
Abstract
Abstract
We examined the transduction efficiency in human embryonic stem (ES) and induced pluripotent stem (iPS) cells using an adenovirus (Ad) vector. RT-PCR analysis revealed the expression of the coxsackievirus and adenovirus receptor, a receptor for Ad, in these cells. However, gene expression after the transduction with an Ad vector was observed only in the periphery of ES and iPS cell colonies, when human ES and iPS cells were passaged as small colonies. This suggests that the Ad vector could not enter inside the ES and iPS cell colonies by their tight connection. We thus attempted to transduce foreign genes into the dissociated form of human ES and iPS cells, which were passaged using Rho-associated kinase inhibitor. In this condition, transduction efficiency in human ES and iPS cells was markedly increased and transgene expression was observed even inside the colonies by using Ad vectors. Furthermore, Ad vector-mediated transduction did not alter the expression of undifferentiated markers such as Oct-3/4, Nanog, and SSEA-4. Our results indicate that Ad vectors are effective tools for transduction into human ES and iPS cells.
Introduction
The development of an efficient gene transfer system in human ES and iPS cells is important to differentiate these pluripotent cells into lineage-committed cells and to analyze gene function. So far, many researchers have utilized the constitutive transgene expression system by establishing of antibiotic-resistant human ES cell lines using plasmid-based stable transfection methods (Eiges et al., 2001; Vallier et al., 2004) or lentivirus vector systems (Ben-Dor et al., 2006; Kim et al., 2007; Ma et al., 2003). However, a stable transduction system might have unexpected effects, such as overexpression or inactivation of unrelated genes, due to the plasmid DNA or lentivirus genomes being integrated into the host genomes.
Recombinant adenovirus (Ad) vectors have been widely used to deliver foreign genes to cells and tissues because they can achieve high transduction efficiency in both dividing and nondividing cells. Importantly, in an Ad vector system, transgenes are not integrated in the host genomes and transgene expression is transient. Ad vector transduction is thus thought to be appropriate for regulating cellular differentiation. However, few studies have examined Ad vector transduction efficiency for human ES and iPS cells using Ad vectors (Brokhman et al., 2009; Rufaihah et al., 2007; Smith-Arica et al., 2003), possibly because the cytomegalovirus (CMV) promoter is mostly used in the Ad vector and the Ad vector containing the CMV promoter is inactive in human ES and iPS cells, as shown in the present study. We and other groups have shown that the CMV promoter did not work well in some immature cells (Chung et al., 2002; Kawabata et al., 2005; Sakurai et al., 2005; Tashiro et al., 2008, 2009a). Recently, we demonstrated that Ad vectors containing the CMV enhancer/β-actin promoter with β-actin intron (CA) promoter or the human elongation factor (EF)-1α promoter, but not the CMV promoter, efficiently transduce foreign genes into mouse ES and iPS cells (Kawabata et al., 2005; Tashiro et al., 2009a).
In this study, we investigated the transduction efficiency in human ES and iPS cells using Ad vectors containing various types of promoters. The results showed that the Ad vector containing the EF-1α promoter could drive strong transgene expression in both human ES and iPS cells, without changing their undifferentiated states.
Materials and Methods
Ad vectors
Escherichia coli β-galactosidase (LacZ)-expressing Ad vectors (Ad-RSV-LacZ, Ad-CMV-LacZ, Ad-CA-LacZ, and Ad-EF-LacZ), mCherry-expressing Ad vector (Ad-EF-mCherry), and transgene-deficient Ad vector (Ad-null) were constructed previously (Kawabata et al., 2005; Sakurai et al., 2008; Tashiro et al., 2009a). The rous sarcoma virus (RSV) promoter, the CMV promoter, the CA promoter, (a kindly gift from J. Miyazaki, Osaka University, Osaka, Japan) (Niwa et al., 1991), or the EF-1α promoter-driven LacZ gene was inserted into the E1 deletion region of the Ad genome. Ad vectors were prepared and purified by CsCl2 step gradient ultracentrifugation followed by CsCl2 linear gradient ultracentrifugation. The vector particle (VP) titer and biological titer were determined by a spectrophotometric method (Maizel et al., 1968) and an Adeno-X Rapid Titer Kit (BD Clontech, Mountain View, CA, USA), respectively. The ratios of the biological-to-particle titer were 1:41, 1:21, 1:14, 1:22, 1:28, and 1:11 for Ad-RSV-LacZ, Ad-CMV-LacZ, Ad-CA-LacZ, Ad-EF-LacZ, Ad-EF-mCherry, and Ad-null, respectively.
