
Abstract
Abstract
This study was conducted to determine the effect of microinjection of a single blastomere from in vitro fertilization (IVF)-derived eight-cell embryo into eight-cell cloned embryos harboring the gene encoding recombinant human lactoferrin (rhLF), GFP, and NEO markers in bovine. The reconstructed chimeric embryos were assessed for their development to blastocyst, or to term after transfer, and tissues of offspring were evaluated by polymerase chain reaction (PCR) for the presence of nuclear transfer (NT)-derived transgenic cells, and the cloned embryos without microinjection were used as controls. The chimeric embryos showed slightly higher blastocyst rate than that for controls. The single IVF-derived blastomere appeared to preferential contribute to inner cell mass (ICM) in the chiemric blastocysts. After transfer, the rates of development of chimeric embryos to day 60, to term, and to weaning were significantly higher than those of controls. Sixty-three chimeric blastocysts were transferred and 11 calves were born: 7 calves of them were dead, and the remaining 4 calves are apparently normal and healthy. Most of the tissues collected from dead fetus were transgenic, whereas NT-derived transgenic cells were not detected in some tissues of the living calves. Our results indicated that a single blastomere from IVF-derived eight-cell embryo improves the in vivo developmental potential of transgenic cloned eight-cell embryos in bovine; however, the single IVF-derived blastomere appeares to be better able to populate the ICM and many tissues of offspring than NT-derived blastomeres.
Introduction
An increasing number of studies suggest that epigenetic reprogramming defects, including aberrant DNA methylation or histone acetylation patterns and dysregulation of a large number of imprinted and nonimprinted genes, give rise to the abnormal cloned animals (Cho et al., 2007; Dean et al., 2001; Niemann et al., 2008; Shi and Haaf, 2002). However, the factors that influence the embryos derived from SCNT can develop to full term are still largely unknown. Therefore, it is urgent to devise approaches to increase the frequency of cloning success. Efforts have been made to improve cloning efficiency—such as the selection of donor cells with different cell cycles, origins, and passages (Gibbons et al., 2002; Kasinathan et al., 2001; Yang et al., 2007), changing the methylation or acetylation status of donor cells (Enright et al., 2005; Yang et al., 2007), using zona-free serial nuclear transfer (Hall et al., 2006) and autologous SCNT (Yang et al., 2006), but the effects of these strategies have not been obvious.
In mouse, clone–clone aggregation at the two-cell stage increases the term rate by eightfold (Boiani et al., 2003). In cattle, aggregation of cloned embryos stimulates gene expression and in vitro embryo development (Zhou et al., 2008); however, no improvements in term rate and calf survival rate were noted (Misica-Turner et al., 2007). It was reported that parthenogenetic embryos of mammals die during embryogenesis, but the chimeric embryos combined with parthenogenetic embryos and in vivo fertilized embryos (Clarke et al., 1988; Nagy et al., 1989) or IVF-derived blastomeres (Boediono et al., 1999) can develop normally and further give rise to offspring that live to adulthood, which demonstrates that fertilized embryos or IVF-derived blastomeres can rescue the development of parthenogenetic embryos by way of complementation. Furthermore, Wells and Powell (2000) found a strong bias for NT blastomeres to populate the inner cell mass (ICM) when aggregated with IVF embryos at the eight-cell stage, but no subsequent transfer results were reported. Thus, constructing an embryo with trophectoderm originating from IVF and a nonchimeric ICM originating from NT embryos is possible.
Based on the above presumption, microinjection of an IVF-derived blastomere into a cloned embryo may provide a new method to correct the developmental deficiency of the cloned embryo through complementation, resulting in a more competent embryo. Our aim was to (1) compare the cellular effects, term rates, and calf survival rates of cloned versus chimeric embryos and determine whether an IVF-derived blastomere at the eight-cell stage can improve the developmental potential of cloned bovine embryos, and (2) compare the competency of IVF-derived and NT-derived cells in the chimeric blastocysts and offspring by detecting NT-derived transgenic cells in tissues of the offspring.
Materials and Methods
Materials
Chemicals were purchased from Sigma Chemical Company (St. Louis, MO), and all embryo manipulations were carried out at 38.5°C unless stated otherwise. All experimental parameters were kept the same, for example, the pool of donor embryos and oocytes, activation method, and culture medium.
In vitro maturation of oocytes
Ovaries were collected from a local slaughterhouse and transported to the laboratory in a thermos filled with physiological saline at 25–35°C; these ovaries were derived from Holstein and Yellow cattle. Cumulus–oocyte complexes were harvested from follicles (3–8 mm in diameter) and washed twice in maturation medium (TCM-199 (Gibco, Rockville, MD) supplemented with 10% FBS (HyClone, Logan, UT), 0.01 U/mL FSH, 0.01 U/mL LH, 1 mg/mL estradiol-17b, and 1% (v/v) penicillin/streptomycin). Approximately 50–60 cumulus–oocyte complexes were transferred into 0.5-mL maturation medium in four-well dishes overlaid with paraffin oil, and cultured at 38.5°C, 5% CO2 for 18–20 h. After maturation, cumulus cells were completely removed by vortexing in 0.1% (w/v) hyaluronidase for 2–3 min.
