
Abstract
Abstract
Oxygen consumption reflects overall metabolic activity of mammalian embryos. We measured oxygen consumption in individual porcine somatic cell nuclear transfer (SCNT) and in vitro-fertilized (IVF) embryos by modified scanning electrochemical microscopy. Oxygen consumption in IVF embryos rapidly increased at day 5 of the blastocyst stage (D5BL). IVF embryos that consumed >0.81 × 1014/mol sec−1 of oxygen at D5BL exhibited significantly higher hatching and hatched rates at D7BL, whereas D5BL SCNT embryos using porcine fetal fibroblasts did not show an increase in oxygen consumption until D7BL. The numbers of inner cell mass and trophectoderm (TE) cells and incidence of apoptosis did not significantly differ between IVF and SCNT embryos at D5BL. At D7BL, a significant lower number of TE cell and higher incidence of apoptosis were observed in SCNT than in IVF embryos; this significantly correlated with their oxygen consumption at D5BL. Use of cumulus cells as donor cells neutralized the low oxygen consumption in SCNT embryos at D5BL, regardless of the difference between the recipient cytoplasm and donor nucleus. Some of SCNT embryos at D7BL were retrieved the hatching completion and were improved the number of TE cell and apoptosis incidence by using cumulus cells. Thus, anomalous oxygen consumption in porcine SCNT embryos at D5BL could be sign of limited hatchability, which may be responsible for the low TE cell number and high apoptosis incidence.
Introduction
In general, oxygen consumption is a good indicator of the overall metabolic activity of individual embryos because ATP is mostly generated by oxidative phosphorylation, a process in which oxygen plays a key role (Houghton et al., 1996; Leese, 2003; Thompson et al., 1996; Thompson, 2000; Trimarchi et al., 2000). Further, our previous studies have shown that oxygen consumption in individual in vitro-fertilized (IVF) bovine embryos directly correlated with the subsequent embryonic development (Abe and Hoshi, 2003; Shiku et al., 2001). These studies were performed using modified scanning electrochemical microscopy (SECM). The SECM system appears to be a reliable, noninvasive, and a highly sensitive method for the measurement of oxygen consumption in individual embryos (Shiku et al., 2001). In the present study, for a better understanding of the underlying cause of low cloning efficiency in pigs, we measured oxygen consumption by using SECM to evaluate energy metabolism in porcine SCNT embryos.
Materials and Methods
In vitro maturation of oocytes
Ovaries of prepubertal gilts from three-way crossing (Duroc boars × Landrace-Large White sows) were collected at a local slaughterhouse and transported to the laboratory within 2 h in a container containing warm saline. Porcine oocytes were aspirated from the antral follicles, 3 to 6 mm in diameter, using an 18-gauge needle attached to a 5-mL disposable syringe. Cumulus–oocyte complexes (COCs) with uniform ooplasm and compact cumulus cell mass were selected in modified Dulbecco's phosphate-buffered saline (PBS) medium containing 5.56 mM glucose (Wako, Osaka, Japan), 0.33 mM sodium pyruvate (Wako), 1% (v/v) antibiotic antimycotic solution (Sigma Chemical Co., St. Louis, MO, USA), and 4 mg/mL fatty acid-free bovine serum albumin (BSA; 128K7400, Sigma) (PB1) (Quinn et al., 1982). After washing in PB1, the COCs were cultured in BSA-free NCSU-23 medium (Miyoshi et al., 2000) for 44 h at 39°C in a highly humidified atmosphere of 5% CO2 in air. COCs were sequentially cultured as described by Funahashi et al. (1993). For the first 22 h of maturation culture, the medium was supplemented with 10 IU/mL equine Chronic Gonadotropin (eCG; Serotropin; Teikokuzouki, Tokyo, Japan), 10 IU/mL human chorionic gonadotropin (hCG; Puberogen; Sankyo, Tokyo, Japan), 0.1 mg/mL cysteine (Sigma), 1 mM dibutyryl cyclic AMP (Sigma), and 10% (v/v) porcine follicular fluid. The culture for the subsequent 22 h was carried out in the same medium without eCG and hCG. After culturing, expanded cumulus cells of the COCs were removed by vortexing for 30 sec in PB1 containing 1 mg/mL hyaluronidase (Sigma), and the oocytes with the first polar body were selected as matured oocytes under a stereomicroscope, and they were transferred into PB1 and used for the experiments.