Cells
One human ES cell line, KhES-1, and three human iPS cell lines, 201B2, 201B7, and 253G1, were used in this study. KhES-1 (Suemori et al., 2006), was obtained from the Institute for Frontier Medical Science, Kyoto University (Kyoto, Japan), and KhES-1 was used following the Guidelines for Derivation and Utilization of Human Embryonic Stem Cells of the Ministry of Education, Culture, Sports, Science and Technology, Japan, after approval by the review board. The human iPS cell lines, 201B2, 201B7, and 253G1, were kindly gifted from Dr. S. Yamanaka (Kyoto University, Kyoto, Japan) (Nakagawa et al., 2008; Takahashi et al., 2007). 201B2 and 201B7 were generated from human dermal fibroblasts (HDF) by transducing four factors (Oct-3/4, Sox2, c-Myc, and Klf4) (Takahashi et al., 2007), while 253G1 was generated from HDF by transducing three factors (Oc-3/4, Sox2, and Klf4) (Nakagawa et al., 2008). Human ES and iPS cells were maintained in culture medium [Dulbecco's modified Eagle's medium (DMEM)/F12 (Sigma, St. Louis, MO, USA) supplemented with 20% Knockout Serum Replacement (Invitrogen, Carlsbad, CA, USA), 2 mM L-glutamine, 0.1 mM nonessential amino acids, 0.1 mM 2-mercaptoethanol, and 5 ng/mL recombinant human basic fibroblast growth factor (R&D systems, Minneapolis, MN, USA)] on the mitomycin C-treated mouse embryonic fibroblasts (MEF). For passage, human ES and iPS cell colonies were dissociated into small clumps by the use of 0.1 mg/mL dispase (Roche Diagnostics, Burgess Hill, UK). After centrifugation, the cells were resuspended in culture medium and plated into a T25 flask on feeder cells. The medium was changed daily, and passage was performed every 5 to 6 days. The 293 cells (a human embryonic kidney cell line) and SK HEP-1 cells (a human hepatoma cell line) were cultured in DMEM supplemented with 10% fetal bovine serum and antibiotics.
Ad vector-mediated LacZ transduction
Human ES and iPS cells were plated on 12-well plates. The next day, they were transduced with each Ad vector (Ad-null, Ad-RSV-LacZ, Ad-CMV-LacZ, Ad-CA-LacZ, or Ad-EF-LacZ) at 3000 VP/cell for 1.5 h. After culturing for indicated days, X-gal staining was carried out as described previously (Kawabata et al., 2005). For single cell transduction, human ES and iPS cells were treated with Y-27632 (Wako, Osaka, Japan), a Rho-associated kinase (ROCK) inhibitor, at 10 μM for 1 h before cells were detached from the feeder layer (Watanabe et al., 2007). Human ES and iPS cells were dissociated by pipeting using Pipetman P-1000, and were then passed through the Cell Strainer (BD Biosciences, San Jose, CA, USA). The dissociated cells were seeded on the MEF layer in 12-well plates. On the following day, they were transduced with Ad vectors as described above.
Immunofluorescence staining
Cells were fixed with 4% paraformaldehyde/phosphate-buffered saline (PBS) for 15 min, permeabilized with 0.2% Triton X-100/PBS for 5 min, and blocked with 2% bovine serum albumin/PBS for 30 min. Cells were then stained with appropriate primary antibodies and AlexaFluor-conjugated secondary antibodies (Invitrogen). The primary antibodies for Oct-3/4 (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), Nanog (ReproCELL, Tokyo, Japan), and stage specific embryonic antigen (SSEA)-4 (Chemicon, Temecula, CA, USA) were used in the staining.