Nuclear donor cell culture
Fetal fibroblasts from the skin of live healthy transgenic cloned cattle 040825 (about 16 months of age, female) and 050211 (about 8 months of age, female) containing the genes encording rhLF,GFP and Neo (Yang et al., 2008) were isolated and cultured as previously described (Gong et al., 2004). Briefly, tissue biopsies were washed, cut into small pieces, and then cultured in 25-cm2 tissue culture flasks. When cells reached confluency after 6–7 days, they were passaged and frozen. Fetal fibroblasts were used as nuclear donors at passage 5–10. Immediately before SCNT, cells were trypsinized, centrifuged, and resuspended in Dulbecco PBS supplemented with 10% (v/v) fetal bovine serum (FBS).
Preparation of eight-cell NT embryos
NT embryos were produced as previously described (Gong et al., 2004). Briefly, maturated oocytes were enucleated by using a beveled glass pipette of 20-μm diameter. Donor cells were then placed between the zona pellucida and the cytoplast membrane. Fusion was performed in a chamber overlaid with fusion buffer [0.3 mmol/L mannitol, 0.15 mmol/L CaCl2, 0.15 mmol/L MgCl2, 0.5 mmol/L HEPES, 0.05% bovine serum albumin (BSA)]. Once the reconstructed embryos were manually aligned, a DC pulse of 2.5 kV/cm for 10 μsec each, with the interval of 1 sec was applied to stimulate oocytes. After the electrical stimulus, the embryos were washed several times and incubated in M199 medium supplemented with 10% FBS for 30 min, and then examined for fusion by light microscopy.
Activation was induced by incubation in 5 μmol/L ionomycin in CR1aa medium for 4 min at 37°C. Embryos were then washed in CR1aa for 5 min before culture in 1.9 mmol/L 6-dimethylaminopurine in CR1aa for 4 h. Embryo culture was performed in CRlaa supplemented with 5% FBS at 38.5°C and 5% CO2. On day 2.5 and day 7.5 after fusion, the number of embryos that developed to the cleavage and blastocyst stages, respectively, were recorded.
Preparation of eight-cell IVF embryos
Twenty-two hours postmaturation, oocytes were placed in fertilization medium [Brackett-Oliphant (B-O) medium supplemented with 10 mg/mL BSA]. Frozen sperm cells (from a fertile Holstein bovine) were thawed, and washed with B-O medium supplemented with 10 mmol/L caffeine, 50 μg/mL heparin, and 1% BSA. Fertilization was conducted for 8 h in droplets of 100 μL B-O medium supplemented with 5 mM caffeine and 1% BSA. The fertilized oocytes were washed twice with M199 and then cultured in CR1aa supplemented with 10% FBS. On day 3, the embryos developed into the eight-cell stage. For GFP+-IVF embryos, frozen sperm from GFP transgenic male cattle was used.
Production of chimeric embryos
Seventy-two hours after SCNT, cloned embryos were placed in manipulation medium [phosphate-buffered saline (PBS) plus 20% FBS]. Using a beveled pipette of 20-μm diameter, only one IVF-derived blastomere (eight-cell stage) was introduced into the cloned embryos. Embryos were then washed twice with M199 and placed in a 500-μL culture drop containing CR1aa supplemented with 10% FBS.
Embryo transfer, pregnancy monitoring, and parturition
Morphological grade 1 to 2 cloned and chimeric blastocysts were selected for embryo transfer and embryo transfer was performed in every April to May of 2006 and 2007. Two to three blastocysts were transferred to each synchronous recipient at 7 days after standing estrus. Using ultrasonography, the pregnancy status of recipient cows was determined on day 60 of gestation. The pregnant cows were allowed to calve naturally if possible or with manual assistance when necessary. Calves were weaned when they weighed more than 100 kg (at about 3 months of age).
Differential staining and cell counting
Blastocysts were exposed to 0.5% (w/v) pronase solution for 1 min to remove the zona pellucidae, and then washed three times for 5 min each in DPBS supplemented with 1 mg/mL PVA. The zona-free blastocysts were placed into a 1:5 dilution of rabbit–pig whole serum for 50–60 min and were then wash and transferred into a 1:10 dilution of guinea pig complement containing 10 μg/mL propidium iodide and 10 μg/mL Hoechst 33342 for 50–60 min. After being washed in DPBS at least five times, the stained blastocysts were loaded onto slides and observed under UV light with an epifluoresecnt microscope (Nikon E800, Japan). Cells stained blue and pink were considered to be ICM and TE cells, respectively, and cell numbers were recorded.
