyte. Aspiration was used to break the oolemma. The somatic cell and the aspirated ooplasm were then expelled into the oocyte with a minimal volume of PVP. Smaller somatic cells were used as donors. Subsequently, reconstituted eggs were cultured in 100 μL droplets of maturation medium for 2 h to allow nuclear reprogramming and randomly assigned to the following activation treatments: Io-DhL1/CB; Io-DhL1/long CB (treatment Io-DhL1/CB followed by exposure to CB 5 μg mL−1 in 2 mL of TCM-199 medium for 3 h); and Io-DMAP.
Statistical analysis
Each experiment was repeated at least three times. Differences among treatments in each experiment were determined by Fisher's exact test using Graph Pad PRISM® software 5.01 version. Differences between blastocyst total cell numbers were analyzed for significance using the one-way ANOVA test or the Kruskal–Wallis test when data did not fit a Gaussian distribution, using InfoStat® software 2008 version. For all analyses a difference of p < 0.05 was considered to be significant.
Results
Experiment 1: Pronuclear dynamics and second PB extrusion of oocytes activated with DhL
Pronuclear dynamics and extrusion of the second PB of MII oocytes were studied after the following activation treatments: Io-DhL1, Io-DhL1/CB, Io-DhL5, Io-DhL5/CB, Io-DMAP and Io. An IVF group was included. Pronuclear stages of 1125 oocytes or zygotes were evaluated at 4, 11, and 17 h.p.a., considering Io exposure as time 0 (Fig. 1). IVF evaluation hours were corrected for sperm penetration time, which was estimated to occur around 4 h after the beginning of sperm–oocyte coincubation (data not shown). Io-alone-treated oocytes formed very few pronuclei at any time (0; 5.9 and 1.9% for 4, 11, and 17 h.p.a., respectively). On the other hand, Io-DMAP treatment induced significantly higher rates of one PN formation (75.4%) at 4 h.p.a., and this proportion did not increase with time (82.4 and 69.8% for 11 and 17 h.p.a., respectively) (Fisher's test, p < 0.05). DhL-treated and IVF oocytes showed significantly lower activation rates at 4 h.p.a. (1.6 to 14.1 and 14.8%, respectively) (p < 0.05). Pronuclear formation induced by the four DhL treatments took place between 4 and 11 h.p.a., (19.7 to 36.9%). No differences were found between PN rates of the DhL groups at 17 h.p.a. (25 to 37.5%). PN rates seen for DhL treatments at 17 h.p.a. were statistically lower than those observed for Io-DMAP (69.84%) and IVF (83.33%) groups, and higher than Io alone treatment (1.92%) (p < 0.05). However, only IVF oocytes continued PN formation from 11 to 17 h.p.a. (83.3%) (p < 0.05).

Pronuclear dynamics of bovine oocytes activated with Io-DhL1, Io-DhL1/CB, Io-DhL5, Io-DhL5/CB, Io-DMAP, and Io, and of IVF oocytes. h.p.a: hours postactivation. PN: pronuclei. IVF hours were corrected by sperm penetration time.
Rates of second PB emission were 82.7; 80.2, 54.4, 51.2, 7.1, and 96.6% for Io-DhL1, Io-DhL5, Io-DhL1/CB, Io-DhL5/CB, Io-DMAP, and Io, respectively. These percentages were determined over total activated oocytes within each group, including results from 11 and 17 h.p.a, as no differences were found between them. Inhibition of second polar body emission was higher for DhL and CB combined treatments than for Io-DhL treatments, but lower than Io-DMAP (p < 0.05).
Experiment 2: In vitro development of parthenogenetic embryos
Metaphase II oocytes were exposed to the same treatments described in the first experiment and in vitro cultured. Developmental rates and blastocyst cell numbers were determined (Table 1). All DhL treatments showed significantly lower cleavage rates than the control groups (Io-DMAP and IVF treatments) (p < 0.05). The Io-DhL1 treatments, combined or not with CB, induced higher rates of morulae and blastocyst over cleaved embryos than Io-DhL5 (p < 0.05). Although developmental rates in all DhL treated groups were lower than those observed in Io-DMAP and IVF groups, Io-DhL1 (combined or not with CB) showed blastocyst-cleaved embryo ratios that were not statistically different from those of control groups (p > 0.05). Finally, blastocysts produced with Io-DhL1/CB treatment showed the highest total cell number, which was significantly different from other DhL treatments but not from Io-DMAP and IVF groups.
