
Abstract
Abstract
High rates of embryonic, fetal, or placental abnormalities have consistently been observed in bovine cloning. Segregation of inner cell mass (ICM) and trophectoderm (TE) lineages in early embryos is an important process for fetal and placental formation. In mouse embryos, differentiation of ICM and TE is regulated by various transcription factors, such as OCT-4, CDX2, and TEAD4, but molecular mechanisms that regulate differentiation in bovine embryos remain unknown. To clarify gene transcripts involved in segregation of ICM and TE lineages in bovine embryos, we examined the relative abundances of OCT-4, CDX2, TEAD4, GATA3, NANOG, and FGF4 transcripts in blastocyst embryos derived from in vitro fertilization (IVF). Furthermore, transcript levels of such genes in bovine embryos derived from somatic cell nuclear transfer (NT-SC) and in vivo (Vivo) were also compared. OCT-4, NANOG, and FGF4 transcript levels in IVF embryos were significantly higher in ICM than in TE. In contrast, the CDX2 transcript level was lower in ICM than in TE. In NT-SC embryos at the blastocyst stage, transcript levels of all genes except CDX2 were lower than that in Vivo embryos. In the elongated stage, expression levels of the six genes did not differ between NT-SC and Vivo embryos. We observed aberrant expression patterns of various genes involved in segregation of ICM and TE lineages in bovine NT-SC embryos. These results raise the possibility that abnormalities in the cloned fetus and placenta are related to the aberrant expression of genes involved in segregation and differentiation process in the early developmental stage.
Introduction
In mammalian embryos, the first lineage segregation occurs during the transition from the morula to blastocyst stage where blastomeres differentiate into the ICM or TE lineages. The ICM portion differentiates into the embryonic tissue, which comprises ectoderm, mesoderm, and endoderm, and the TE lineage differentiates into the placental organ after implantation (Pedersen et al., 1986). In mouse embryos, the transcription factors POU class 5 homeobox 1 (OCT-4) and Caudal-related homeobox 2 (CDX2) play pivotal roles in segregation of ICM and TE (Nichols et al., 1998; Niwa et al., 2005; Strumpf et al., 2005). Recently, TEA domain family transcription factor 4 (TEAD4) (Nishioka et al., 2008; Yagi et al., 2007), GATA binding protein 3 (GATA3) (Home et al., 2009), and NANOG (Chen et al., 2009) have also been observed to be important in regulation of CDX2 expression. On the other hand, fibroblast growth factor 4 (FGF4) is required for functional ICM formation, and this factor plays an important role in differentiation of the TE portion into the placental tissue (Feldman et al., 1995; Niswander and Martin, 1992). Segregation of ICM and TE lineages is regulated by various genes that play a key role in the process that is critical to subsequent fetal and placental formation. However, molecular mechanisms that regulate segregation of ICM and TE lineages in bovine embryos are largely unknown.
As described above, gene transcripts involved in segregation of ICM and TE lineages are important factors for embryo development, and may influence the function of NT-SC embryos. Here we tried to clarify gene transcripts involved in segregation of ICM and TE lineages in bovine embryos by examining the relative abundances of several gene transcripts that are well known as principal factors responsible for segregation and maintenance of ICM and TE lineages in mouse embryos. Furthermore, we tried to evaluate the ability of differentiation in bovine NT-SC embryos by comparing expression levels of such genes in these embryos derived from NT-SC and in vivo (Vivo).
Materials and Methods
All experiments used animals were approved by the Animal Ethics Committee, Hokkaido Animal Research Center in Shintoku, Japan. In vitro maturation (IVM) of oocytes, in vitro production of embryos, and analysis of gene transcripts in bovine embryos were essentially performed as described previously (Sawai et al., 2005, 2007, 2010).
In vitro maturation of oocytes
Cow ovaries were collected at a local slaughterhouse and maintained at room temperature during transport to the laboratory. Cumulus–oocyte complexes (COCs) were aspirated from follicles of 2–8 mm. Ten bovine COCs were matured in a 100 μL drop of M-199 medium (Invitrogen, Carlsbad, CA, USA) with 10% fetal calf serum (FCS; Invitrogen) for in vitro fertilization (IVF), or a 100 μL drop of IVMD-101 medium (Research Institute for the Functional Peptides, Yamagata, Japan) (Abe et al., 1999) for NT-SC at 39°C in a humidified atmosphere containing 5% CO2 in air for 22 h.