Donor cells
The present study was approved by the Ethics Committee for Care and Use of Laboratory Animals for Biomedical Research of the Graduate School of Agricultural Science, Tohoku University. Donor cells were isolated as described by Wakai et al. (2008). In brief, pig fetuses of Goettingen breed (CSK, Suwa, Japan) were collected from the sow on day 56 of pregnancy. Each fetus was decapitated and eviscerated. The remaining tissues were washed in PBS (Sigma) and then digested with 0.1% (w/v) trypsin-ethylenediaminetetraacetic acid (EDTA) (Sigma) for 45 min at 38.5°C. After digestion, the cells collected as porcine fetal fibroblast (pFF) cells were cultured in Dulbecco's modified Eagle medium (DMEM, Sigma) supplemented with 10% (v/v) fetal bovine serum (FBS; A51408X, Gemini Bio-Products, Chalabasas, CA, USA). The culture medium was replaced every 2 days until the cells reached confluence. Subsequently, the cells were harvested with 0.1% (w/v) trypsin in PBS containing 0.5 mM EDTA for 5 min at 38.5°C, frozen with a cryoprotectant (Cellbanker, Zenyaku, Tokyo, Japan) and stored in liquid nitrogen (passage 0). For an overall experiment, fetal fibroblasts derived from one female Goettingen breed were used (Wakai et al., 2008). However, in the only experiment for evaluating potential sex specific effects (Supplementary Table 1; see online supplementary material at www.liebertonline.com); we used male fetal fibroblasts from littermates with this female fetal fibroblasts. Prior to the start of experiments, the cells were thawed and cultured in DMEM plus 10% FBS and used between passages 4 and 9. The fibroblasts were cultured for 1 week after they had attained confluence and were then used as donor cells. As another type of donor cell to use, the porcine cumulus cells (pCC) were prepared. After in vitro culture of COCs for 44 h, cumulus cells were isolated from oocytes by pipetting in PB1 containing hyaluronidase. Then they were transferred to PB1 for use as donor cells. They were injected to enucleated oocytes either from the same female (pCC-Auto) or from a different female (pCC-Allo).
Through the experiments, donor cells were used pFF. We further used pCC in Experiments 6 and 7.
Nuclear transfer
We produced the SCNT embryos by the reconstruction of enucleated oocytes with somatic cells in accordance with previous work, in which we had already confirmed the reconstructed embryos to be survivable until adult nuclear transplant miniature pigs (Wakai et al., 2008). The cumulus-free oocytes were stained with 5 μg/mL Hoechst 33342 (Sigma) for 5 to 10 min at 39°C and transferred into PB1 containing 2.5 μg/mL cytochalasin D (Sigma). Enucleation was performed by aspirating the first polar body and a small volume of the adjacent cytoplasm with a enucleation pipette of 20 μm diameter using a piezo-driven unit (Primtech, Tokyo, Japan), and its success was confirmed by visualizing the presence of metaphase plates within the removed cytoplasm under ultraviolet (UV) light. The enucleated oocytes were washed three times with porcine zygote medium-3 (PZM-3) (Yoshioka et al., 2002) and transferred to a 100-μL droplet of PZM-3 for the subsequent microinjection or fusion of the donor nucleus. The donor cells were transferred to a 50 μL droplet of PB1. To produce SCNT embryos by microinjection of donor cells, their plasma membranes were broken by gently aspirating them in and out of the injection pipette of 10 μm in diameter before microinjection into cytoplasm. To produce SCNT embryos by the electric fusion, intact donor cells were placed in the perivitelline space of recipient oocytes through the same slit in the zona pellucida that had been made for enucleation. Then they were fused with the recipient oocytes using LF 101 (TR Tech, Tokyo, Japan) by stimulating direct current pulses of 150 V/mm for 50 μsec in 0.28 M mannitol solution supplemented with 0.1 mM MgSO4 and 0.01% (w/v) PVA (Yin et al., 2003). Throughout the experiments, nuclear transfer was performed by a microinjection method of donor cells into recipient cytoplasts. Only in Experiment 3 did we further perform the electrofusion method.
Activation and culture of SCNT embryos
The activation of embryos was performed as described by Yamanaka et al. (2007). In brief, to activate the SCNT embryos, the reconstructed embryos at 3 h after injection or fusion of donor cells were treated with 15 μM ionomycin (Sigma) in PZM-3 containing fatty acid-free BSA (Yoshioka et al., 2002) for 20 min at 38.5°C in 5% CO2 in humidified air and then washed with PZM-3. Next, 20 SCNT embryos were cultured in a 100-μL droplet of PZM-3 containing 5 μg/mL cycloheximide (Sigma) with cytochalasin D (Sigma) for 5 h at 38.5°C in 5% CO2 in humidified air and then washed five times with cycloheximide-free PZM-3 (Sugimura et al., 2008). Last, the 20 embryos were transferred to a 100-μL droplet of PZM-3 and cultured at 38.5°C in an atmosphere of 5% CO2 in air.
In vitro fertilization
IVF was also performed with in vitro-matured (IVM) oocytes. After the IVM, oocytes were washed three times with fertilization medium (TU medium) (Miyoshi et al., 1999), and 30 to 40 oocytes were transferred to a 100-μL droplet of IVF medium. Cryopreserved semen from Duroc boars was thawed, and spermatozoa were washed twice by centrifugation (1000 × g for 4 min) in Dulbecco's PBS supplemented with 1 mg/mL BSA. Spermatozoa were resuspended in IVF medium, and 20 to 30 μL of the suspension was added to the drop containing oocytes to give a final concentration of 3 × 105 cells/mL. At 6 h postinsemination, 20 oocytes were washed three times in 100 μL droplet of PZM-3 and cultured in 100 μL droplet of PZM-3 at 38.5°C in 5% CO2 in air. In our laboratory IVF system using the boar sperm, the rate of penetration and normal fertilization (both male and female pronuclei and two polar bodies) was 95.1 and 61.5%, respectively (Yamanaka et al., 2009).