Io-DhL1: activation with 5 μM ionomycin plus DhL 1 μM; Io-DhL5: activation with 5 μM ionomycin plus DhL 5 μM; both alone or combined with 5 μg mL−1 Cytochalasin B (Io-DhL1/CB and Io-DhL5/CB respectively); Io: 5 μM ionomycin alone; Io-DMAP: ionomycin followed by 1.9 mM 6-DMAP.
Percentage of morulae and blastocysts are calculated over cleaved embryos.
:Values with different superscripts in the same column differ significantly (p < 0.05).
On the basis of these results, a concentration of 1 μM DhL was chosen for the following experiments.
Experiment 3: Ploidy of parthenogenetic embryos
Karyotypes of embryos activated with Io-DhL1, Io-DhL1/CB and a new treatment, Io-DhL1/long CB, were studied (Table 2 and Fig. 2). Treatment Io-DhL1/long CB was added to reduce the rates of second PB extrusion observed in Experiment 1. As expected, prolonged exposure to CB diminished haploid embryo rates, reaching values similar to Io-DMAP. Io-DhL1/long CB treatment produced the highest rates of diploid embryos, with minor percentage of aneuploidies. Finally, all DhL treatments showed polyploidy rates that were significantly lower than those occurring in the Io-DMAP group (p < 0.05). The CB addition did not raise chromosomal anomaly percentages in any case.
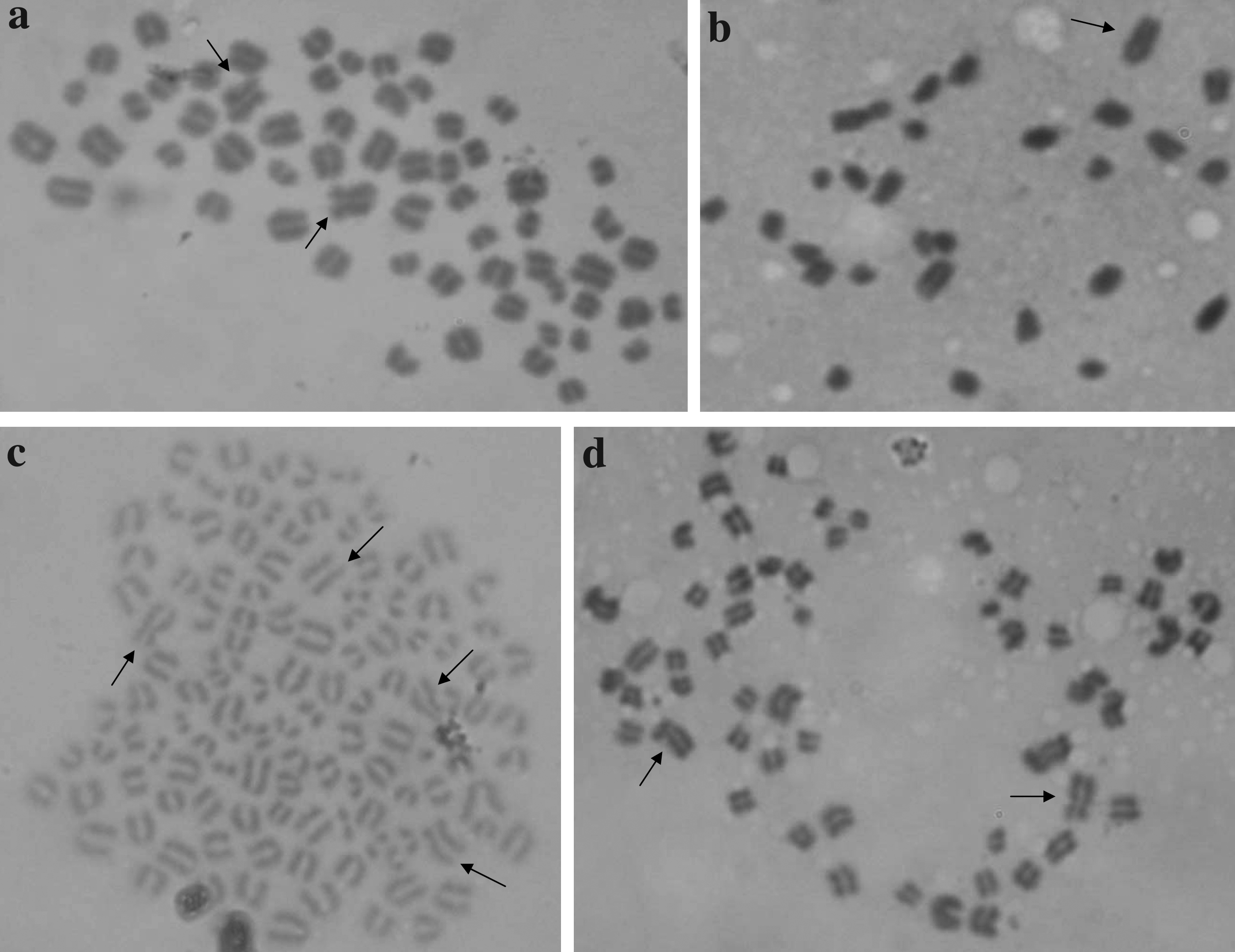
Metaphase spreads of bovine parthenogenetic embryos activated with: (
Io-DhL1: activation with 5 μM ionomycin plus DhL 1 μM; Io-DhL1/CB: activation with 5 μM ionomycin plus DhL 1 μM combined with 5 μg mL−1 Cytochalasin B; Io-DhL1/long CB: Io-DhL1 plus lengthened (6-h) exposure to CB; Io-DMAP: ionomycin followed by 1.9 mM 6-DMAP.