In vitro fertilization
IVF was performed as described by Niwa and Ohgoda (1988). Briefly, in vitro matured COCs were coincubated with frozen-thawed semen in modified Brackett and Oliphant medium (Brackett and Oliphant, 1975) containing 10 mg/mL of BSA (Sigma-Aldrich, St. Louis, MO, USA), 5 mM of Caffeine (Sigma), and 2.0 IU/mL of heparin (Novo Nordisk A/S, Bagsvard, Denmark) for 6 h at 39°C under 5% CO2 in air. The final sperm concentration added per fertilization drop was 5 × 106 sperm/mL. Following coincubation with sperm, oocytes were freed from cumulus cells.
Nuclear transfer
Bovine fibroblast cells for NT-SC were collected from a skin tissue biopsied from a calf ear using 0.1% collagenase (Sigma) and 0.25% trypsin (Invitrogen). Primary cells were cultured in Dulbecco's modified Eagle medium (DMEM; Invitrogen) with 10% FCS (Roche Diagnostics, Tokyo, Japan) for up to four passages. Prior to NT-SC, fibroblast cells at 50–70% confluence were cultured for a further 4–5 days in serum-depleted medium (DMEM with 0.5% FCS). Following IVM, COCs were freed of their cumulus cells by vortexing in 0.1% hyaluronidase (Sigma) for 5 min. Zona pellucidae of oocytes were cut using a fine glass needle. The oocytes were enucleated by removing the first polar body and metaphase II plate with a small amount of surrounding cytoplasm. A single donor cell was inserted into the perivitelline space of the enucleated oocyte. Recipient oocyte–cell complexes were pulsed with two direct current electric pulses of 20 V/150 μm for 50 μsec with a 1-sec interval and then treated with 10 μg/mL of cycloheximide (Sigma) for 5 h.
In vitro culture of bovine embryos
Following IVF or NT-SC, embryos were cultured in modified TALP (mTALP) medium (Bavister et al., 1983) with 0.1% BSA at 39°C in 5% CO2, 5% O2, and 90% N2. On day 3 (IVF or NT-SC = day 1), embryos were transferred to mTALP supplemented with 3% new-born calf serum (Invitrogen) and subsequently cultured at 39°C in 5% CO2, 5% O2, and 90% N2 until day 8.
Embryo transfer, recovery of elongated embryos and production of in vivo embryos
Transfer of NT-SC embryos and their recovery from recipient cows were performed essentially as described previously (Sawai et al., 2007, 2010). Briefly, recipient cows for embryo transfer were estrus-synchronized by insertion of the CIDR device (EAZI-Breed; InterAg, Hamilton, New Zealand) for 7–14 days and by administration of PGF2α (cloprostenol 0.5 mg, Resipron-C; Teikoku Hormone, Tokyo, Japan) injection 1 day before the removal of the device. A total of 26 NT-SC embryos at the blastocyst stage were nonsurgically transferred to three recipients (8 or 10 embryos per recipient) 6–7 days after estrus. On day 16 (NT-SC = day 1), elongated embryos were recovered nonsurgically by uterine flushing using a Foley catheter (20 French gauge).
Vivo embryos were obtained from donor cows that had been treated for superovulation and artificial insemination (AI) using methods described previously (Sawai et al., 2007, 2010). Briefly, a total of 20 IU of FSH (Antorin; Denka Pharmaceutical, Kanagawa, Japan) was given to the donors twice daily in decreasing doses over 3 days. To induce luteolysis, PGF2α was injected on the third day of superovulation. Donors were bred by AI at 12–24 h after the onset of estrus. On days 8 and 16 (AI = day 1), blastocyst and elongated embryos were nonsurgically recovered from 4 (day 8) or 2 (day 16) donor cows by uterine flushing using the Foley catheter (20 French gauge).
Determination of the relative abundances of gene transcripts in bovine embryos
Under an inverted microscope, blastocyst embryos were divided into the following two parts using the micro surgical blade (Feather, Osaka, Japan): intact ICM with surrounding TE cells and a TE portion (Fig. 1A and B). ICM and TE samples obtained from four to five different blastocyst embryos were pooled to form single samples. ICM and TE portions were washed three times in PBS containing 1% polyvinyl pyrrolidone (PVP; Sigma), added to 5 μL of lysis buffer [0.8% Igepal (ICN Biomedical Inc., Aurora, OH, usa), 5 mM of DTT (Invitrogen), and 1 U/μL of RNAsin (Promega, Madison, WI, USA)), snap frozen in liquid nitrogen, and stored at −80°C. Recovered elongated embryos were transferred to 1% PVP-PBS. Elongated stage embryos in which we could identify both TE and the embryo disc (ED) were used for analysis (Fig. 1C and D). Under a stereomicroscope, elongated embryos were then divided into the following two parts using a surgical knife: TE and ED regions. The TE region was divided into several pieces, approximately 3–5 mm wide. Pieces of TE and ED from single elongated embryos were washed three times in 1% PVP-PBS. Total RNA was isolated from TE and ED using the RNeasy Mini kit (Qiagen, Tokyo, Japan) according to the manufacturer's instructions and stored at −80°C.
