
Abstract
Abstract
The early events in the nuclear reprogramming process during somatic cell nuclear transfer (SCNT) consist of morphological remodeling of the donor nucleus including premature chromosome condensation (PCC). In the present study, the objective was to increase oocyte M-Phase Promoting Factor (MPF) kinase activity and to examine the fate of the donor nucleus and the development of SCNT embryos thereafter. Indeed, in controls, recipient oocytes activated upon nuclear transfer, undergo a decrease in MPF activity, responsible for the inability to promote PCC in 77.8% of reconstituted embryos. Here we showed that exposure of the recipient oocyte to the proteasome inhibitor MG132 prior to fusion inhibited the degradation of cyclin B, which normally occurred immediately after activation by electro stimulation, and therefore sustained a high level of MPF. Treatment with MG132 also significantly increased the percentage of SCNT embryos with PCC when compared to the nontreated SCNT control embryos (94.1 vs. 22.2%, respectively, p < 0.01). The frequency of development to the blastocyst stage did not differ between MG132-treated or untreated recipient oocytes. However, we observed a significant increase of the total cells number in embryos produced after MG132 treatment. Investigation of the global nuclear organization by immunodetection of heterochromatin protein 1 (CBX1) showed that SCNT embryos derived from MG132-treated recipient oocytes displayed organization patterns similar to the ones observed in IVF embryos in contrast to the nontreated SCNT controls. Taken together, these results suggest that the PCC induced by MG132 treatment allows reorganization of the chromatin at an appropriate time potentially, leading to better reprogramming.
Introduction
In most somatic cell nuclear transfer (SCNT) experiments, differentiated nuclei are transferred into enucleated oocytes at the metaphase II (MII) stage, where nuclear modifications and reprogramming take place. Early events in this process consist in morphological remodeling of the donor nucleus, which includes breakdown of the nuclear membrane, premature chromosome condensation (PCC), spindle assembly and the formation of pronucleus-like structures after activation (Campbell and Alberio, 2003; Peter et al., 1990, Wrenzycki et al., 2001). Previous reports suggested that the reprogramming of nuclei from differentiated cells only occurs completely and consistently, after nuclear envelope breakdown and chromosome condensation, initiated and sustained by a high level of M-Phase Promoting Factor (MPF), (Campbell et al., 2005). In mice (Wakayama et al., 1998), pig (Kawahara et al., 2005), and primates (Mitapilov et al., 2007), a higher in vitro development rate was obtained by inducing PCC, due to delayed activations after nuclear transfer; suggesting that PCC might promote effective nuclear reprogramming of the donor cell and also the developmental competence of SCNT embryos.
In cattle it is more controversial; some authors have reported the beneficial effect of a prolonged exposure to MPF of transferred nuclei to promote PCC (Shin et al., 2002; Wells et al., 1998); but successful reprogramming of nuclei, without PCC, has also been achieved after transfer into recipient oocytes simultaneously with or just prior to activation (Akagi et al., 2003; Sung et al., 2007; Vignon et al., 1998; Wilmut et al., 1997). Indeed, when SCNT embryos were submitted to immediate activation (simultaneous fusion and activation) 80% of transferred nuclei gradually swelled and formed pronuclear-like structures, whereas PCC was only observed in only 20% of reconstructed embryos, and interestingly, up to 38% of development to the blastocyst stage was obtained using such a protocol (Vignon et al., 2002). This may suggest that, in cattle, reprogramming of the somatic nucleus is not directly linked to the occurrence of PCC upon nuclear transfer (Sung et al., 2007; Tani et al., 2003); this has also been demonstrated in sheep, where the frequency of development to the blastocyst stage was not shown to differ whatever the rate of PCC (Lee and Campbell, 2006).
Altogether, it appears that we do not know whether PCC is essential for efficient reprogramming of a somatic nucleus in cattle and that the relationship between morphological remodeling, such as PCC, and development is still poorly understood.