Values with different superscripts in the same column differ significantly (p < 0.05).
Experiment 4: In vitro development of SCNT bovine embryos activated with DhL and CB
Treatments Io-DhL1/CB, Io-DhL1/long CB and Io-DMAP were employed to activate reconstituted embryos. Parthenogenetic control groups were also included. Reconstituted and parthenogenetic embryos were in vitro cultured for 8 days to determine developmental rates and blastocyst cell numbers (Table 3). Both DhL and CB combined treatments induced reconstituted morulae and blastocyst rates that were similar to those induced by Io-DMAP (Fig. 3). Prolonged exposure to CB did not negatively affect embryo development to the blastocyst stage. However, reconstituted blastocyst cell numbers were lower for Io-DhL1/long CB treatment compared to Io-DhL1/CB and Io-DMAP groups (p < 0.05). Such observation was not valid for parthenogenetic embryos.
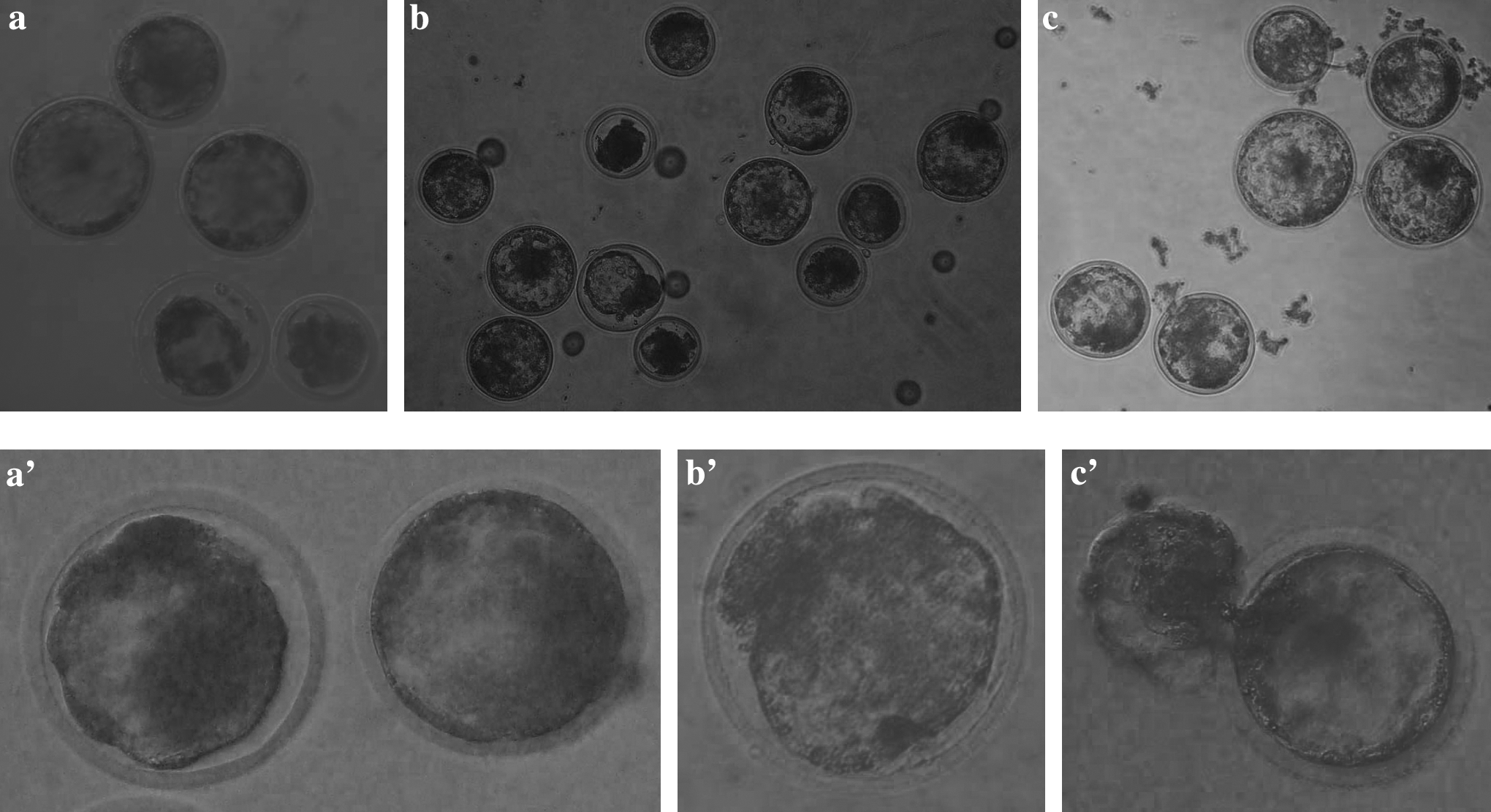
Parthenogenetic (
(+) Reconstituted embryos by enucleation and intracytoplasmic cell injection. (−) Parthenogenetic control groups. Io-DhL1/CB: activation with 5 μM ionomycin plus DhL 1 μM combined with 5 μg mL−1 Cytochalasin B; Io-DhL1/long CB: treatment Io-DhL1/CB followed by additional exposure to 5 μg mL−1 Cytochalasin B for 3 h; Io-DMAP: ionomycin followed by 1.9 mM 6-DMAP.
Percentage of morulae and blastocysts are calculated over cleaved embryos.
Values with different superscripts in the same column differ significantly (p < 0.05).
Discussion
In this study we demonstrated that DhL induced parthenogenetic activation of bovine oocytes, with pronuclear dynamics that were more similar to those produced by IVF than the widely employed DMAP activating agent. Moreover, parthenogenetic embryos derived by DhL activation exhibited lower rates of polyploidy than those treated with DMAP. For the first time, a combined treatment of Dehydroleucodine and Cytochalasin B was employed to induce development of cloned and parthenogenetic embryos to the blastocyst stage.