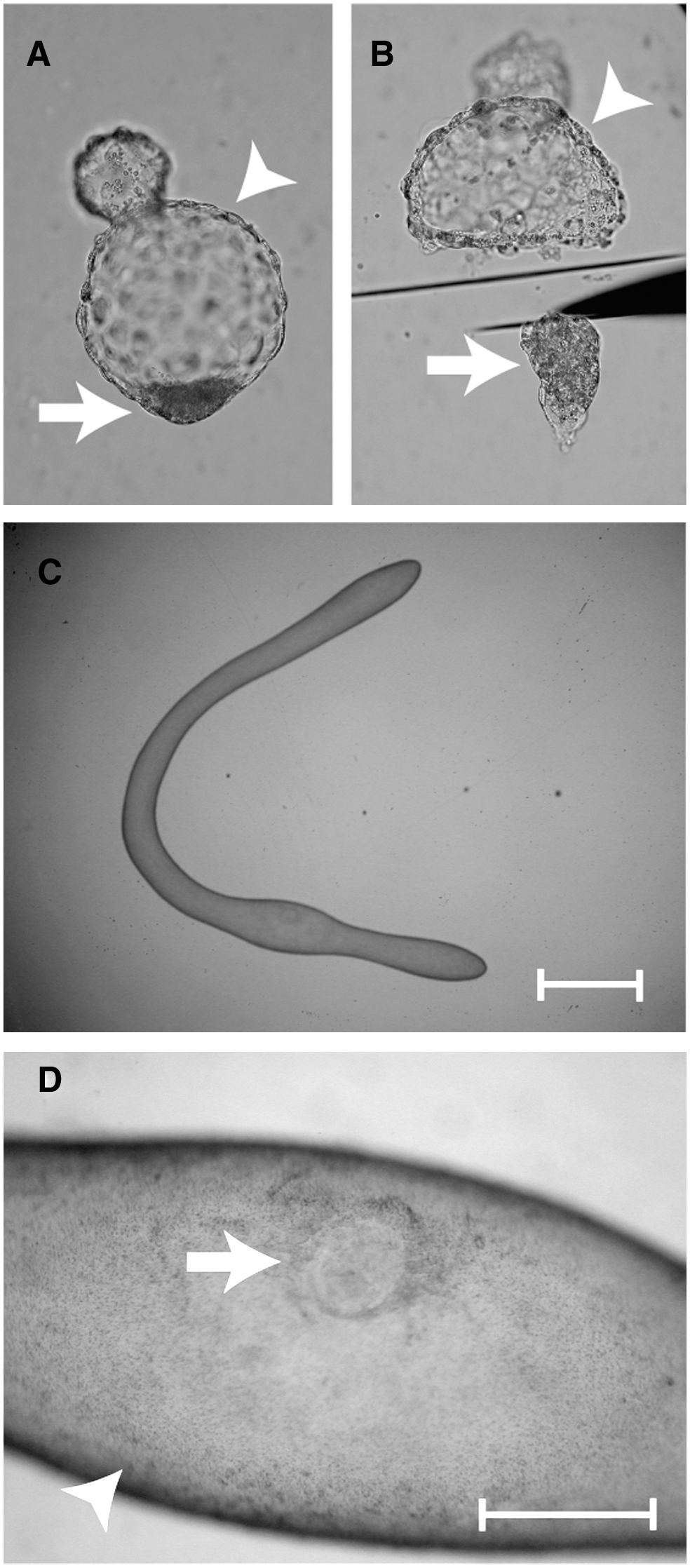
Representative photographs of bovine blastocyst (
RNA samples were heated to 80°C for 5 min and treated for reverse transcription (RT) using the QuantiTect Reverse Transcription kit (Qiagen) according to the manufacturer's instructions. The reaction mixture of blastocyst and elongated stage samples was diluted with DEPC-treated water to obtain a final volume of 33 and 40 μL, respectively. Real-time PCRs were performed using StepOneTM (Applied Biosystems, Tokyo, Japan), and products were detected with SYBR Green, which is included in the QuantiTect SYBR Green PCR master mix (Qiagen). For each quantification, 2 μL of the RT product was used. The amplification program was as follows: preincubation at 95°C for 15 min to activate HotStar Taq DNA polymerase (Qiagen), followed by 45 cycles of denaturation at 94°C for 15 sec, annealing of primers at different temperatures (Table 1) for 30 sec, and elongation at 72°C for 30 sec. After the end of the last cycle, a melting curve was generated by starting fluorescence acquisition at 60°C and taking measurements every 0.3°C until 95°C was reached.