PCR detection of transgenic gene
Genomic DNA was isolated from a spectrum of tissues from each aborted animal (heart, liver, spleen, lung, kidney, ear, muscle, etc.) and newborn calves (ear, tail, and blood). PCR analysis was carried out using a pair of rhLF primers, that is, sense primer, 5′-CTC AAA GAA AGT CCC AAC CC-3′ and antisense primer, 5′-GTC TCT TGG ATG CGT TGC C-3′. Reactions were run for 30 cycles of 95°C for 30 sec, primer annealing at 62°C for 30 sec, and extension for 30 sec at 72°C, followed by extension for 7 min at 72°C; the amplified product was an 832-bp fragment. Samples were analyzed by separation in a 1.5% TAE agarose gel containing ethidium bromide.
Western blotting
The living female bovine was induced to produce milk with lactating agents (National Caotan Pharmacy Company, Xi'an, China). The hormone-induced milk was collected and stored at −20°C. The collected milk, nontransgenic milk, and natural human LF (Sigma, St. Louis, MO, USA) control were analyzed by SDS-PAGE and transferred to a nitrocellulose membrane for Western blotting analysis.
Statistical analysis
All values were presented as mean ± SD, unless indicated otherwise. Proportional data was analyzed by a chi-square test. Cell numbers were analyzed using the two-tailed t-test with equal variance. Differences were considered significant at p < 0.05.
Results
In vitro development of cloned versus chimeric embryos
In this experiment, two different types of donor cells (040825 and 050211) were used, and chimeric embryos were reconstructed at the eight-cell stage. As shown in Table 1, cloned and chimeric embryos had similar blastocyst rates; the average rates were 62% (1608/2576) for cloned and 65% (416/643) for chimeric embryos. There was also no significant difference between 040825 and 050211 cells for cloned and chimeric embryos (64 vs. 58% and 64 vs. 65%, respectively). Thus, no significant difference in the preimplantation development was detected between chimeric and cloned embryos.
Proportion of reconstructed embryos at the eight-cell stage developed into blastocysts.