RNA extraction and RT-PCR
Total RNA was isolated using ISOGENE (Nippon Gene, Tokyo, Japan) or the RNeasy Kit (Qiagen, Valencia, CA, USA) according to the manufacturer's instructions. cDNA was synthesized by using the SuperScript II reverse transcriptase (Invitrogen) and the oligo(dT) primer. Semiquantitative RT-PCR was carried out by using TaKaRa ExTaq HS DNA polymerase (Takara, Shiga, Japan). PCR products were visualized by ethidium bromide staining after being separated on 2% agarose gel. The sequences of the primers used for semiquantitative RT-PCR were as follows: CAR(F), 5′-GCCTTCAGGTGCGAGATGTTAC-3′; CAR(R), 5′-TCGCACCCATTCGACTTAGA-3′; Oct-3/4(F), 5′-GAGCAAAACCCGGAGGAGT-3′; Oct-3/4(R), 5′-TTCTCTTTCGGGCCTGCAC-3′; Nanog(F), 5′-TTCCTTCCTCCATGGATCTG-3′; Nanog(R), 5′-CTGGGGTAGGTAGGTGCTGA-3′; GAPDH(F), 5′-ACCACAGTCCATGCCATCAC-3′; GAPDH(R), 5′-TCCACCACCCTGTTGCTGTA-3′. The expression level of CAR mRNA was also quantified with the SYBR Premix Ex Taq (TaKaRa), and normalized to that of GAPDH. The sequences of the primers for real time quantitative PCR were as follows: CAR(F), 5′-CAGAAGCTACATCGGCAGTAATCA-3′; CAR(R), 5′-CTCTGAGGAGTGCGTTCAAAGTC-3′; GAPDH(F), 5′-GGTGGTCTCCTCTGACTTCAACA-3′; GAPDH(R), 5′-GTGGTCGTTGAGGGCAATG-3′.
Results and Discussion
Gene transfer using a conventional Ad vector depends on the expression levels of coxsackievirus and adenovirus receptor (CAR), a primary receptor for Ad, on the cellular surface (Bergelson et al., 1997; Tomko et al., 1997). We thus initially examined CAR expression in human ES and iPS cells by semiquantitative RT-PCR. As shown in Figure 1a, we found that Oct3/4- and Nanog-expressing undifferentiated human ES and iPS cells expressed CAR, and that the expression level of CAR was similar between ES and iPS cells. Notably, the expression levels of CAR mRNA in these pluripotent cells were higher than those in the 293 cells and SK-HEP1 cells, both of which were easily infected with conventional Ad vectors (Fig. 1b). These results indicate that conventional Ad vectors should be sufficient to transduce a foreign gene into human ES and iPS cells.
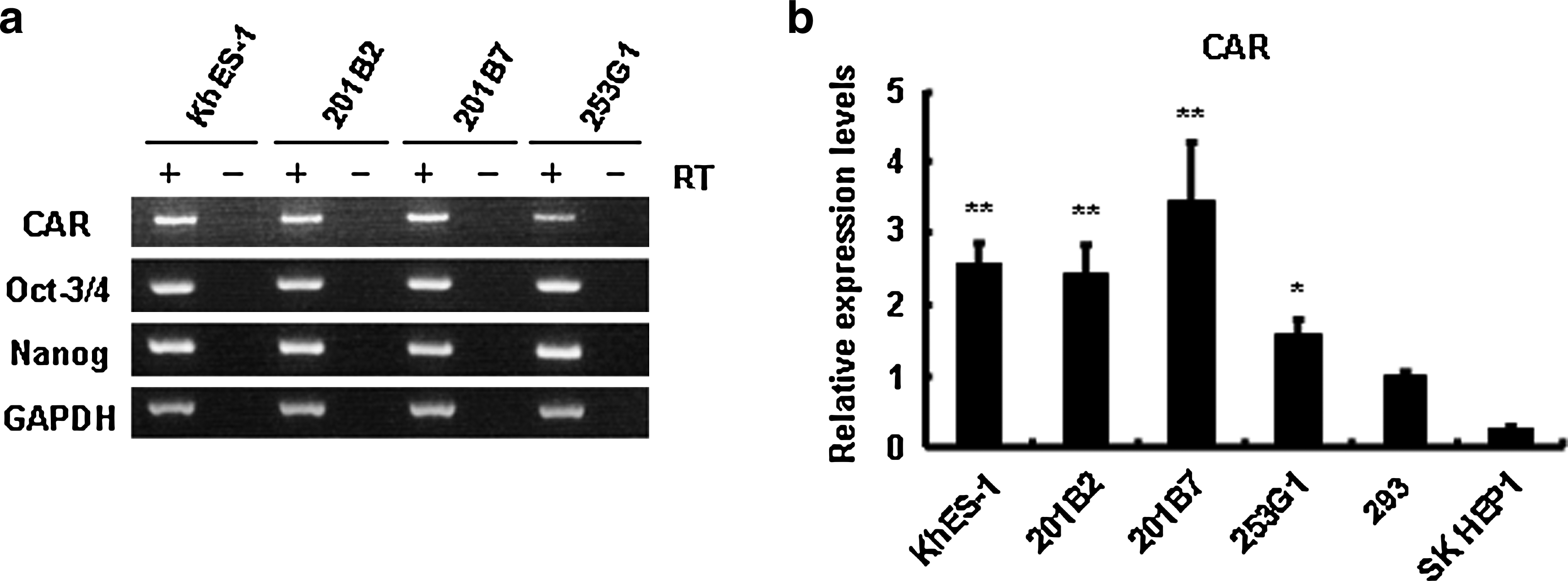
CAR expression in human ES and iPS cells. Total RNA was isolated from human ES cells (KhES-1), iPS cells (201B2, 201B7, 253G1), 293 cells, and SK HEP1 cells, and (
We have demonstrated that the choice of promoter was important for efficient transduction into immature cells such as mouse ES cells, mouse iPS cells, and human hematopoietic stem/progenitor cells (Sakurai et al., 2005; Tashiro et al., 2009a, 2009b). Thus, we next investigated the transduction efficiency in human ES and iPS cells by using LacZ-expressing Ad vectors containing the RSV promoter, the CMV promoter, the CMV enhancer/β-actin promoter with β-actin intron (CA) promoter, or the human elongation factor-1α (EF-1α) promoter (Ad-RSV-LacZ, Ad-CMV-LacZ, Ad-CA-LacZ, or Ad-EF-LacZ, respectively). A transgene-deficient Ad vector, Ad-null, was used as a control vector. Human ES and iPS cells were transduced with each LacZ-expressing Ad vector, and LacZ expression in the cells was evaluated by X-gal staining (Fig. 2a). The CA and the EF-1α promoters had potent LacZ expression not only in human ES cells but also in iPS cells in comparison with the RSV and the CMV promoters. These results