Primer orientations: F, forward; R, reverse.
A standard curve was generated by amplifying serial dilutions of a known quantity of amplicons. PCR products for each gene were purified using the QIAquick PCR purification kit (Qiagen) and quantified by measuring absorbance at 260 nm using Nano Drop (ND-1000, Thermo Fisher Scientific, Kanagawa, Japan) and diluted. Serial 10-fold dilutions for creating a standard curve were amplified in every real-time PCR run. The standards and cDNA samples were then coamplified in the same reaction prepared from a master mix. Fluorescence was acquired at each cycle to determine the threshold cycle or the cycle during the log-linear phase of the reaction at which fluorescence rises above the background for each sample. Final quantification was done using the StepOneTM quantification software, and results were reported as relative expression after normalization of the amount of transcript to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) in the same samples.
Statistical analysis
Differences in mRNA expression levels in IVF embryos were analyzed by the Mann-Whitney U-test. Data on mRNA expression levels in Vivo and NT-SC embryos were analyzed by the Kruskal-Wallis test followed by multiple pair-wise comparisons using the Scheffe method. A p-value <0.05 denoted a statistically significant difference.
Results
Developmental competence of blastocyst and recovery rate of elongated embryos
The blastocyst developmental rates (number of blastocyst/number of one-cell stage embryos in culture) for IVF and NT-SC embryos were 44.7% (71/159) and 22.4% (73/326), respectively. The recovery rate (number of embryos recovered/number of embryos transferred) of elongated embryos derived from NT-SC was 30.8% (8/26).
Gene transcripts in ICM and TE of blastocyst stage embryos derived from IVF
To clarify expression levels of genes involved in segregation of ICM and TE lineages in bovine embryos, mRNA transcripts of OCT-4, CDX2, TEAD4, GATA3, NANOG, and FGF4 in ICM and TE regions of blastocyst stage embryos derived from IVF were examined. The relative abundances of OCT-4, NANOG, and FGF4 transcripts were significantly higher (p < 0.01–0.05) in ICM than in TE (Fig. 2A, E, and F). In contrast, the CDX2 transcript level in the TE portion was significantly higher (p < 0.05) than that in the ICM portion (Fig. 2B). The relative abundances of TEAD4 and GATA3 transcripts did not differ between ICM and TE (Fig. 2C and D).