Allocation of ICM and TE cells in cloned versus chimeric embryos
In the next experiment, we investigated the morphological differences between cloned and chimeric blastocysts. Blastocysts were differentially stained (Fig. 1), and the number of putative ICM and TE nuclei were quantified (Table 2). The number of ICM, TE, and total cells (TC) of chimeric blastocysts was 17.5, 25.9, and 23.7% higher, respectively, than those of cloned blastocysts (47 vs. 40, 73 vs. 58, and 120 vs. 97, respectively, p < 0.05), but the ICM:TC ratio was very similar (0.41 vs. 0.39).
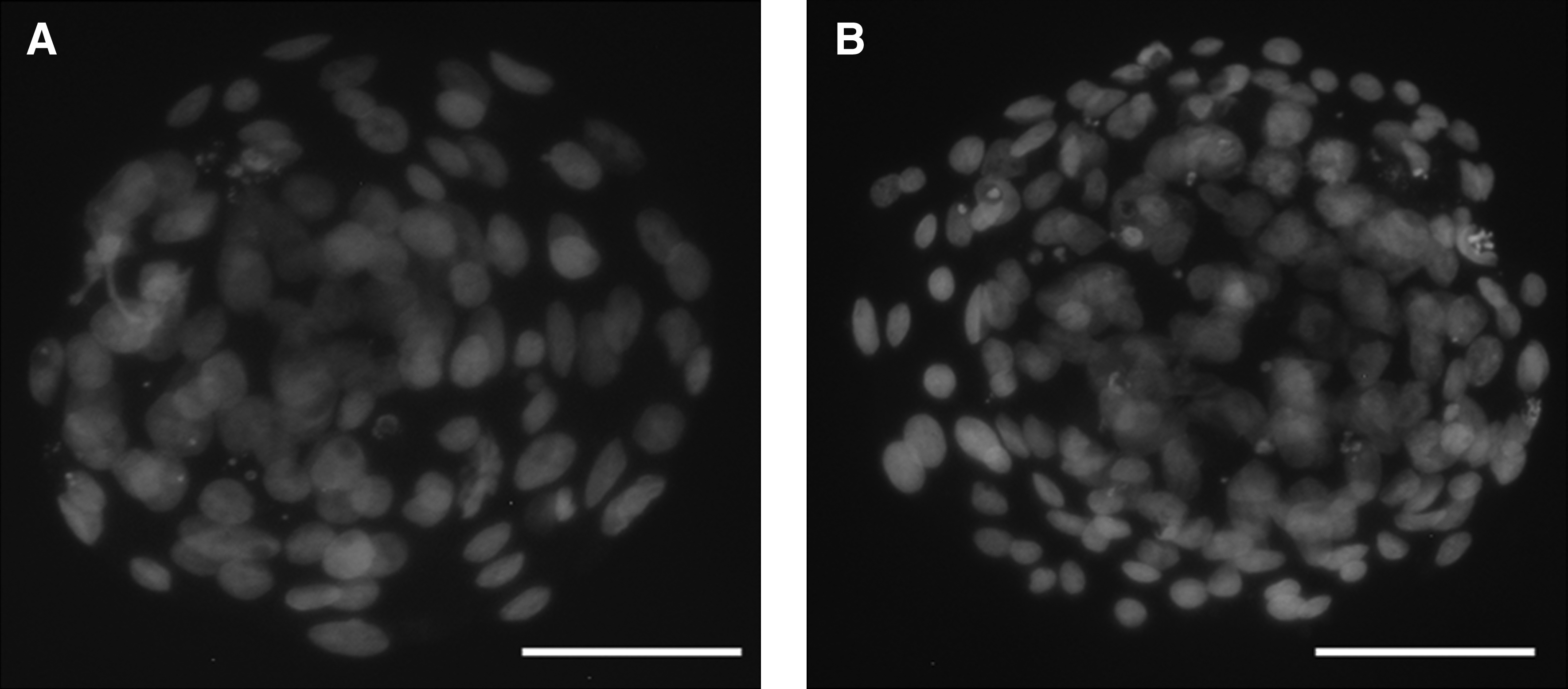
Representative images of differential stained of cloned and chimeric bovine blastocysts. All blastocysts were grade 1–2. Dark (Hoechst 33342) and light (propidium iodide) indicate ICM and TE cells, respectively. (
Values within columns with different superscripts are statistically different (p < 0.05).
In vivo development of cloned versus chimeric embryos
To correlate the results for in vitro development and blastocyst morphology, we transferred cloned and chimeric embryos into recipient cows. Pregnancy establishment at day 60 was significantly different for cloned (13.92%) versus chimeric (28.57%) embryos (Table 3) (p < 0.05). Development to full term or weaning was about threefold higher for chimeric embryos than for cloned embryos (17.46 vs. 6.96%, 6.35 vs. 1.9%, respectively, p < 0.05) (Table 3).
n = total number of independent NT experiments.
nET = total number of embryos transferred.
Proportion of nET developed into fetuses and live calves at day 60, term, or weaning.
Fifty-four blastocysts were transferred to 27 recipients with 2 blastocysts per recipient and 9 blastocysts were transferred to 3 recipients with 3 blastocysts per recipient.
Different superscripts within a column denote a significant difference (χ2, p < 0.05).
Distribution of IVF-derived cells in chimeric blastocysts