Chromosome condensation and nuclear envelope breakdown are mediated by MPF activity, which needs be high in recipient oocytes to promote these processes (Szollosi et al., 1988). MPF activity in recipient oocytes thus seems to be an important factor for the development of cloned embryos. However, MPF activity in recipient oocytes and in SCNT embryos remains poorly understood. MPF is a heterodimer composed of P34 kinase and cyclin B, and its inactivation is triggered by the degradation of cyclin via the ubiquitin–proteasome pathway (Peters, 2002; Sherwood et al., 1993, Tokumoto et al., 1997). MG132, a specific proteasomal inhibitor, can inhibit the degradation of cyclin B and thus maintains high MPF activity (Chesnel et al., 2006; Josefsberg et al., 2000). This inhibitor has been shown to promote a high level of MPF activity in recipient oocytes in various species (rat: Ito et al., 2005; mice: Yu et al., 2005; goat: Wu et al., 2007; primates: Mitapilov et al., 2007), and it was the only specific condition necessary to obtain SCNT rat embryos (Zhou et al., 2003).
During the present study we examined a strategy to further aid the process of reprogramming in bovine SCNT embryos by preserving a high level of MPF activity into the recipient oocyte using MG132 prior to fusion with the somatic cell. Embryos reconstructed with oocytes treated with or without MG132 were compared using biochemical criteria such as MPF activity and cyclin B behavior. We then determined the occurrence of PCC after activation and fusion as well as the developmental potential of SCNT embryos derived from recipient oocytes with or without MG132 treatment. Finally, to evaluate global nuclear organization at the time of embryonic genome activation (EGA, i.e., at the four- to eight-cell stage in bovine; Kopecny et al. 1989) we analyzed the distribution of pericentromeric heterochromatin by immunostaining of heterochromatin protein 1(CBX1). This protein is known to participate in the propagation of heterochromatin into nuclei (Cheutin et al., 2003; Hayakawa et al., 2003) and underlines the formation of nuclear clusters, called chromocenters, at EGA both in mouse and bovine embryos (Martin et al., 2006; Pichugin et al., 2010). However, we previously observed precocious heterochromatin condensation in bovine SCNT embryos during preimplantation development (Pichugin et al., 2010) and hypothesized that it may correspond to poor reprogramming of the transferred nuclei.
We found that MG132 maintained MPF activity at a high level by inhibiting cyclin degradation and promoted PCC in almost all recipient oocytes treated with MG132. The MG132-induced PCC did not appear to be detrimental for the development of SCNT embryos, and nuclear organization at the four-cell stage seemed to be well reprogrammed, like in IVF embryos.
Materials and Methods
All experiments were performed in accordance with the International Guiding Principles for Biomedical Research Involving Animals, as promulgated by the Society for the Study of Reproduction and with the European Convention on Animal Experimentation. All chemicals and reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise stated.
Preparation of in vitro matured oocytes
Bovine ovaries were collected from a local slaughterhouse and transported to the laboratory in phosphate-buffered saline (PBS) at 30°C within a period of 2 h. Immature oocytes were aspirated from antral follicles (2–7 mm in diameter) with a 19-gauge needle. Cumulus–oocyte complexes with more than three compact layers of cumulus cells were selected, pooled, and rinsed in HEPES-buffered TCM199 and then in maturation medium 25 mM bicarbonate-buffered TCM 199 supplemented with 10% fetal calf serum (FCS) (Gibco, Grand Island, NY, USA), 10 μg/mL FSH (NIDDK, Stimufol, Sart-Tilman, Belgium), 10 μg/mPLH (NIDDK, Beckers) and 1 μg/mL estradiol 17β. The oocytes were then cultured for 22 h at 39°C in a humidified atmosphere of 5% CO2 in maturation medium. Twenty-two hours after the beginning of maturation (hpm), the oocytes were incubated in HEPES-buffered TCM199 supplemented with 0.5% (w/v) hyaluronidase for 5 min; gentle pipetting was then performed to remove the cumulus cells. Oocytes were then assigned randomly in two groups: parthenogenetic activation and nuclear transfer.