Collection of in vivo produced embryos
Protocol of “surgical collection of embryos” was according to manual on pig embryo transfer (Kashiwazaki, 1993). Day 160 to 220 prepubertal gilts (Large-White) were treated with 1500 IU eCG, followed 72 h later by 500 IU hCG. Artificial insemination (AI) was performed three times from 24 h after hCG. Embryos on day 4 and 5 (day 0 = the day of first AI) were recovered surgically from the gilts.
Production of parthenogenetically activated embryos
To generate different types of parthenogenetically activated embryos (PA), IVM porcine oocytes without manipulation were activated by using the same methods as those used for generating SCNT embryos. From these oocytes, a small volume of ooplasm was removed, and a minimal amount of PB1 medium was injected (Sham-PA); an enucleated pFF cell was also injected (SCI-PA) in these cells. Enucleation of pFF was performed using the method described by Hayashi et al. (1991) with slight modification. Upon reaching confluence, pFF cells were enucleated by centrifugation (23,000 × g for 30 min) in DMEM with cytochalasin B (10 μg/mL) and Hoechst 33342. Enucleation was confirmed by visualizing the cells under UV light. Subsequently, plasma membranes of the cells, in which enucleation was confirmed, were disrupted by gently aspirating the cells in and out of the beveled pipette; their cytoplasts were injected into the oocytes.
Measurement of oxygen consumption
Oxygen consumption by individual porcine embryos was noninvasively measured by the SECM system (HV-405; Hokuto Denko Co., Tokyo, Japan) (Abe and Hoshi, 2003; Shiku et al., 2001). Embryos were transferred singularly to a plate filled with 5 mL of embryo respiration assay medium-2 (ERAM-2; Research Institute for the Functional Peptides, Yonezawa, Japan) and dropped individually to the bottom of the microwell. The medium temperature was maintained at 37°C by placing on a warming plate on the microscope stage. The measurement of oxygen consumption was carried out according to the procedure previously described by Shiku et al. (2001). Briefly, Pt-microdisc electrodes, sealed in a tapered soft-glass capillary (PG10165-4: World Precision Instruments, Sarasota, FL), were fabricated according to the procedure described by Matsue et al. (1993). The tip potential was held at −0.6 V versus Ag/AgCl with a potentiostat to monitor the local oxygen concentration in the solution. The tip scanning rate was 31.0 μm/sec. The XYZ-stage and the potentiostat were controlled by a notebook computer. Voltammetry of the Pt-microdisc electrode in ERAM-2 solution showed a steady-state oxygen reduction wave. No response from other electrochemically active species was observed near the embryo surface. The oxygen consumption rate of embryos was calculated by software, in which the oxygen concentration difference between the bulk solution and sample surface (ΔC), and the oxygen consumption rate (F) of a single sample were estimated according to spherical diffusion theories (Shiku et al., 2001). We repeatedly scanned the electrode back and forth, three times, to estimate the mean ± standard deviation (n ≥ 4) of the ΔC for each sample.
Differential staining
The cell allocation of blastocysts was assessed by differential staining of the inner cell mass (ICM) and trophectoderm (TE) cells according to the staining procedure described by Thouas et al. (2001) with slight modification. Briefly, TE cells of the blastocysts were stained with 100 μg/mL propidium iodide fluorochrome containing a permeabilizing solution of 2% (v/v) Triton X-100 ionic detergent for 20 sec. Blastocysts were then incubated overnight at 4°C in a second solution of 25 μg/mL Hoechst 33342 in ethanol for fixation. Fixed and stained whole blastocysts were mounted and assessed for cell number using an epifluorescence microscope.
Detection of apoptosis-positive cells by TUNEL assay
Detection of apoptosis-positive cells by TUNEL assay was performed as described by Yamanaka et al. (2009). Blastocysts from SCNT and IVF were washed three times in PBS supplemented with 0.1% polyvinylpyrrolidone and fixed in 2% (w/v) paraformaldehyde and 0.2% (v/v) Triton-X PBS solution for 40 h at room temperature (RT). A commercially available kit (ApopTag; Intergen, Purchase, NY, USA) was used for the detection of apoptosis-positive cells. For the positive control, blastocysts were treated with DNase I (10 IU/mL, Sigma). After washing with PBS supplemented with 0.1% (w/v) polyvinyl alcohol three times for 10 min each, the blastocysts were incubated in the equilibration buffer of the kit for 20 sec at RT. Then, they were incubated at 37°C for 2 h in a moist chamber with 70% (v/v) reaction buffer containing 30% (v/v) terminal deoxynucleotidyl transferase, digoxigenin-11-dUTP. The reaction was stopped by addition of 3% (v/v) stop/wash Buffer (Intergen) at 37°C for 10 min. After washing with PBS including 0.2% Triton-X for four times for 2 min each, they were incubated with antidigoxigenin antibody conjugated to horseradish peroxidase at RT for 1 h. After washing with 0.2 % Triton-X and 0.1% PVA PBS four times, blastocysts were stained with 10 μg/mL propidium iodide in PBS for 1 h at RT. All samples were examined under a laser-scanning confocal microscope (MCR-1024; BIO-RAD Hercule). The numbers of apoptosis-positive nuclei and total number of nuclei in blastocysts were determined.