To chase the distribution of the IVF-derived blastomere in chimeric blastocysts, we microinjected a single GFP+-IVF-derived blastomere into GFP−-NT embryos at the eight-cell stage. In total, 216 GFP+-IVF × GFP−-NT hatching chimeric blastocysts were produced. In these 216 IVF × NT chimeric blastocysts, 143 IVF-derived blastomeres populated the ICM (66.2%), and the remaining 73 IVF-derived blastomeres distributed in the ICM and TE (33.8%) (Fig. 2).
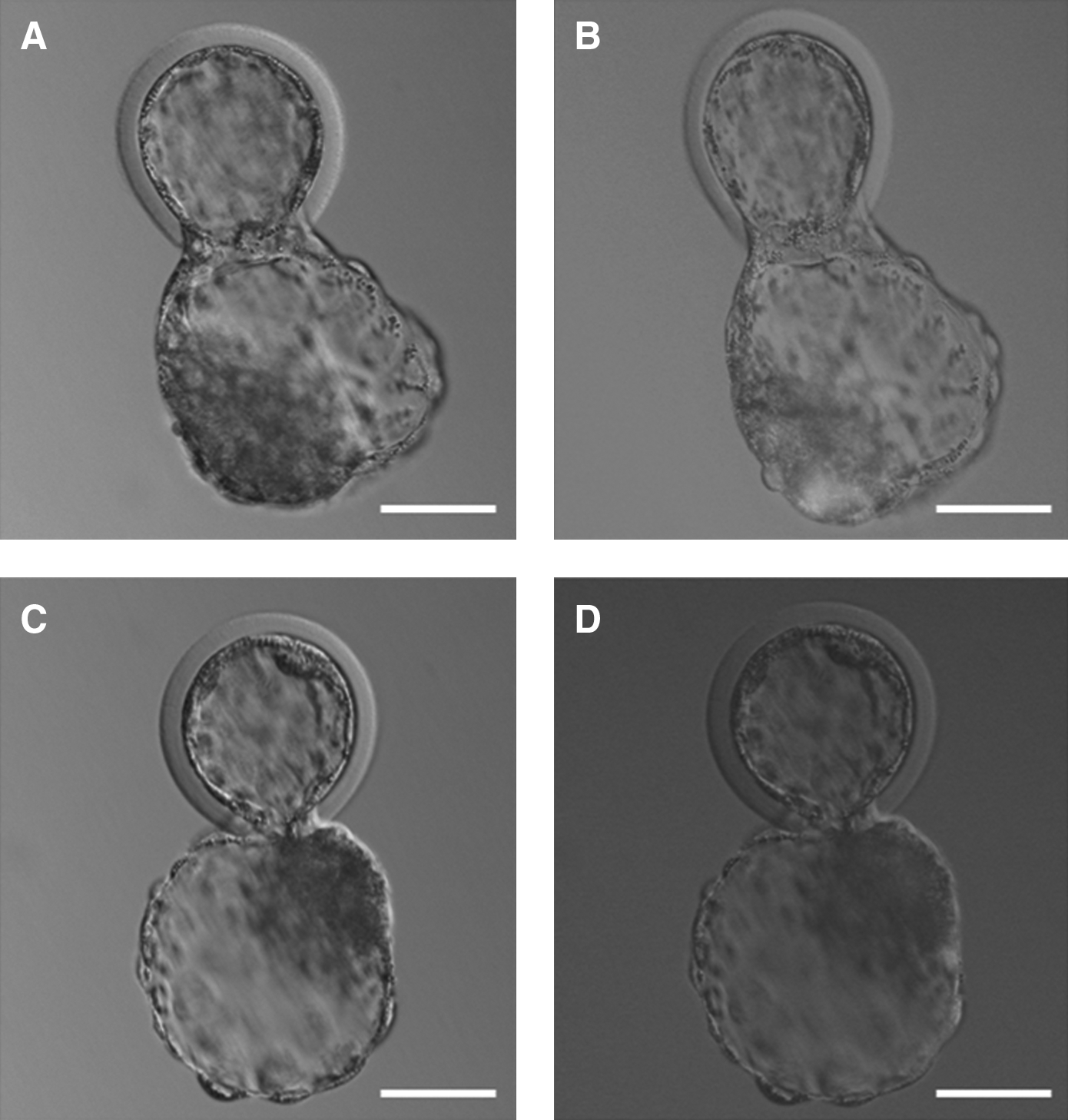
Representative images illustrating the distribution of GFP+-IVF-derived cells in IVF × NT hatching chimeric blastocysts. (
Offspring of chimeric embryos
Sixty-three chimeric blastocysts were transferred into 30 recipient cows; only 11 calves representing 5 singletons and 2 sets of triplets were produced. Unfortunately, one singleton was dead 5 days after birth because of respiratory distress syndrome and two sets of triplets were aborted at perinatal; two of them had renal insufficiency, the rest had no apparent abnormalities (Table 4). The remaining four calves are apparently normal and healthy, except that C3 has no tail (Fig. 3). Furthermore, the placentas we collected did not show obvious abnormities such as hydrallantois and placentomegaly. The weights of the 11 calves were all normal; no calves with “large offspring syndrome” were observed (Table 4).
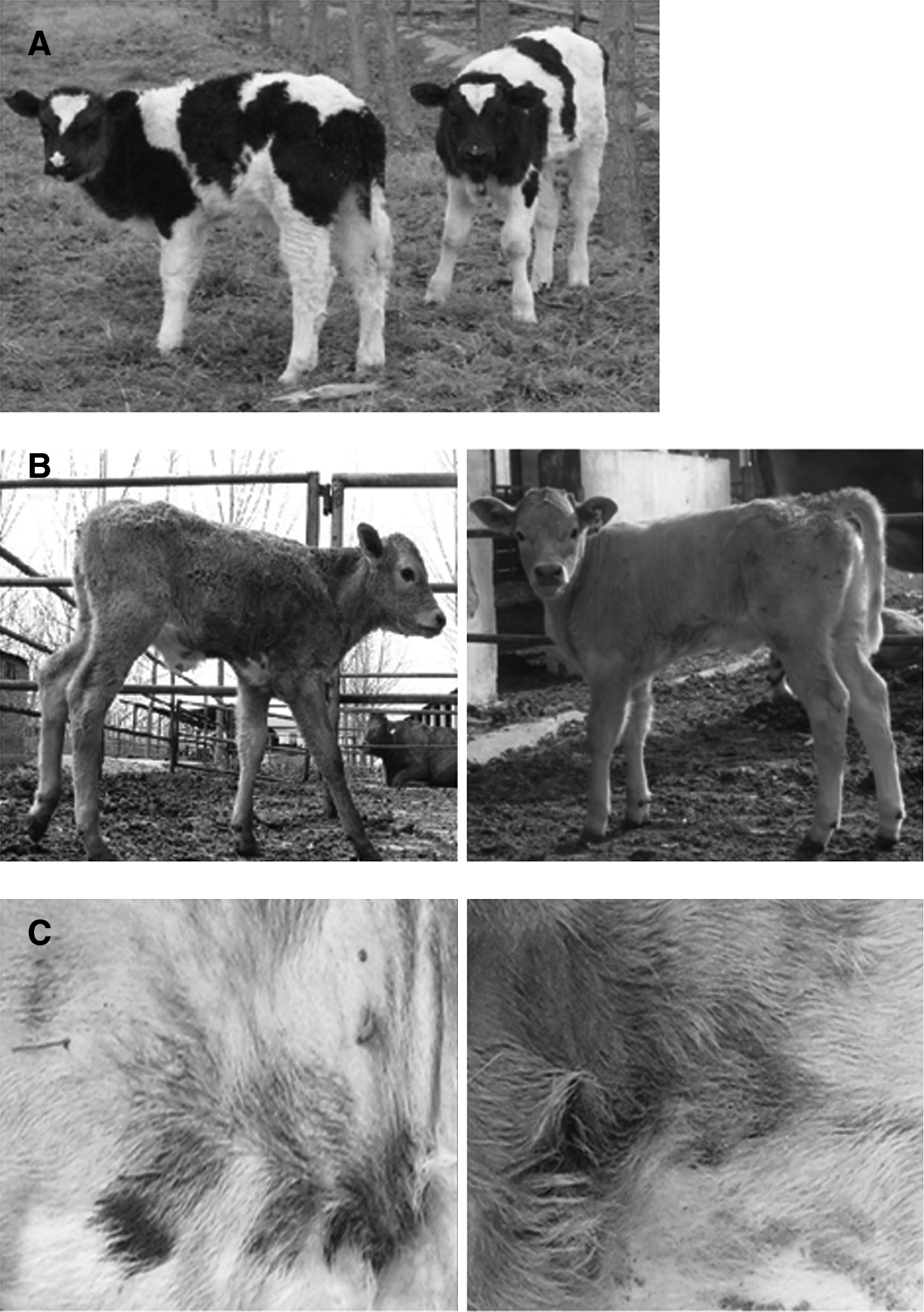
Living offspring from chimeric embryos derived from microinjection of a single IVF-derived blastomere into cloned embryos at the eight-cell stage. (
This calf died 5 days after birth because of respiratory distress syndrome.
For one recipient with triplet fetuses, abortion was induced at perinatal—one of them had renal insufficiency, the other two had no apparent abnormalities.
M, male; F, female; BW, black and white; G, gray.
The skin pigment color and hair color pattern were used as phenotypic confirmation of chimerism. Only stillborn calves C6-1 and C6-2 showed the chimeric phenotypic color pattern, their gray hair originated from IVF-derived cells, whereas dark hair originated from NT-derived cells, and the NT-derived dark hair only occupied with a small percentage compared with the IVF-derived gray hair (Fig. 3C). PCR showed that most of the collected tissues or organs of stillborn calves