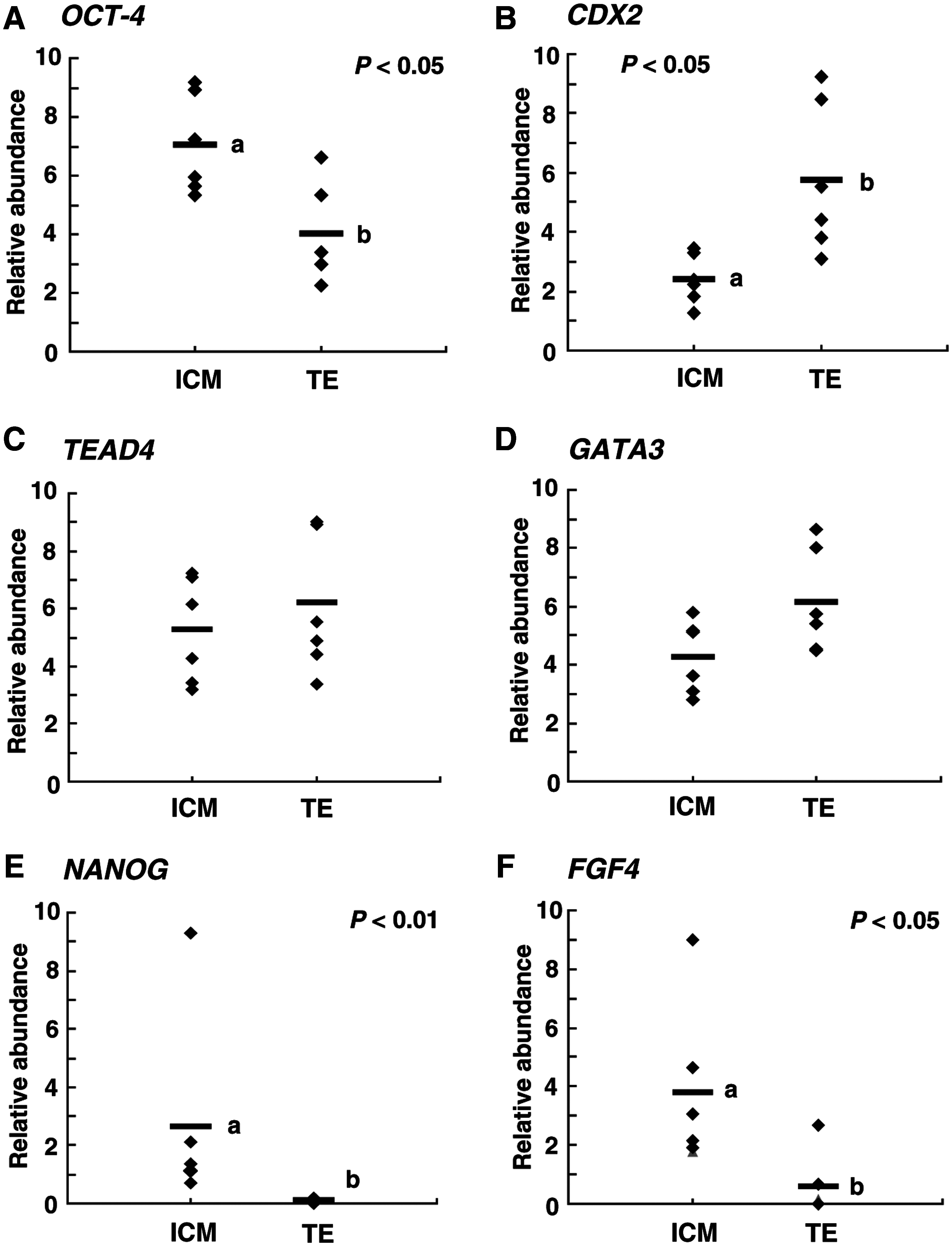
Relative abundances of (
Comparisons of gene expression levels in NT-SC and Vivo embryos
To evaluate expression levels of genes involved in segregation of ICM and TE lineages in bovine NT-SC embryos, OCT-4, CDX2, TEAD4, GATA3, NANOG, and FGF4 transcript levels in NT-SC and Vivo embryos were compared. As shown in Figure 3A, E, and F, in the blastocyst embryos, the relative abundances of OCT-4, NANOG, and FGF4 transcripts in Vivo embryos were significantly higher (p < 0.01–0.05) in ICM than in TE, the same pattern was seen in IVF embryos. However, transcript levels of these genes in NT-SC embryos did not differ between ICM and TE; both ICM and TE portions showed low transcript levels. The CDX2 transcript level did not differ between ICM and TE derived from Vivo and NT-SC embryos (Fig. 3B). The relative abundances of TEAD4 and GATA3 transcripts did not differ significantly between ICM and TE regardless of the origin of the embryos (Fig. 3C and D). However, these transcript levels were lower both in ICM and TE derived from NT-SC than from Vivo embryos.
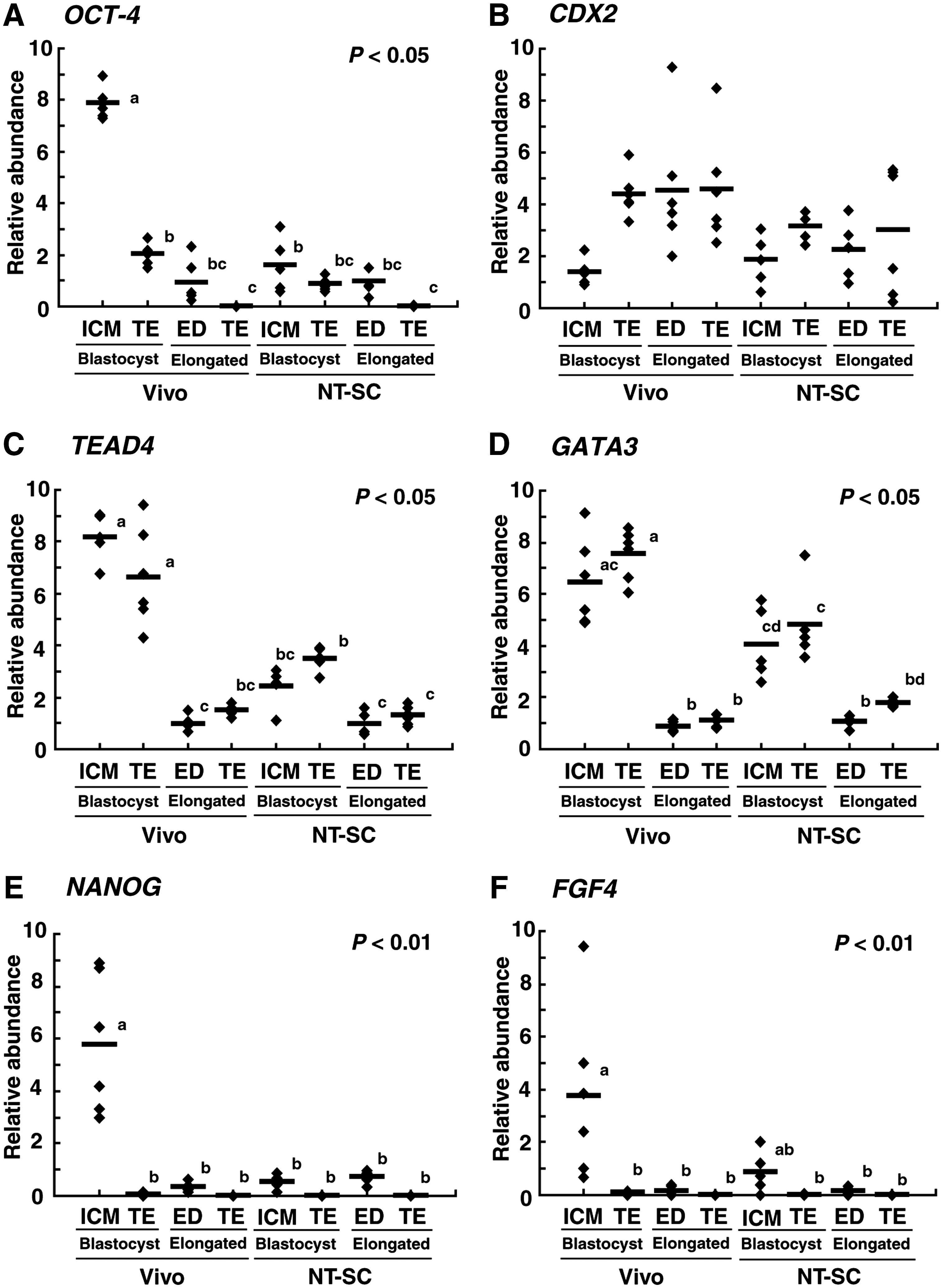
Relative abundances (mean ± SEM) of (
In Vivo embryos, OCT-4, TEAD4, and GATA3 transcript levels were significantly (p < 0.05) reduced from the blastocyst to elongated stage (Fig. 3A, C, and D). Similarly, the relative abundances of NANOG and FGF4 transcripts in Vivo embryos was significantly (p < 0.01) reduced in ED than in ICM (Fig. 3E and F). On the other hand, transcript levels in NT-SC embryos showed two patterns. First, lower transcript levels in both ICM and TE portions at the blastocyst stage, OCT-4, NANOG, and FGF4 transcript levels were sustained at the elongated stage (Fig. 3A, E, and F). Second, the relative abundances of TEAD4 and GATA3 transcripts were significantly (p < 0.05) reduced at the elongated stage (Fig. 3C and D). The CDX2 transcript level did not change between blastocyst and elongated stages regardless of the differences in the origin and tissue of the embryos (Fig. 3B). At the elongated stage, the relative abundances of all gene transcripts did not differ between Vivo and NT-SC embryos.
Discussion