Individual culture of blastocysts
At day 5 after in vitro culture, the blastocysts were collected and washed three times in 100-μL droplets of PZM-3. Thereafter, they were individually cultured in 96-well plates containing 100 μL of PZM-3 for 48 h at 38.5°C in 5% CO2 in humidified air. The individual culture system was used in Experiments 4 and 5 (shown in Figs. 2, 3, and 4). Both IVF and SCNT blastocysts derived from the individual culture system did not differed from those of conversional group culture system, which was used in Experiment 7 (shown in Table 5), on the embryo cell numbers and apoptosis incidence.
Experimental Studies
In Experiment 1, the rates of cleavage and blastocyst formation in IVF and SCNT embryos were assessed on day 2 and on days 5, 6, and 7 (the day of IVF was defined as day 0) of in vitro culture, respectively. In each trial, IVF and SCNT embryos were cultured in each drop.
In Experiment 2, oxygen consumption in the preimplantation IVF and SCNT embryos was measured at the two-cell stage on day 1 (2C); four-cell stage on day 2 (4C); morula stage on day 4 (MO); and blastocyst stages on days 5 (D5BL), 6 (D6BL), and 7 (D7BL). In each trial, IVF and SCNT embryos were cultured in each drop, and measured oxygen consumption at each stage. Some of embryos measured oxygen consumption at D5BL, D6BL, and D7BL were counted total cells number with Hoechst 33342 for calculating oxygen consumption per one cell. As physiological control, in vivo produced embryos at molura stage on day 4 (MO) and blastocyst stage on day 5 (D5BL) were also measured oxygen consumption.
In Experiment 3, to further explore some of the possible causes of lower oxygen consumption in the SCNT embryos, we examined the effects of the following four parameters on oxygen consumption of SCNT embryos: removal of ooplasm, contamination of cytoplasm of the donor cell, method of nuclear transfer (microinjection or electrofusion), and artificial activation. In each trial, IVF, PA, Sham-PA, SCI-PA, IN-SCNT, and EF-SCNT embryos were cultured in each drop. The embryos at D5BL were measured for oxygen consumption and then counted for total cell numbers with Hoechst 33342.
In Experiment 4, we determined the correlation between oxygen consumption in embryos at D5BL and their subsequent hatching competence at D7BL. In each trial, IVF and SCNT embryos were cultured in each drop and oxygen consumption was measured at D5BL. Furthermore, these blastocysts were individually cultured, and the developmental stage—unexpanded, expanded (i.e., expansion of the blastocoele and decrease in the thickness of the zona pellucida), hatching (i.e., hatching of the embryos from the zona pellucida), and hatched (i.e., those embryos that had hatched from the zona pellucida) (Kameyama, 1998)—was assessed at 48 h after extended culture (D7BL). Expanded and hatching of SCNT could not be strictly assessed. This was because the SCNT embryos already had mechanical slits or holes in their zona pellucid.
In Experiment 5, to gain an in-depth understanding of the reason behind the lower oxygen consumption in the SCNT embryos, we focused on the total, ICM, and TE cell numbers and apoptosis incidence of embryos. First, we examined the total, ICM, and TE cell numbers and apoptosis incidence of embryos at D5BL and D7BL. In each trial, IVF and SCNT embryos were cultured in each drop and oxygen consumption was measured at D5BL. Next, we determined the correlation between oxygen consumption in embryos at D5BL and their total, ICM, and TE cell numbers and at D7BL. In each trial, IVF and SCNT embryos were cultured in each drop and oxygen consumption measured at D5BL. Furthermore, these blastocysts were individually cultured, and total, ICM, and TE cell numbers and apoptosis incidence were assessed at D7BL.
In Experiment 6, to evaluate whether oxygen consumption was altered by the difference in donor cell types such as pFF and pCC, the effect of donor cells on oxygen consumption in SCNT embryos at D5BL was studied. In each trial, IVF and three types of SCNT (pFF, pCC-Auto, and pCC-Allo) embryos were cultured in each drop. The embryos at D5BL were measured oxygen consumption and then counted total cells number with Hoechst 33342.
In Experiment 7, hatchability, embryo cell numbers and apoptosis incidence of SCNT embryos with pFF and pCC at D7BL was evaluated. In each trial, two types of SCNT (pFF and pCC) embryos were cultured in each drop. SCNT embryos with pCC were derived from both pCC-Auto and pCC-Allo. On the days specified in Experiment 1, the rates of cleavage and blastocyst formation were calculated. The embryos at D7BL assessed the rate of hatched blastocyst, ICM, and TE cell numbers and apoptosis incidence.
Statistical analysis
Each experiment was performed in at least triplicate. The correlation coefficients between oxygen consumption and cell allocation or apoptosis were determined by simple regression analysis. Percentage data were arc-sine transformed, and other data were analyzed using analysis of variance (ANOVA), followed by the Bonferroni procedure (p < 0.05). The statistical significance of all data was examined using StatView (Abacus Concepts Inc., Berkeley, CA, USA).