were positive for the rhLF transgene (Table 5). However, PCR results with ear, tail, and blood from newborn calves showed that C1 and C4 were positive for rhLF, indicating that C4 was also a chimera; its LF gene originated from the NT embryo, whereas the gray color originated from the IVF-derived blastomere. However, it is noteworthy that the majority of tissues were rhLF-negative (Fig. 4), that is, most tissues originated from IVF-derived nontransgenic cells. Furthermore, calf C1 is also a chimera; its male sex originated from IVF-derived cells, and its rhLF gene originated from NT-derived cells; we are not sure whether calves C1 and C2 were chimerism.

Detection of rhLF transgene in living calves by PCR. E: ear; LE: left ear; RE: right ear; T: tail; B: blood; S: sperm; E1: in vivo embryo; P: positive control; N: negative control; W: blank control; M: 100-bp DNA ladder marker.
H, heart; L, liver; S, spleen; L, lung; K, kidney; M, muscle; U, uterus; B, brain; S, stomach; G, gallbladder; MG, mammary gland; I, intestine; G, gonad; E, ear. The genomes of calves C7-1, 2, 3 were all degraded.
F, female; M, male; ND, not detected.
Germline and mammary gland chimerism
When the male offspring C1 and C2 were sexually mature, their sperm were collected and fertilized with matured oocytes. However, the sperm did not appear to be transgenic because the fertilized oocytes could have developed into blastocysts, but no GFP was observed in the fertilized oocytes under fluorescence microscopy, and the sperm were rhLF-negative according to the polymerase chain reaction (PCR) resluts. When the female offspring C3 and C4 were sexually mature, they were superovulated, inseminated, and lactated by lactating agents, and then blastocysts and milk were collected. Unfortunately, no GFP was observed under fluorescence microscopy, which was consistent with the failure to detect rhLF by PCR (Fig. 4). Western blotting also indicated that milk was negative for LF (Fig. 5). All of the above results suggest that the germline and mammary glands might have originated from IVF-derived nontransgenic cells.