are in agreement with our previous data using mouse ES and mouse iPS cells (Kawabata et al., 2005; Tashiro et al., 2009a). However, unlike the case with mouse ES and iPS cells, LacZ expression was observed only in the periphery of human ES and iPS cell colonies, even though the CA or the EF-1α promoter was used. This result suggested that Ad vector could not bind the CAR in human ES and iPS cell colonies although the CAR was highly expressed in human pluripotent cells (Fig. 1). Therefore, we examined whether fiber-modified Ad vectors could improve the transduction into human ES and iPS cell colonies. Fiber-modified Ad vectors could efficiently transduce an exogenous gene into target cells that express quite low levels of CAR (Koizumi et al., 2003; Kurachi et al., 2007; Mizuguchi et al., 2001). We prepared AdRGD-CA-LacZ, AdK7-CA-LacZ, and AdTAT-CA-LacZ, which contain an Arg-Gly-Asp (RGD) peptide, a polylysin (KKKKKKK; K7) peptide, and a transactivator of transcription (TAT; GRKKRRQRRRPQ) peptide, respectively, on the fiber knob of the Ad vector (Koizumi et al., 2003; Kurachi et al., 2007; Mizuguchi et al., 2001). Human ES and iPS cells were transduced with three types of fiber-modified Ad vectors, and the transduction efficiency was estimated by X-gal staining. However, as was the case with conventional Ad vector, the cells in the periphery of ES or iPS cell colonies efficiently expressed LacZ, but little LacZ expression was observed inside the ES and iPS cell colonies (data not shown). Although it is unclear why transgenes were not expressed inside ES and iPS cell colonies, it is possible that the Ad vector could not enter inside these colonies by their tight connection. Thus, we prepared the dissociate form of human ES and iPS cells using Rho-associated kinase (ROCK) inhibitor Y-27632, which is shown to decrease dissociation-induced apoptosis in human ES cells (Watanabe et al., 2007). After dissociated human ES and iPS cells were passaged in the presence of Y-27632, the cells were transduced with LacZ-expressing conventional Ad vectors. In this condition, LacZ activity in the cells transduced with Ad-CA-LacZ or Ad-EF-LacZ was markedly increased and was observed inside ES and iPS cell colonies (Fig. 2b). LacZ was also efficiently expressed in ES and iPS cell colonies when dissociated form of ES and iPS cells was transduced with fiber-modified Ad vectors (data not shown). However, the LacZ expression levels in the cells transduced with fiber-modified Ad vectors were not different from those in the cells transduced with conventional Ad vectors. We thus concluded that conventional Ad vector was sufficient for transduction in human pluripotent stem cells after dissociation with Y-27632. Notably, the EF-1α promoter was more effective than the CA promoter for transducing LacZ genes into human ES and iPS cell colonies. Consistent with our data, other groups have shown that, in lentivirus vector systems, the EF-1α promoter could robustly drive the transgene expression in human ES cells (Kim et al., 2007; Ma et al., 2003), indicating that the EF-1α promoter has strong transduction ability in human pluripotent cells. Taken together, in combination with Y-27632, human ES and iPS cells could be transduced by conventional Ad vectors containing the EF-1α promoter.