MG132 treatment
To determine the cytotoxicity of MG132 on bovine embryo development, we used parthenogenetic embryos as controls. Metaphase II oocytes (MII) were selected and incubated in 25 mM bicarbonate-buffered TCM199 supplemented with 10% (v/v) FCS and different concentrations of MG132 (0, 2.5, and 5 μM; Calbiochem, Germany) for 45 or 90 min and then washed in HEPES-buffered TCM199 and activated immediately. MII oocytes were activated by electrostimulation in a 0.3 M mannitol solution containing 0.1 mM Ca2+ and Mg++, applying double-direct current pulses of 2.0 kV/cm for 30 μsec at 1-sec intervals using a Grass S44 stimulator. They were then incubated in 25 mM bicarbonate-buffered TCM199 supplemented with 10% FCS, containing 5 μg/mL cytochalasin B and 10 μg/mL cycloheximide for 5 h and rinsed sequentially in 25 mM bicarbonate-buffered TCM199 supplemented with 10% (v/v) FCS and in B2 medium (Laboratories CCD, France) with 2.5% (v/v) FCS. Activated oocytes were then transferred into micro droplets of B2 medium supplemented with 2.5% (v/v) FCS and seeded with monkey liver Vero cells at 39°C under 5% CO2 in humidified air (Menck et al., 1997). Cleavage was assessed at day 2 after activation and the blastocyst stage was evaluated at day 7. Noncleaved oocytes were fixed at day 2, DNA was stained with 10 μg/mL propidium iodide, and examined using laser confocal microscope to determine the exact activation rate (Zeiss LSM 310; MIMA2 Platform, INRA).
Culture and preparation of fibroblast cells for nuclear transfer
The cells used in this study came from fibroblast cultures initiated from an ear of a 5-month-old calf as previously described (Vignon et al., 1998). Frozen cells at passage 4–10 were thawed and the cells were allowed to grow for 5 days in DMEM supplemented with 10% (v/v) FCS until confluency to synchronize at the G0/G1 stage of the cell cycle by contact inhibition. Nuclear donor cells were then dissociated by 5 min of trypsinization at 37°C, resuspended in 1 mL DMEM containing 10% (v/v) FCS and maintained in this medium until nuclear transfer.
Somatic cell nuclear transfer and in vitro culture
Before use, MII oocytes were incubated for 45 min with two different concentrations of MG132: 2.5 μM and 5 μM. During this 45-min period of MG132 treatment, MII oocytes were first stained with Hoechst 33342 (1 μg/mL) for 10 min in 25 mM bicarbonate-buffered TCM 199, 10% (v/v) FCS, and then enucleated in HEPES-buffered TCM199, 10% (v/v) FCS, and 5 μg/mL cytochalasin B. The control group was subjected to the same protocol but without MG132. At the end of the 45-min MG132 treatment, one somatic cell was inserted under the zona pellucida of the enucleated oocyte in HEPES-buffered TCM199 + 10% (v/v) FCS and fused by electrostimulation as for parthenogenetic activation (described above). Reconstructed embryos were incubated immediately after fusion in the presence of 5 μg/mL cytochalasin B and 10 μg/mL cycloheximide for 5 h. SCNT embryos were cultured in vitro and evaluated as described above for activated oocytes.
Total cell counting
Blastocysts from each group (5 μM MG132 and control) were fixed in cold methanol for 15 min and the DNA was stained with 10 μg/mL propidium iodide. Entire blastocysts were scanned with 10 μm optical sections. Confocal microscopy was performed with a Zeiss LSM 310 laser-scanning microscope (MIMA2 Platform, INRA). Nuclei were counted using Image Tool software.