Western blot analysis for expression of rhLF in the milk of C3 and C4. The proteins in collected milk and nontransgenic milk were separated on SDS-PAGE and then blotted and hybridized with antihuman LF. Lane 1, human α-LF; lane 2, protein marker; lane 3, C3; lane 4, C4; lane 5, nontransgenic control.
Discussion
We evaluated the effects of microinjection of a single IVF-derived blastomere on the development of a cloned embryo and compared the competency between NT- and IVF-derived cells in chimeric blastocysts and offspring. Our results demonstrated that an IVF-derived blastomere improved the in vivo developmental potential of cloned embryos in bovine, and that the IVF-derived blastomere was more competitive than NT-derived blastomeres, because they were better able to populate the ICM of chimeric blastocysts and many tissues of living offspring.
We microinjected an IVF-derived blastomere as early as the eight-cell stage because of four assumptions. (1) It is easier to micromanipulate a blastomere at the eight-cell stage than at the four-cell stage or other stages. (2) In cattle, the survival of NT embryos derived from eight-cell donor embryos was significantly better than those from 8-to-16-, 9-to-15-, 16-, 16-to-32-, 32- and 32-to-64-cell embryos (Willadsen et al., 1991), indicating that eight-cell-stage blastomeres have better reprogramming capacity than those of later stages. (3) In cattle, the onset of embryonic genome activation occurs at the 8-to-16-cell stage (Frei et al., 1989), and blastomeres isolated at the eight-cell stage can develop to full term (Willadsen and Polge, 1981). Thus, blastomeres isolated at the eight-cell stage have totipotency and may participate in forming embryonic and extra-embryonic tissues, thus complementing all of the disfigurements of the cloned embryos. (4) Using blastomeres earlier than the eight-cell stage may result in a higher percentage of IVF-derived nontransgenic cells. On the contrary, blastomeres later than the eight-cell stage, such as somatic cell-derived stem-like cells (Cibelli et al., 1998), may only participate in forming parts of fetal tissues and/or organs.
Cloned and chimeric embryos showed very similar blastocyst rates (Table 1). This is not surprising because any in vitro manipulation of an embryo tends to compromise its development. Rho et al. (2001) reported that micromanipulated four-cell bovine embryos, in which blastomeres had been separated and then recombined, formed blastocysts at a lower rate than unmanipulated embryos. However, it was reported that IVF-derived bovine blastomeres significantly enhanced the blastocyst rate of chimeric parthenogenetic embryos (Boediono et al., 1999). In our research, IVF-derived bovine blastomeres may also increase the blastocyst rate of chimeric embryos, but due to the antagonism between the negative role of micromanipulation and the positive promotional role of IVF-derived blastomeres, these chimeric embryos exhibited blastocyst rates similar to cloned embryos.
There are two kinds of distribution of IVF-derived cells in chimeric blastocysts. Researchers have found that most of the mouse embryonic stem (ES) cells microinjected into mouse four- or eight-cell-stage embryos, morulae, or blastocysts were incorporated into the ICM and only rarely into the trophectoderm (Beddington and Robertson, 1989; Huang et al., 2008; Saburi et al., 1997; Wood et al., 1993). In our study, unlike that of mouse ES cells, IVF-derived blastomeres microinjected into cloned embryos mainly incorporated not only into ICM but also simultaneously into trophectoderm (Fig. 4). In embryo aggregation experiments between ICM and early eight-cell mouse embryos, eight-cell blastomeres will attach to the ICM and migrate along the ICM until the ICM is completely engulfed (Kimber et al., 1982). Ziomek et al. (1980) proposed that when a polarized and a nonpolarized blastomere are aggregated, the polarized cell attempts to completely engulf the nonpolarized cell. Koyama et al. (1994) found that blastomeres from one- to eight-cell embryos in cattle showed no polarity, and the onset of transitional polarization of microvillous distribution occurred at the 9- to 15-cell stage. In our study, the reconstructed embryos were all from the nonpolarized eight-cell-stage blastomere, so the above theory could not explain this phenomenon. The inside/outside hypothesis proposes that cells on the morula surface will become TE cells, whereas inner cells become ICM cells (Tarkowski and Wroblewska, 1967). When the reconstructed embryos developed into a morula, the microinjected blastomere had divided into four to eight sister cells, which may locate to different positions in the chimeric embryos, so the distribution of IVF-derived cells in chimeric blastocysts may be associated with the position of IVF-derived cells in the morula.