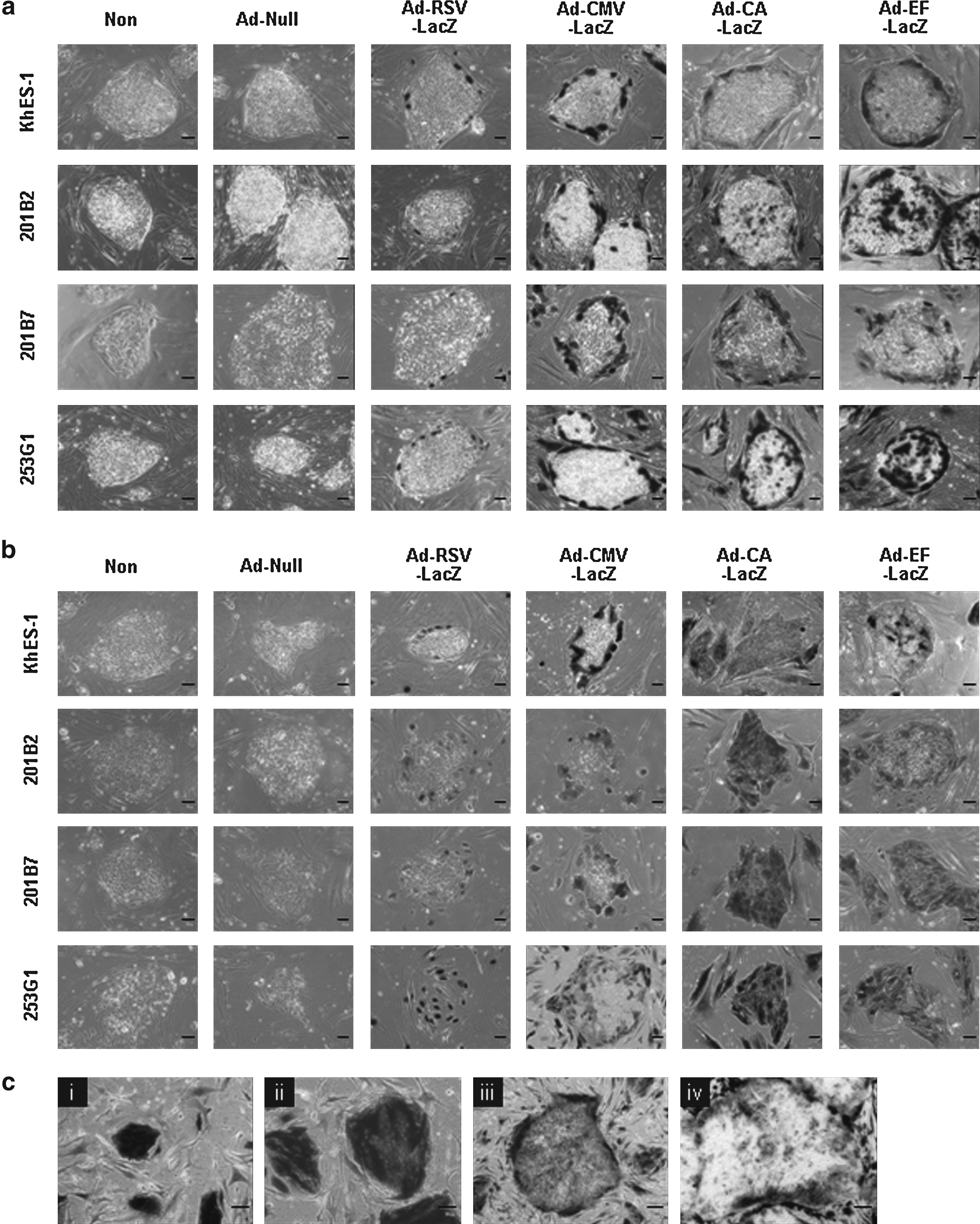
LacZ expression in human ES and iPS cells transduced with Ad vectors containing various types of promoters in the absence or presence of ROCK inhibitor, Y-27632. Human ES cells (KhES-1), and iPS cells (201B2, 201B7, and 253G1) were passaged into culture plates in the absence (
Next, we investigated the time course of LacZ expression in human iPS cells transduced with Ad-EF-LacZ. Dissociated form of human iPS cells (201B7) were cultured for 1, 2, 4, and 6 days after transduction, and LacZ expression was then monitored. Although transgene expression in the cells was continued on day 4 after transduction, LacZ expressing-human iPS cells were decreased on day 6 due to their cell division (Fig. 2c). These data show that Ad vector mediates the transient transgene expression in human pluripotent cells.