Pronuclear dynamics of activated and in vitro fertilized embryos were compared. Our results demonstrated that DhL induces PN formation, although in a more protracted time span than 6-DMAP, showing a nuclear dynamic profile that was more similar to that seen in oocytes subjected to IVF. Some studies have reported the presence of a fully decondensed PN at 4 h.p.a. or even earlier after 6-DMAP treatment (Liu and Yang, 1999; Susko-Parrish et al., 1994; Szöllösi et al., 1993), while in embryos produced by IVF, pronucleus is observed around 12–18 h postfertilization (Liu and Yang, 1999); our results are consistent with this observation.
In Experiment 1, we also observed that DhL allows second PB extrusion, suggesting it would induce haploid parthenogenetic activation. On the other hand, second PB extrusion and PN formation rates induced by 6-DMAP agree with previous reports, showing that PB emission is inhibited through the destruction of the metaphase plate, and gives place to the formation of a single diploid PN (Liu et al., 1998a; Susko-Parrish et al., 1994). On the basis of Experiment 1 results, we could suggest that, unlike DMAP, DhL does not exert its action over cytoskeleton components, because it does not inhibit second PB extrusion. However, further analysis will be necessary to fully understand the mechanism of Dehydroleucodine activation of oocytes.
Experiment 2 evaluated in vitro development rates of parthenogenetic embryos. Results showed that a concentration of 1 μM DhL is able to induce higher rates of development to the blastocyst stage than 5 μM DhL. Total blastocyst cell numbers did not differ significantly between DhL and control groups, indicating equivalent embryo quality according to this parameter (Van Soom et al., 1997). Independently of the embryo production system employed, blastocysts cell numbers were low. We attribute this to the culture conditions employed (SOFaa without coculture, under high oxygen tension), reported to diminish total cell number respect to low oxygen tension (Yuan et al., 2003). All DhL treatments showed significantly lower developmental rates than Io-DMAP and IVF groups. As DhL is supposed to induce mainly haploid activation (determined by second PB extrusion evaluation) and haploid embryos have less developmental capacity than diploid ones (Kawarsky et al., 1996; Liu et al., 2002; Lagutina et al., 2004), we suggest ploidy as a possible reason for the lower in vitro development of DhL embryos with respect to those treated with DMAP. Therefore, we directly evaluated this parameter.