Results
Experiment 1. In vitro development of porcine IVF and SCNT embryos
The cleavage rate did not significantly differ between the IVF and SCNT embryos (Table 1). On the other hand, at D5BL, D6BL, and D7BL, the blastocyst formation rates in the case of SCNT embryos were significantly lower than those in the case of IVF embryos (p < 0.05).
Different letters within the same stage indicate significant difference (p < 0.05).
IVF, in vitro-fertilized; SCNT, somatic cell nuclear transfer.
Experiment 2. Oxygen consumption in preimplantation IVF and SCNT embryos
As shown in Figure 1A, oxygen consumption, expressed as mean ± SD, in the preimplantation IVF embryos at the 2C and 4C stages was stable at a low value (0.35 ± 0.12 and 0.38 ± 0.08 × 1014/mol sec−1, respectively). Thereafter, oxygen consumption significantly increased at the MO stage (0.52 ± 0.11 × 1014/mol sec−1) compared to that at the 2C stage (p < 0.05). Subsequently, this value increased 1.5 times at D5BL (0.82 ± 0.18 × 1014/mol sec−1). In contrast, the SCNT embryos at D5BL did not exhibit any increase in oxygen consumption from the MO stage, even up to D7BL (MO, 0.41 ± 0.08 × 1014/mol sec−1; D5BL, 0.44 ± 0.12 × 1014/mol sec−1; and D7BL, 0.46 ± 0.11 × 1014/mol sec−1) (Fig. 1B). Furthermore, we compared the oxygen consumption calculated by per one cell of the IVF and SCNT embryos at the blastocyst stage (Fig. 1CD). The values in the case of the SCNT embryos were lower than those in the case of the IVF embryos at D5BL, D6BL, and D7BL (p < 0.05).
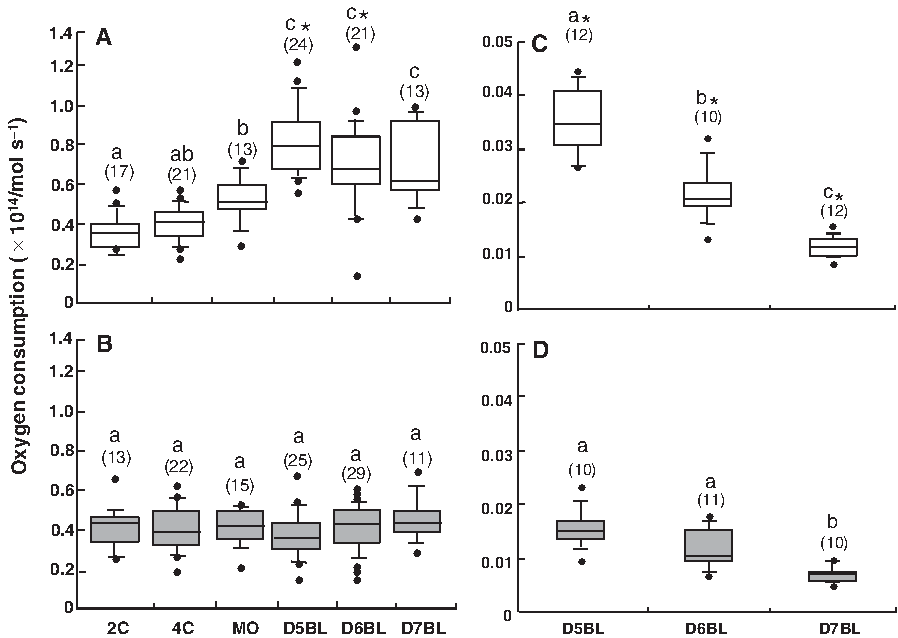
Oxygen consumption in the preimplantation in vitro-fertilized (IVF) and somatic cell nuclear transfer (SCNT) embryos. The box-plot graphs represent oxygen consumption in the preimplantation IVF (
From the MO stage to the D5BL stage, we found the significant difference in the way of changing of oxygen consumption between the IVF and the SCNT embryos (Fig. 1AB). For this confirmation, we further examined the oxygen consumption in in vivo-produced embryos at both the MO and D5BL stages. The result revealed that the oxygen consumption in in vivo produced embryos increased 1.5 times from the MO stage to the D5BL stage as well as in the IVF embryos (MO: n = 4; 0.67 ± 0.06 × 1014/mol sec−1 and D5BL: n = 8; 0.93 ± 0.14 × 1014/mol sec−1).
Experiment 3. Effect of removal of the ooplasm, contamination of the cytoplasm of the donor cell, method of nuclear transfer, and artificial activation on oxygen consumption at D5BL
As shown in Table 2, there were no significant differences with respect to the oxygen consumption and total cell number of embryos at D5BL between the IVF, PA, Sham-PA, and SCI-PA embryos; further, the values of oxygen consumption were significantly higher in these embryos than in the SCNT embryos (p < 0.05). Furthermore, there were no significant differences with respect to the oxygen consumption and total cell number of embryos at D5BL between the method of nuclear transfer; namely, the microinjection (IN-SCNT) and electrofusion (EF-SCNT) methods. Thus, the removal of ooplasm, contamination of the cytoplasm of the donor cell, method of nuclear transfer, and artificial activation did not influence anomalous oxygen consumption in the SCNT embryos.