We examined whether or not Ad vector-mediated transduction alters the expression of undifferentiated markers in ES and iPS cells. After dissociated human ES and iPS cells were transduced with Ad-EF-mCherry at 3000 VP/cell, the expression of mCherry, Oct-3/4, Nanog, and SSEA-4 was observed by fluorescent microscopy. The results showed that approximately 70–80% of human ES and iPS cells expressed mCherry, and that there was no difference in the expression of undifferentiated markers between nontransduced cells and Ad-EF-mCherry-transduced cells (Fig. 3). This suggests that Ad vector transduction did not change the undifferentiated state of human ES and iPS cells. Therefore, our data demonstrated that the Ad vector containing the EF-1α promoter could efficiently transduce exogenous genes into human ES and iPS cells without decreasing the expression of undifferentiated marker genes.
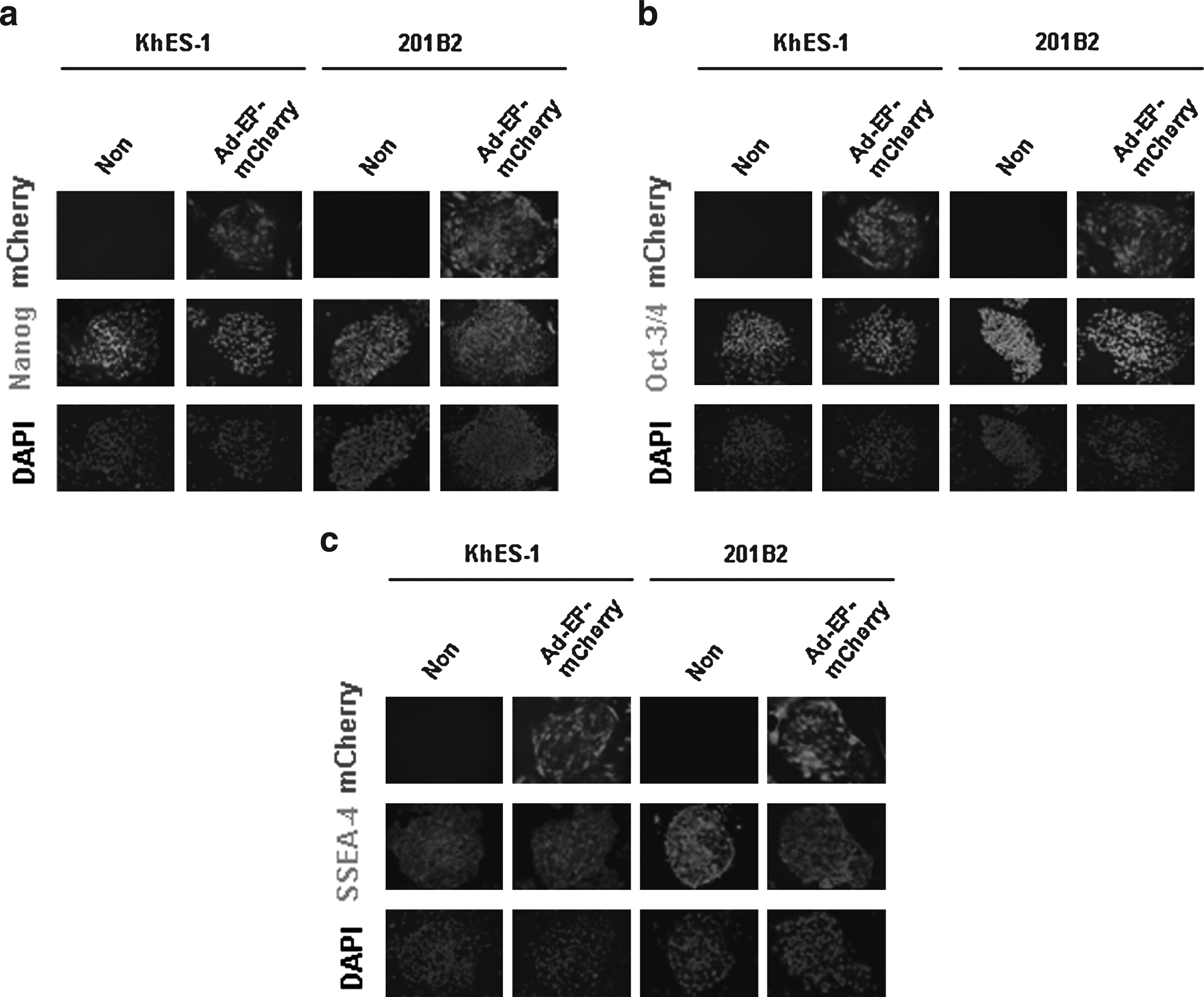
The expression of undifferentiated markers in human ES and iPS cells after the transduction with Ad-EF-mCherry. Human ES cells (KhES-1) and iPS cells (201B2) were plated into culture plates using Y-27632. On the following day, they were transduced with Ad-EF-mCherry at 3000 VP/cell for 1.5 h. Two days later, the expression of Nanog (
Several groups previously reported that 11–80% of human ES cells could express transgenes by conventional Ad vectors (Brokhman et al., 2009; Rufaihah et al., 2007; Smith-Arica et al., 2003). However, they did not optimize the Ad vectors for transduction in human ES cells. They examined the transduction efficiency using only a single Ad vector, such as an Ad vector containing the RSV promoter or the CMV promoter. Human ES cells were transduced with Ad vectors at extremely high titers (500 or 5 × 104 infectious units (ifu)/cell) in their condition (Brokhman et al., 2009; Rufaihah et al., 2007; Smith-Arica et al., 2003). On the other hand, we