Chimeric blastocysts have increased numbers of ICM, TE, and TC, but their ratios of ICM to total cell numbers were very similar to those of cloned embryos. In general, the mean ratios of ICM to total cells for bovine blastocysts produced in vitro or in vivo range from 10 to 40% (Iwasaki et al., 1990; Van Soom et al., 1997), whereas the mean ratios of ICM to total cells for most bovine NT blastocysts range from 40 to 70%. Koo et al. (2002) pointed that fewer TE cells in SCNT embryos lead to abnormal placenta formation. The ICM-to-total cell ratio for our chimeric blastocysts was about 40%, which was very close to the highest value for IVF blastocysts, indicating that the quality of chimeric embryos was very similar to IVF embryos. On the other hand, the ICM-to-total cell ratio for chimeric blastocysts was similar to that of cloned embryos, whereas the number of TE cells in chimeric blastocysts was higher than that of cloned blastocysts. Thus, in the situation of similar ICM-to-total cell ratios, the chimeric embryos with more TE cells would help the development of the placenta. When the offspring were born, the placentas of chimeric blastocysts that we collected did not show obvious abnormalities, such as hydrallantois, placentomegaly, and features of “large offspring syndrome.”
It has been speculated that fewer TE cells and increased ICM-to-TC ratios in SCNT embryos lead to placental abnormalities or early fetal losses (Koo et al., 2002). The results from Misica-Turner et al. (2007) do not support this hypothesis, but our results do. Misica-Turner et al. (2007) found SCNT embryos showed increased ratios after aggregation but no reduction in survival to term. In our experiments, chimeric blastocysts showed a slightly decreased ICM-to-TC ratio (0.39 vs. 0.41) after blastomere injection; correspondingly, fewer fetus losses (3/14 vs. 11/22) were observed between day 60 and term. The rates of development to day 60, to term, and to weaning of chimeric embryos were approximately threefold higher than those of cloned embryos, which indicated that the chimeric embryos were more competitive than cloned embryos. However, it is not known how the stimulatory effect of the IVF-derived bovine blastomere is mediated in these cloned embryos. The growth factors produced by the IVF-derived blastomere (Ducibella and Anderson, 1975) may have played a role in the enhanced development of the chimeric embryos in the present study. At the eight-cell stage, genomic activation begins, some paracrine and/or autocrine signaling factors such as insulin and members of the insulin-like growth factor family are present (Daliri et al., 1999; Watson et al., 1992), and the developing intercellular junctions between blastomeres of different origins (Ducibella and Anderson, 1975) enable intercellular communication and subsequent signaling. In general, aberrant reprogramming (including the lack of telomere restoration of donor chromatin, aberrant expression of specific genes, and aberrant methylation patterns) is thought to be the key factor affecting developmental failures of NT embryos (Niemann et al., 2008). Chimeric blastocysts theoretically have a higher chance of containing some fully reprogrammed cells. However, the underlying mechanism needs to be further investigated.
Although some cattle that were transgenic rhLF-positive in most of the collected tissues (including the mammary gland and gonad) were stillborn, those living cattle from which parts of the tissues were transgenic-positive were apparently normal and healthy, indicating that IVF-derived blastomeres could incorporate into ICM and differentiate into fetal tissues and/or organs, and thus could complement the deficiencies of cloned fetal tissues, reduce the incidence of abortion, and improve the health conditions of newborn cattle. Furthermore, the IVF-derived cells were more competitive, and they could even completely replace the NT-derived cells in most tissues and organs. In particular, the IVF and PCR results from the gametes of living offspring showed that the gametes were all from IVF-derived cells. Researchers found that ES cells injected into eight-cell-stage embryos efficiently yielded F0-generation mice that were fully ES cell-derived and exhibited 100% germline transmission (Dechiara et al., 2009; Huang et al., 2008; Poueymirou et al., 2007). Thus, the eight-cell-stage bovine blastomere from IVF embryos may have developmental competency similar to mouse ES cells.
In summary, microinjection of a single blastomere from IVF embryos at the eight-cell stage into cloned embryos enhanced the in vivo development of cloned embryos in bovine. However, IVF-derived nontransgenic cells occupied the chimeric tissues or organs at a high proportion or even totally replaced the cloned cells, indicating that blastomeres derived from eight-cell-stage IVF embryos were more competitive than those derived from cloned embryos in chimeric embryos. To lower the proportion of nontransgenic cells, blastomeres from the 16-cell stage or even later blastocysts may be more suitable. Even without a full explanation of the observations presented here, this technique will be still useful. Although the original hypothesis that production of an embryo with ICM of NT origin and trophectoderm of IVF origin has been dismissed, this technique can be used to evaluate interactions of different genotypes/mutations in early embryonic development.
Footnotes
Acknowledgments
We are grateful to Lili Wang, Haiping Wang, Jing Li, Fenglei Gao, Min Zheng, and Meili Wang for micromanipulation and to Dr. Guangbin Zhou for his suggestion on this article. This work was supported by the Postdoctoral Foundation of China (No. 20080440452) and the National High Technology Research and Development Program of China (863 Program) (No. 2008AA101006).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.