Different letters within the same column indicate significant difference (p < 0.05).
PA, parthenogenetically activated embryos; Sham-PA, parthenogenetically activated embryos from which a small volume of ooplasm was removed and a minimal amount of PB1 medium was injected; SCI-PA, parthenogenetically activated embryos in which only the cytoplasm of the donor cell was injected; IN-SCNT, SCNT embryos generated by microinjection; EF-SCNT, SCNT embryos generated by electrofusion. Data are presented as mean ± SD.
Experiment 4. Correlation between oxygen consumption at D5BL and subsequent developmental competence
To clarify the correlation between oxygen consumption and subsequent development, the sample embryos that were used for analysis were aligned as previously described (Kameyama, 1998). We selected IVF and SCNT embryos at D5BL with similar morphology—in these embryos, the volume of the blastocoele cavity up to the vacuole was half that of the entire embryo (Fig. 2A). The oxygen consumption (mean ± SD) in the IVF embryos at D5BL definitely reflected their hatching ability at D7BL (Fig. 2B). However, the oxygen consumption was significantly lower at D5BL in the SCNT embryos than in the IVF embryos that underwent expansion at D7BL (p < 0.05).

Oxygen consumption on day 5 after in vitro culture and development of the embryo in the blastocyst stage on day 7. (
Next, on the basis of the results shown in Figure 2B, oxygen consumption in the IVF and SCNT embryos at D5BL was classified into four classes, as shown in Figure 2C: <0.59, 0.59–0.74, 0.75–0.93, and >0.93 × 1014/mol sec−1, representing embryos with the potential of undergoing one of the following developmental stages, respectively: unexpanded, expanded, hatching, and hatched. These four groups comprised 15.6, 31.3, 28.1, and 25% of the IVF embryos at D5BL, respectively. In contrast, at D5BL, most of the SCNT embryos were categorized as being in the unexpanded or expanded stages (60.9 and 34.8%, respectively), but none of them were classified as hatched embryos.
Experiment 5. Correlation between oxygen consumption and embryo cell numbers and apoptosis incidence
As shown in Figure 3A and B and Figure 4A and B, no significant differences with respect to the total, ICM, and TE cell numbers and incidence of apoptosis were observed between the IVF and SCNT embryos at D5BL. However, at D7BL, significant differences were observed between the IVF and SCNT embryos (p < 0.05) with regard to the following parameters: the total (IVF, 75.9 ± 27.5 vs. SCNT, 38.3 ± 13.0) and TE cell numbers (IVF, 65.8 ± 27.5 vs. SCNT, 29.9 ± 13.5); ICM/TE ratio (IVF, 0.18 ± 0.11 vs. 0.33 ± 0.22); and incidence of apoptosis (IVF, 4.8 ± 2.4 vs. SCNT, 19.7 ± 7.6).
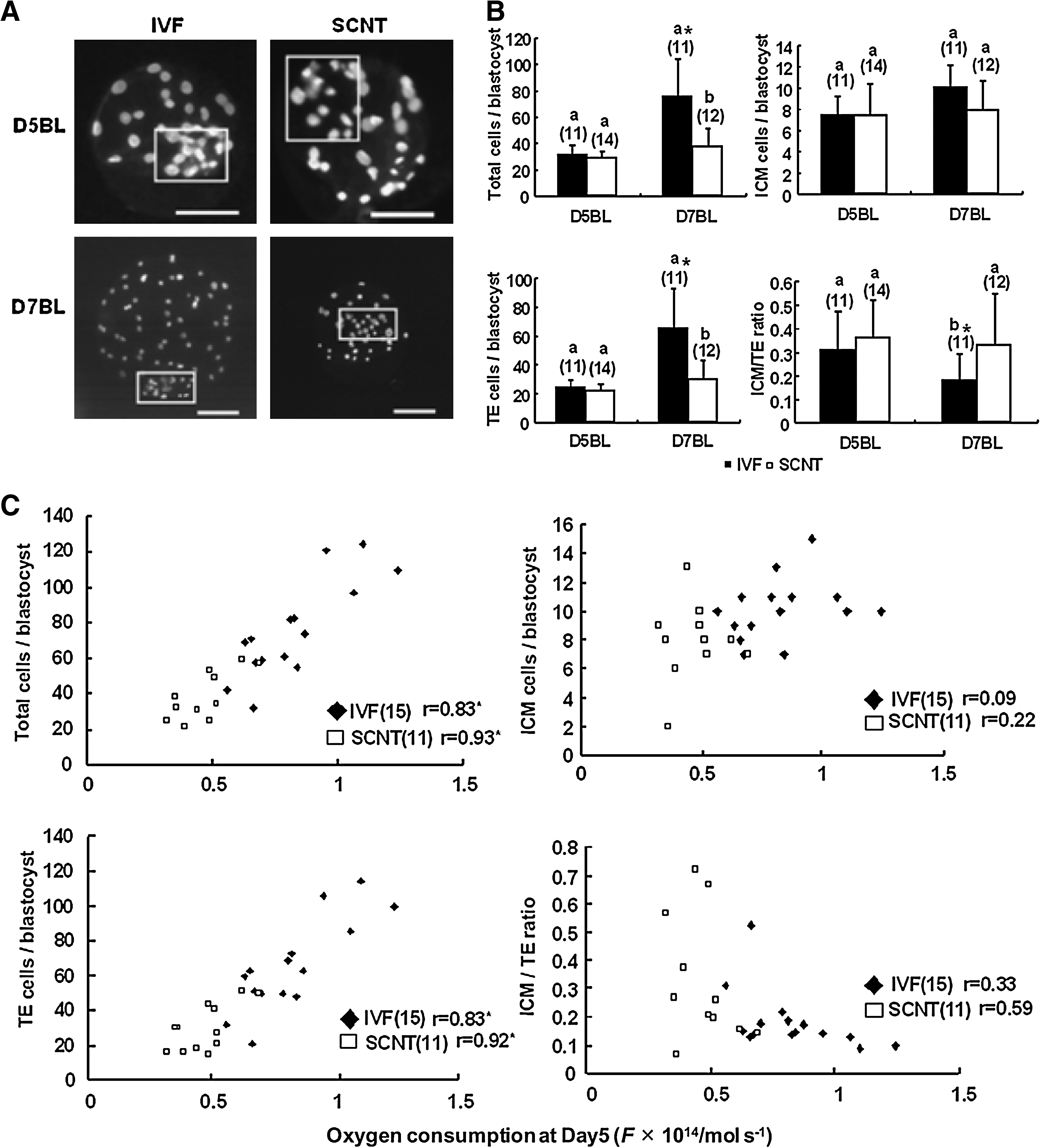
Total, inner cell mass (ICM), and trophectoderm (TE) cell numbers in the IVF and SCNT embryos at D5BL and D7BL and correlation between oxygen consumption at D5BL and the values of these parameters at D7BL. (