Several studies of mouse embryos have indicated clearly that segregation of ICM and TE lineages is regulated by the mutually antagonistic effects of OCT-4 and CDX2 (Niwa et al., 2005). At the morula stage, OCT-4 and CDX2 are initially coexpressed throughout the embryo, but after initiation of blastocyst formation, OCT-4 and CDX2 expression are restricted to ICM and TE, respectively (Niwa et al., 2005). CDX2 and OCT-4 form a transcription network by which they activate their own transcription while reciprocally suppressing each other's expression. This transcription network contributes to the establishment of the mutually exclusive CDX2 and OCT-4 expression, and thus the establishment of TE and ICM lineages (Niwa et al., 2005). Furthermore, recent investigations have revealed that TEAD4 and GATA3 play important roles in the specification and development of the TE lineage in mouse embryos (Home et al., 2009; Nishioka et al., 2008, 2009; Yagi et al., 2007). TEAD4 expression was observed in both ICM and TE of mouse embryos (Nishioka et al., 2008). In TEAD4-deficient mouse embryos, CDX2 protein was not detected at the blastocyst stage, and showed defects in the specification of the TE lineage (Nishioka et al., 2008; Yagi et al., 2007). These data suggest that TEAD4 regulates TE development through a pathway that includes activation of CDX2 expression (Nishioka et al., 2008). In mouse embryos, GATA3 expression is restricted to TE and directly regulates the CDX2 transcript level at the blastocyst stage (Home et al., 2009). In addition, as similar to CDX2 expression, GATA3 expression depends on the TEAD4 transcript (Ralston et al., 2010). NANOG, which is an ICM specific transcription factor and under the control of OCT-4 (Rodda et al., 2005), and CDX2 could also bind each promoters and thus influence gene expression (Chen et al., 2009). On the other hand, FGF4 expression is restricted to ICM, and produced in the ICM signal to TE, maintaining it in a proliferative state. Thus, FGF4 plays an important role in TE proliferation (Tanaka et al., 1998), and FGF4 transcript is an optimal indicator for evaluating ICM function. In bovine embryos, limited information is available about the factors regulating segregation of ICM and TE lineages. To date, TEAD4 and GATA3 transcripts have not been investigated in bovine blastocyst stage embryos. In the present study, we indicated that six genes including TEAD4 and GATA3 expressed in bovine embryos.