Immunofluorescence
One-cell (1 h postfusion) and four-cell (36 h postfusion) SCNT embryos were fixed in 4% (w/v) paraformaldehyde (PFA) and then washed for 10 min in PBS and permeabilized with 0.5% (v/v) Triton X-100/PBS solution for 30 min at room temperature. All samples were then blocked with 2% (w/v) BSA–PBS for 30 min and then incubated for 1 h at 37°C with the first antibody. Then, SCNT embryos were rinsed extensively in PBS–BSA and incubated for 1 h at 37°C with the secondary antibody followed by a further extensive wash with PBS. One-cell embryos were then exposed for a few minutes to 10 μg/mL propidium iodide. The one-cell and four-cell SCNT embryos were mounted on glass slides with antifading medium (Vectashield, Vector Laboratories, Burlingame, CA, USA).
Microtubules were labeled with a mouse monoclonal anti-α tubuline antibody (Amersham, France; 1:500 in PBS–BSA) and a fluorescein isothiocyanate (FITC)-labeled anti-mouse secondary antibody (Biosys, Vector Labs; 1:500 in PBS–BSA). CBX1 protein was detected with a mouse monoclonal anti-CBX1 antibody (Euromedex, Souffelweyersheim, France; clone 1MOD-1A9-As, 1:200 in PBS–BSA) and a rhodamine-conjugated anti-mouse secondary antibody (Jackson Immunoresearch, France; 1:200 in PBS–BSA).
The embryos were observed under a Zeiss LSM 510 confocal microscope (MIMA2 Platform, INRA).
Western Blot analysis
One hour or 2 h after activation by electric pulses (set of 20), control and MG132 treated-oocytes, enucleated or not, were lysed in sodium dodecyl sulfate (SDS)-sample buffer and polypeptides were separated using 10% (w/v) SDS polyacrylamide gel electrophoresis (SDS-PAGE) as previously described by Laemmli (1970). Stained proteins of known molecular weights (range: 14–200 kDa, Amersham) were run simultaneously as standards. The polypeptides separated by electrophoresis were transferred onto a polyvinylidene difluoride membrane (hybond-P PVDF; Amersham). The membrane was blocked by incubation in PBS 1/1000 Tween-20 (PBS-T; Prolabo, France) containing 4% (w/v) nonfat dried milk and incubated for 2 h at room temperature with a mouse monoclonal antibody directed against cyclin B (as previously described in Hue et al., 1997) diluted 1/1000 in PBS-T 4% (w/v) nonfat dried milk. The blot was then washed three times with PBS-T and incubated in PBS-T 4% (w/v) nonfat dried milk containing the peroxydase-conjugated anti-mouse IgG antibody (1/5000, Biosys, Vector Labs) and washed extensively in PBS-T. Peroxydase activity was revealed using the ECL-Plus Western blotting detection system as instructed by the manufacturer (Amersham). Immunoreaction signals were analyzed using an image analysis system (Advanced Image Data Analyzer software, LAS 1000 camera, Fuji film). Each experiment was repeated at least three times.
Histone H1 kinase assay
Histone H1 kinase activity was measured as previously described (Kubiak et al., 1992). Control embryos and embryos derived from MG132-treated oocytes were lysed individually just after and 1 h after activation by electrostimulation, in 80 mM β-glycerophosphate, 20 mM EGTA, pH 7.3, 15 mM MgCl2, 1 mM DTT supplemented with protease inhibitors and incubated for 45 min at 37°C in the presence of 3.3 mg/mL histone H1, 1 mM ATP, and 0.25 mCi/mL γ32P-ATP (Amersham). The reaction was stopped by adding 3× SDS-sample lysis buffer and boiling for 5 min. The samples were analyzed in 10% SDS-PAGE. Dried gels were finally exposed to a sensitive screen and radioactive signals were revealed with a FLA-3000 phosphorimager (Fujifilm, Raytest, Paris, France). Band intensity was quantified using Advanced Image Analyzer software (Fujifilm). Six independent series of experiments were performed and during each experiment 10 oocytes at each stage were analyzed individually.