Number of apoptosis-positive cells in the IVF and SCNT embryos at D5BL and D7BL and correlation between oxygen consumption at D5BL and the number of apoptosis-positive cells at D7BL. (
Next, the correlation between oxygen consumption in embryos at D5BL and the total, ICM, and TE cell numbers and incidence of apoptosis in the embryos at D7BL was determined. As shown in Figure 3C and 4C, a significant correlation (p < 0.01) was observed between the oxygen consumption in embryos at D5BL and the total (IVF: r = 0.83, SCNT: r = 0.93) and TE cell numbers (IVF: r = 0.83, SCNT: r = 0.92) and incidence of apoptosis (IVF: r = 0.57, SCNT: r = 0.84) in embryos at D7BL in the IVF and SCNT embryos; however, the ICM cell number did not significantly differ between the IVF and SCNT embryos. This finding indicates that the oxygen consumption in the embryos at D5BL correlated with the total and TE cell numbers and incidence of apoptosis at D7BL.
Experiment 6. Effect of donor cell type on oxygen consumption in the SCNT embryos at D5BL
As shown in Table 3, oxygen consumption was significantly higher in the SCNT embryos generated using cumulus cells (pCC-Auto and pCC-Allo) as donor cells than in the pFF embryos; however, oxygen consumption was significantly lower in these embryos than in the IVF embryos (p < 0.05). Furthermore, oxygen consumption and the total cell number did not significantly differ between the pCC-Auto and pCC-Allo embryos.
Different letters within the same column indicate significant difference (p < 0.05).
pFF, SCNT embryos with fetal fibroblast; pCC-Allo and pCC-Auto represent allogeneic and autologous SCNT embryos with cumulus cells, respectively. Data are presented as mean ± SD.
Experiment 7. Hatchability, embryo cell numbers, and apoptosis incidence in SCNT embryos reconstructed with cumulus cells at D7BL
At D7BL, the hatched blastocyts were observed in pCC embryos (3/30), but not in the pFF embryos (0/27) (Table 4). Both the total and TE cell numbers of pCC embryos significantly increased compared with those of pFF embryos; in contrast, the apoptosis incidence in pCC embryos significantly decreased (p < 0.05) (Table 5).
Different letters within the same stage indicate significant difference (p < 0.05).
pFF and pCC represent SCNT embryos with fetal fibroblasts and cumulus cells, respectively.
Different letters within the same column indicate significant difference (p < 0.05).
pFF and pCC represent SCNT embryos with fetal fibroblast and cumulus cell, respectively. Data are presented as mean ± SD.
Discussion
To evaluate the overall metabolic activity in individual SCNT embryos during the preimplantation stages, we measured their oxygen consumption levels by SECM. Our results clearly indicate lower oxygen consumption in the SCNT embryos than in the IVF embryos at blastocyst stage (Fig. 1). Oxygen consumption in the IVF and in vivo-produced embryos rapidly increased from the MO stage to the D5BL stage. In contrast, no such increase in oxygen consumption was observed in the SCNT embryos.