In mouse embryos, expression of abovementioned proteins other than TEAD4 is confined to ICM or TE tissues. OCT-4, NANOG, and FGF4 are expressed in the ICM, and CDX2 and GATA3 are expressed only in the TE (Chambers et al., 2003; Home et al., 2009; Mitsui et al., 2003; Nichols et al., 1998; Nishioka et al., 2008; Niswander and Martin, 1992; Ralston et al., 2010; Yagi et al., 2007). In bovine embryos, it is well known that OCT-4 protein and mRNA are detectable in both ICM and TE (Degrelle et al., 2005; Kirchhof et al., 2000; Kuijk et al., 2008; van Eijk et al., 1999). In the present study, OCT-4 and CDX2 transcripts were detected in both ICM and TE portions, but these transcript levels were higher in ICM and TE, respectively. Furthermore, the OCT-4 transcript level was scarce in TE at the elongated stage and was considerably lower than that at the blastocyst stage. The OCT-4 protein seems to be restricted to the ICM lineage of early elongated bovine embryos (Vejlsted et al., 2005). Degrelle et al. (2005) suggested that the detection of OCT-4 in TE of such blastocyst stage embryos was caused by either untranslated OCT-4 transcripts, which do not disappear quickly in bovine TE tissue or by the presence of tiny amounts of OCT-4 protein. Similarly, Kuijk et al. (2008) reported that CDX2 protein was detected only in the TE portion of bovine blastocyst stage embryos. The CDX2 transcript level in the bovine ICM might be insufficient for protein expression. In contrast, NANOG and FGF4 expression in bovine embryos were restricted to ICM as in mouse embryos.
Recent findings obtained by epigenetic analysis provide evidence that as a consequence of serious errors in reprogramming of the donor nucleus, aberrant expression patterns of various genes are seen in NT-SC embryos. It is generally accepted that fetal and placental development anomalies are caused by unconventional patterns of gene expression in NT-SC embryos. In fact, many investigators have reported that both morphological changes and aberrant gene expression were observed in the cloned fetus and placenta after implantation. In the present study, we focused on gene transcripts involved in segregation of ICM and TE lineages in bovine NT-SC embryos, and on changes in gene transcript levels from the blastocyst to elongated stage when the first step in differentiation has been concluded. In the present study, OCT-4, NANOG, and FGF4 transcript levels in Vivo embryos showed the same patterns as in IVF embryos, and the levels were higher in ICM than in TE. However, transcript levels of these genes in NT-SC embryos were lower in both ICM and TE portions. Similar findings were obtained in mouse NT-SC embryos where OCT-4 transcript is often downregulated (Kishigami et al., 2006). Daniels et al. (2000, 2001) reported aberrant expression of FGF4 mRNA in bovine NT-SC embryos, which is consistent with our observations. In previous reports, most blastocyst stage embryos derived from murine NT-SC expressed CDX2 normally (Kishigami et al., 2006), but bovine NT-SC embryos showed aberrant CDX2 expression (Hall et al., 2005). In the present study, the significantly difference in the CDX2 transcript level between ICM and TE as seen in IVF embryos was not showed in Vivo and NT-SC embryos. However, CDX2 transcript level in TE obtained from Vivo embryos were higher than that in ICM. On the other hand, the difference of CDX2 transcript level between ICM and TE in NT-SC embryos was not clear. TEAD4 and GATA3 transcript levels in IVF and Vivo embryos did not differ between ICM and TE, and therefore, the role of these two genes in segregation of ICM and TE lineages in bovine embryos remains unknown. However, TEAD4 and GATA3 transcript levels were less in elongated stage embryos than in blastocyst stage embryos. Thus, as in mouse embryos, these genes might participate in regulation of transcripts of signaling factors such as OCT-4 and CDX2. Furthermore, TEAD4 and GATA3 transcript levels were lower in both ICM and TE portions of NT-SC embryos than those in Vivo embryos. In the present study, it is difficult to consider that gene expression patterns in NT-SC embryos were due to the in vitro procedures such as IVM or embryo culture condition. If these factors affected gene expression patterns, IVF embryos should have showed the similar patterns in NT-SC embryos. Therefore, our results obtained from analysis of six genes indicate clearly presence of