In vivo development of SCNT blastocysts
SCNT blastocysts derived from MG132-treated recipient oocytes (n = 5 blastocysts) and control SCNT blastocysts (n = 5 blastocysts) were transferred to recipient heifers (one blastocyst per recipient), as previously described (Heyman et al., 2003) in order to evaluate their in vivo potential into offspring. Pregnancy diagnosis was first assessed by a plasma progesterone test on day 21. Using ultrasonography, the pregnancy status of recipient was then determined on day 35 of gestation and confirmed every 30 days from day 35 to day 90. Pregnant recipients were kept until calving.
Statistical analysis
The developmental rates of parthenotes and SCNT embryos as well as nuclear behavior were analyzed using χ2 analysis. Cell numbers were compared using Student's t-test. Statistical significance was accepted for p < 0.05 unless otherwise specified in the text.
Results
Cytotoxicity of MG132 regarding in vitro development
Before starting to use MG132 for SCNT, cytotoxicity was evaluated on bovine parthenogenetically activated oocytes. The effect of various durations and concentrations of MG132 treatment is shown in Table 1. When compared to the control group, treatment with MG132 for 90 min significantly reduced the activation rate and altered cleavage as well as blastocyst rates, whichever MG132 concentration was used. However, an incubation period of 45 min with MG132 did not reduce cleavage and blastocyst rates in any groups. Likewise, MG132 treatment for 45 min at 2.5 μM or 5 μM generated a blastocyst rate similar to that observed in the control group. We therefore performed all the subsequent MG132 treatments for 45 min.
The percentages of activated, cleaved, and blastocysts were calculated from the number of matured oocytes.
Within columns, values with different letters are statistically different (p < 0.001).
In vitro development of SCNT embryos using MG132-treated recipient oocytes
The effect of MG132 treatment on the in vitro development of SCNT embryos was examined. Three different treatments were used to prepare recipient oocytes, based on the above results: 0 μM (control group), 2.5 μM, and 5 μM MG132, for 45 min. As shown in Table 2, no significant difference was observed between the three groups in terms of fusion and cleavage rates. Similarly, development to the blastocyst stage of SCNT embryos derived from MG132-treated recipient oocytes, using 2.5 μM or 5 μM MG132, did not differ from the control group: 52.2, 53.3, and 50.5% respectively.
Percentages of fused embryos were based on the number of oocytes.
Percentages of cleaved embryos and blastocysts were based on the number of fused embryos.
Microtubules and genome reorganization after fusion
The state of the transferred nucleus was examined in SCNT embryos derived from recipient oocytes with or without MG132 treatment. We therefore performed immunostaining of the microtubules and counterstained the DNA to evaluate whether the transferred nuclei had undergone PCC or not, 1 h after fusion. Representative examples are shown in Figure 1. Results summarized in Table 3 show a higher rate of PCC in the 5 μM MG132-treated group (n = 51) than in 2.5 μM (n = 37) and in controls (n = 54): 94.1 vs. 54.1 vs. 22.2% (p < 0.01). Moreover, in almost all the embryos derived from MG132-treated oocytes microtubules were well reorganized around the condensed chromosomes after PCC, forming a metaphasic like spindle. In light of these results, we chose to use the concentration of 5 μM MG132 during subsequent experiments.