Results indicated that Io-DhL1 and Io-DhL1/CB induce higher rates of haploid activation than DMAP. This information agrees with the second PB extrusion data. Despite this, these treatments produced rates of diploid embryos that did not differ from Io-DMAP, whereas Io-DMAP treatment presented higher rates of polyploidy than all DhL treatments. A high incidence of polyploidy has been observed by other authors at the blastocyst stage of parthenogenetic (De La Fuente and King, 1998; Winger et al., 1997) and SCNT bovine embryos (Bhak et al., 2006; Yoo et al., 2003) after DMAP activation. Some have attributed these anomalies to the accelerated pronuclear formation and premature DNA synthesis observed in DMAP-activated oocytes, followed by karyokinesis without cytokinesis during the first cell cycle (De La Fuente and King, 1998). To improve DhL diploidy rates, a new treatment, Io-DhL1/long CB was included. Prolonged exposure to CB (6 h) raised the inhibition of second PB extrusion of parthenogenetic embryos from 56.5 to 92.6% (data not shown), which is consistent with previous reports for long CB exposure (Liu et al., 1998b). Ock and Rho (2008) reported that a 5-h exposure to CB combined with cycloheximide could induce high rates of diploidy. In the present work, Io-DhL1/long CB treatment induced significantly more diploid parthenogenetic embryos than Io-DMAP and lower rates of polyploidy and aneuploidy.
On the basis of in vitro development, second PB extrusion and diploid embryo rates, Io-DhL1/CB and Io-DhL1/long CB treatments were selected to assist nuclear transfer. Results of Experiment 4 demonstrate that both DhL treatments (Io-DhL1/CB and Io-DhL1/long CB) induce rates of SCNT blastocyst development which do not differ from that of Io-DMAP. Unexpectedly, blastocysts cell number after SCNT assisted by Io-DhL1/long CB was lower than for Io-DhL1/CB and Io-DMAP. Our group previously reported activation of classical SCNT bovine embryos with Ionomycin plus DhL (Vichera et al., 2009). In the present study, we established a new oocyte activation protocol (Io-DhL1/long CB) to produce high rates of diploid embryos to assist nuclear transfer procedures. To date, the production of viable offspring by nuclear transfer remains inefficient because only a low percentage of the reconstructed blastocysts (0.5–5%) can develop to term (Cibelli et al., 1998; Kato et al., 1998; Wells et al., 1999a,b; Wilmut et al., 1997). This is mainly due to the high rates of abortion observed in cattle (Heyman et al., 2002), partly as a consequence of chromosomal anomalies. Additionally, embryonic loss due to chromosomal anomalies was suggested to take place after the blastocyst stage in the bovine embryo (De La Fuente and King, 1998; Kawarsky et al., 1996) and reported live born-cloned calves are chromosomally normal (Kato et al., 1998), indicating that embryos with abnormal ploidy are eliminated before birth. As the activation protocol is one of the steps of the nuclear transfer procedure that influence the ploidy of the resulting embryos, we believe that our new activation protocol could help to diminish this problem.
Moreover, SCNT is becoming more important for regenerative medicine and basic biology than for producing live animals (Wakayama, 2007). It has been shown that embryonic stem cells produced by NT are equivalent to ES cells derived from fertilized embryos and they can be generated from a variety of mouse genotypes and cell types (Wakayama, 2007). However, loss and reduced pluripotency in the ICM of cloned blastocysts has been reported (Boiani et al., 2002). New activating agents, such as DhL, could help to improve parthenogenic and NT embryonic stem cell lines outcome.
In conclusion, we demonstrated that Dehydroleucodine induces parthenogenetic activation of bovine oocytes, with early pronuclear dynamics that are similar to IVF embryos. We designed a combined treatment with Dehydroleucodine and Cytochalasin B, which induces higher rates of diploid parthenogenetic embryos and lower rates of polyploidies than Io-DMAP, and that results in the development of cloned embryos to the blastocyst stage at the same rate as the widely used treatment, Io-DMAP.
Footnotes
Acknowledgments
This work was supported by Agencia de Promoción Científica y Tecnológica (PICT Redes No. 35142 2005-2009), by Fundación Juan B. Sauberan and by Universidad de Buenos Aires (UBACYT G808). The authors thank C.I.A.L.E. for providing bull semen, and the Instituto de Embriología e Histología de Mendoza, Argentina for providing purified Dehydroleucodine. We are also thankful to Karina Hodara for assistance with statistical analysis.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.