epigenetic abnormalities involved in segregation and differentiation in bovine NT-SC embryos. In the present study, ICM and TE samples were prepared by cutting method using the microsurgical blade. There was no difference of technical obstacle between NT-SC and Vivo embryos at blastocyst stage. However, trace amount of TE portion was contained in ICM samples. Relative abundances of OCT-4, NANOG, and FGF4 were higher in ICM portion obtained from IVF and Vivo than that in TE. In contrast, transcript level of CDX2 was lower in ICM. These results were in agreement with the results of mouse embryos. Thus, we infer that present results were not highly influenced by contamination of TE portion. However, in the future, analysis using whole-mount immune fluorescence is necessary for more strict evaluation of gene transcripts localization in individual embryos.
In the present study, except for CDX2, the relative abundances of all gene transcripts decreased in both ED and TE at the elongated stage. OCT-4 and NANOG are key factors in the maintenance of pluripotency or undifferentiation in several kinds of stem cells such as embryonic stem cells (Chambers et al., 2003; Niwa et al., 2000). Our findings suggest that transcript levels of these genes decreased according to differentiation during the elongation process of bovine embryos. Our results also present a persuasive argument that these genes are involved in segregation of ICM and TE lineages in bovine embryos. In the present study, the CDX2 transcript level was not reduced at the elongated stage. Sakurai et al. (2009) suggested that the CDX2 transcript in bovine TE cell lines regulates interferon tau (IFN-τ), which is an important factor in implantation of elongated stage embryos. Similarly, Gata family genes, including GATA2 and GATA3, expressed in the bovine conceptus at days 17–22 after estrus, are also involved in IFN-τ transcription (Bai et al., 2009). In the present study, GATA3 and TEAD4 transcript levels in the TE portion at the elongated stage did not disappear completely as of OCT-4, NANOG, and FGF4. Therefore, we consider that CDX2, TEAD4, and GATA3 expression are sustained or retained in elongated stage embryos to maintain secondary roles such as secretion of IFN-τ at the elongated stage.
In the present study, expression levels of all genes did not differ between NT-SC and Vivo embryos at the elongated stage. In our previous studies (Sawai et al., 2010), the aberrant methylation level in bovine NT-SC embryos was corrected as a result of demethylation and retention of methylation as the embryo developed and differentiated. Furthermore, the transcript level of some genes, including insulin-like growth factor-related genes in NT-SC embryos that exhibited gene expression abnormality at the blastocyst stage, became similar to levels in control embryos derived from both IVF and Vivo at the elongated stage (Moore et al., 2007; Sawai et al., 2007). Six genes evaluated in this study were downregulated or sustained from the blastocyst to elongated stage, and thus we could not conclude that changes in expression levels in NT-SC embryos were normalized during the elongation process. On the other hand, several investigators (Giraldo et al., 2008; Sawai et al., 2005) reported about variations of gene expression levels in individual NT-SC embryos at blastocyst stage. Therefore, NT-SC embryos, which expressed these genes at normal levels, could possibly grow to the elongated stage in the uterus. However, further analysis of transcript levels in individual embryos at various stages is necessary to demonstrate our hypothesis.
In the present study, we have demonstrated aberrant expression patterns of various genes involved in segregation and maintenance of ICM and TE lineages in bovine NT-SC embryos. Some investigators have reported aberrant allocations of ICM and TE cells in bovine NT-SC embryos at the blastocyst stage (Koo et al., 2002; Li et al., 2006b). We provide additional epigenetic characters of ICM and TE tissues derived from NT-SC, and this finding presents the possibility that abnormalities of the cloned fetus and placenta are caused by the epigenetic errors at the first step in embryo differentiation.
Footnotes
Acknowledgments
This work was supported by a Grant-in-Aid for Scientific Research on Priority Areas (No.21028001).
Author Disclosure Statement
The authors declare that no conflicting financial intersts exist.