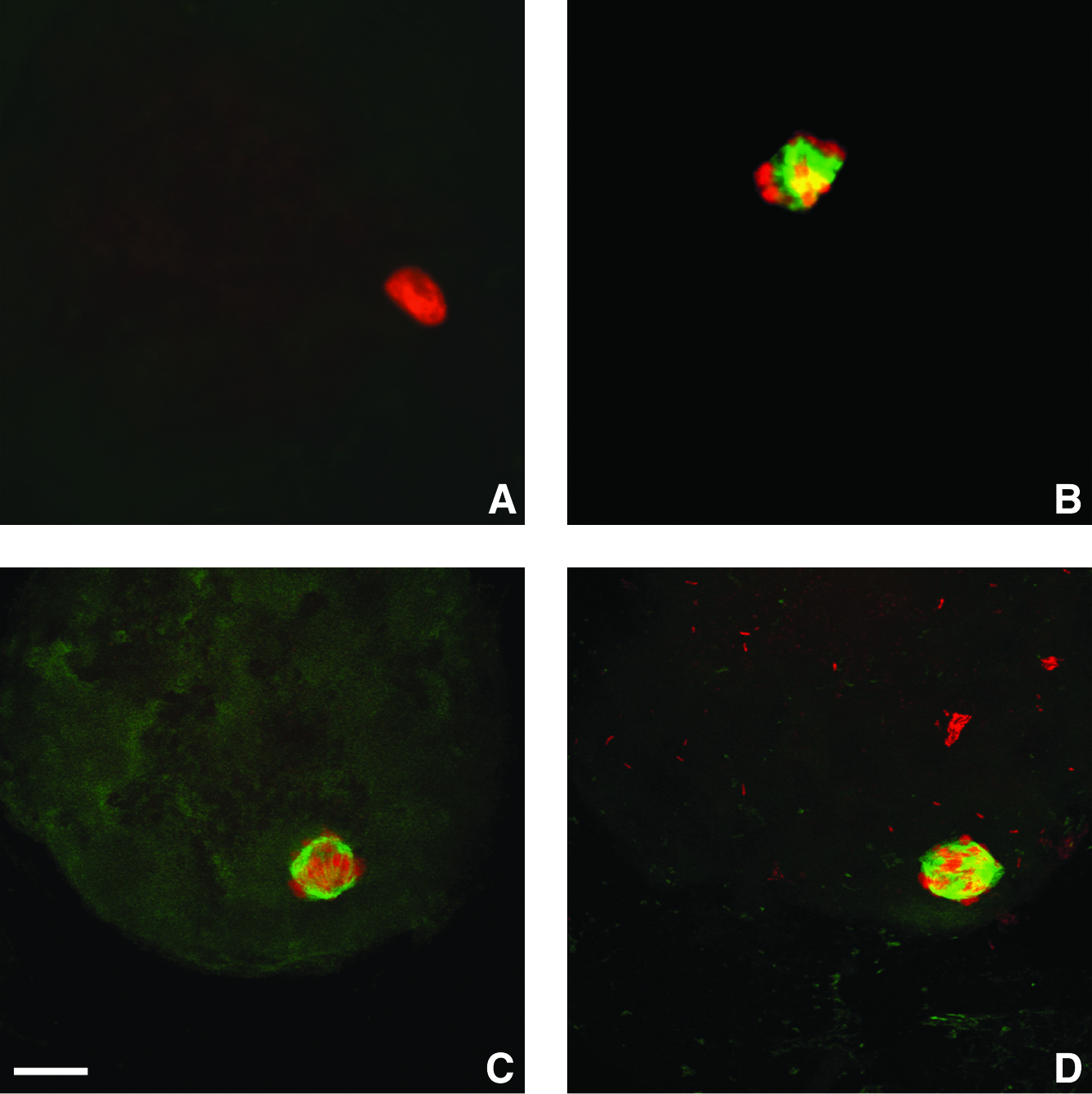
Laser-scanning confocal images of bovine SCNT embryos 1 h after fusion using treated and untreated recipient oocytes. Green, microtubules; red, chromatin. (
Values with different letters are significantly different (p < 0.01).
Total cells number in SCNT embryos
The total number of cells in SCNT blastocysts derived from 5 μM MG132-treated oocytes (grade 1 blastocysts, according to IETS classification) was significantly higher than that in blastocysts obtained using untreated recipient oocytes: 133 ± 24 and 104 ± 40, respectively (Table 4).
Within columns, values with different letters are statistically different (p < 0.05).