investigated the transduction efficiency by using various types of Ad vectors including fiber-modified ones, and optimized the Ad vectors for efficient transduction into human ES and iPS cells. This is the first study to report detailed transduction characteristics in human ES and iPS cells with various types of Ad vectors. Our results showed that, in combination with ROCK inhibitor Y-27362, human ES and iPS cells were transducible by a conventional Ad vector containing the EF-1α promoter, and that approximately 70–80% of the ES and iPS cells expressed transgenes by an optimized Ad vector at only 107–136 ifu/cell (this titer is equivalent to 3000 VP/cell). Therefore, the results of the present study suggest that an Ad vector containing the EF-1α promoter is the most suitable vector for efficiently transducing an exogenous gene in human ES and iPS cells.
In summary, we successfully developed the methods to efficiently transduce human ES and iPS cells using Ad vectors. We have demonstrated the usefulness of Ad vector transduction in the field of stem cell differentiation (Tashiro et al., 2008, 2009a, 2009b). Therefore, our transduction system could be a valuable tool to promote the cellular differentiation of human ES and iPS cells into functional cells, such as hematopoietic cells, osteoblasts, hepatocytes, and so on, when an appropriate transcription factor is expressed with Ad vectors.
Footnotes
Acknowledgments
We thank Dr. S. Yamanaka and Dr. J. Miyazaki for kindly providing the human iPS cell lines (201B2, 201B7, and 253G1) and the CA promoter, respectively. We also thank Hiroko Matsumura and Misae Nishijima for their technical assistance. This work was supported by grants from the Ministry of Health, Labor, and Welfare of Japan. K. T. is the Research Fellow of the Japan Society for the Promotion of Science.
Author Disclosure Statement
The authors have no financial conflicts of interest.