In the present study, we hypothesized that low embryo numbers (Boiani et al., 2003; Koo et al., 2004) and high incidence of apoptosis (Hao et al., 2003) in the SCNT embryos might have resulted in the low oxygen consumption at D5BL. We, therefore, analyzed both the ICM and TE cell numbers and apoptosis incidence in the SCNT embryos. Although at D7BL significantly lower embryo cell numbers and higher apoptosis incidence were observed in the SCNT embryos than in the IVF embryos, these differences were not significant at D5BL (Figs. 3 and 4). Thus, the low embryo cell numbers and high apoptosis incidence in the SCNT could not have accounted for the low oxygen consumption in the SCNT embryos at D5BL. Furthermore, we examined the effects of the nuclear transfer methods on the oxygen consumption in the SCNT embryos—removal of ooplasm; methods of introduction of the donor nuclei into the enucleated oocyte, such as injection or electrofusion; contamination of the cytoplasm of the somatic cell, artificial activation, and treatment with cytochalasins. We found that the abovementioned methods did not appear to be the major causes of lower oxygen consumption in the SCNT embryos at D5BL (Table 2). The sex difference of donor cells also did not affect the lower oxygen consumption at D5BL (male pFF, 0.45 ± 0.11 × 1014/mol sec−1 vs. female pFF, 0.48 ± 0.16 × 1014/mol sec−1) (Supplementary Table 1). This is in accordance with the previous reports, which show no significant difference between sexes on oxygen consumption of bovine in vitro produced blastocysts (Agung et al., 2005; Lopes et al., 2005). On the other hand, the oxygen consumption levels in the SCNT embryos at D5BL that were reconstructed with cumulus cells was neutralized, regardless of the difference between the recipient cytoplasm and donor nucleus in an individual. This result indicates that the type of donor nucleus might be responsible for the low oxygen consumption in the SCNT embryos at D5BL. In a previous study, incomplete reprogramming of donor nuclei that were transferred to recipient cytoplasts was suggested to cause aberrant expression of genes associated with the mitochondria; this aberrant expression was associated with developmental arrest and various other disorders in the cloned embryos (Jincho et al., 2008). Although further investigation is necessary for clarifying the mechanism underlying the lower oxygen consumption in the SCNT embryos than in the IVF embryos, incomplete reprogramming of the donor nucleus might have led to mitochondrial dysfunction, resulting in anomalous oxygen consumption in the SCNT embryos at D5BL.
The activation of oxygen consumption at the blastocyst stage was associated with the energy demand for protein synthesis and activity of the plasma membrane Na+-K+-dependent ATPase during blastocoele formation and embryo implantation (Harvey et al., 2002; Nilsson et al., 1982). Hence, the low oxygen consumption in the SCNT embryos at D5BL appears to be involved in various disorders, including low TE cell number (Koo et al., 2002), abnormal placentation (Yang et al., 2007b), and high apoptosis incidence in cells of extra-embryonic tissues such as TE (Chae et al., 2006; Lee et al., 2007). In the present study, we observed that when the IVF embryos actively consumed oxygen at D5BL (>0.81 × 1014/mol sec−1), they exhibited higher hatching and hatched rates at D7BL (Fig. 2). In fact, >50% of the IVF embryos at D5BL exhibited active oxygen consumption along with hatching competence, whereas only 4% of the SCNT embryos at D5BL exhibited active oxygen consumption with a similar level of competence (Fig. 2). Furthermore, it has been suggested that the low level of mitochondrial metabolism causes a decrease in the TE cell number (Thouas et al., 2006), and that low oxygen consumption is associated with the apoptosis incidence (Manser and Houghton, 2006; Trimarchi et al., 2000). Our results also suggest the involvement of low oxygen consumption at D5BL in the low TE cell number and high apoptosis incidence at D7BL (Figs. 3 and 4).
In conclusion, we showed that most SCNT embryos exhibited anomalous oxygen consumption at D5BL and low TE cell number and high apoptosis incidence at D7BL. However, we found that anomalous oxygen consumption at D5BL and limited hatching competence, low TE cell number, and high apoptosis incidence at D7BL in the SCNT embryos could be improved by using cumulus cells as donor nuclei (Tables 3, 4, and 5). These results are consistent with the results of previous studies that reported that in the case of many species, cumulus cells might be the appropriate donor nuclei for SCNT embryos (Kato et al., 2000; Wakayama et al., 1998; Yang et al., 2007a). Thus, although embryo transfer after measurement of oxygen consumption needs to be investigated in the future, our findings suggest that measurement of oxygen consumption by using SECM could be useful as a noninvasive and physiological evaluation method for the developmental competence of porcine SCNT embryos.
Footnotes
Acknowledgments
We thank the staff of the Meat Inspection Office, Sendai City, for supplying porcine ovaries. This study was supported by a grant-in-aid from the Japan Society for the Promotion of Science (JSPS-16108003) to E.S., grants-in-aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan (17380164, 19021006) to H.A., and Research Fellowships for Young Scientists from the Japan Society for the Promotion of Science to S.S.
Author Disclosure Statement
The authors do not have any competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.