Effect of MG132 on cyclin B expression
Oocytes with or without MG132 treatment, enucleated or not, were parthenogenetically activated and analyzed 1 and 2 h after activation by immunoblotting, in order to determine the effect of MG132 on cyclin B expression in recipient oocyte. The immunoblot, using an antibody directed against cyclin B protein, revealed specifically a 65- kDa polypeptide in bovine oocytes (Fig. 2). Cyclin B was detected in control nonactivated MII oocytes as attempted. One hour after activation, cyclin B could not be anymore detected in parthenotes derived from nontreated oocytes, with or without enucleation. On the other hand, cyclin B remained present in parthenotes derived from MG132-treated oocytes, enucleated or not. Finally cyclin B could not be revealed in any group 2 h after activation (Fig. 2). These results thus show that MG132 prevented the degradation of cyclin B normally occurring after activation by electrostimulation.
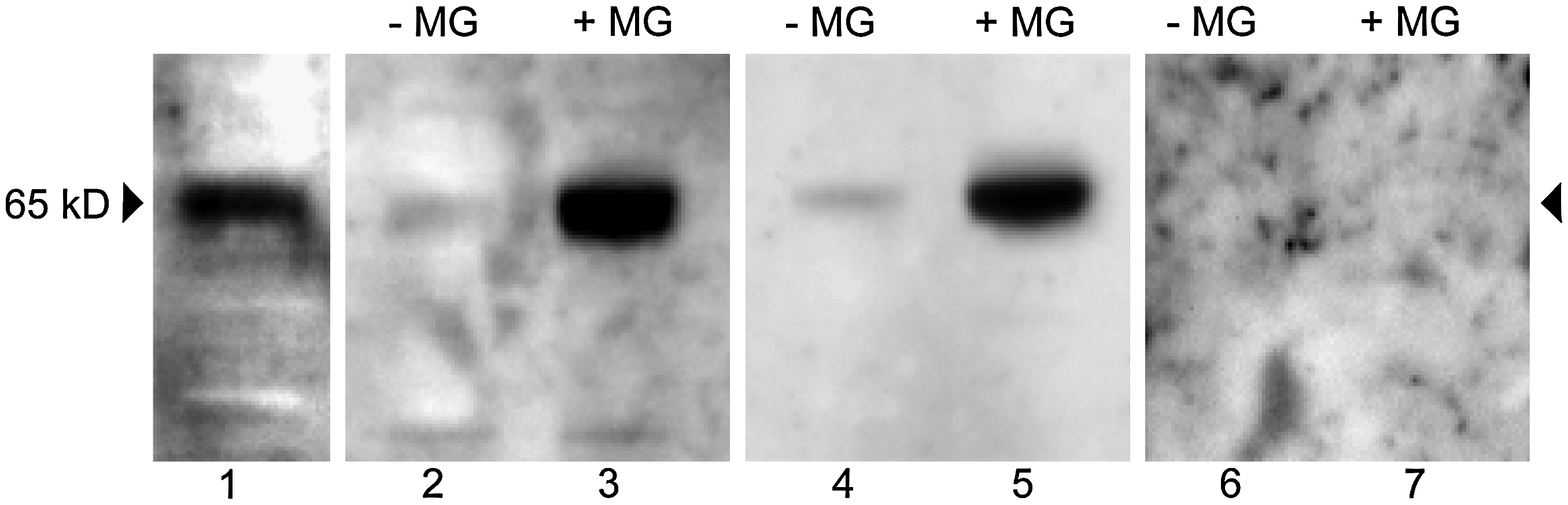
Immunodetection of cyclin B. Oocytes with or without MG132 (45 min, 5 μM), enucleated or not, were artificially activated and analyzed 1 h or 2 h after activation by immunobloting using anti-cyclin B antibody. Lane 1: oocyte at the MII stage used as a control; lanes 2, 3: oocytes 1 h after activation without (−MG) or with (+MG) MG132; lanes 4, 5: enucleated oocytes 1 h after activation without (−MG) or with (+MG) MG132; lanes 6, 7; oocytes 2 h after activation without (−MG) or with (+MG) MG132. After activation without MG132 cyclin B was no longer detected in oocytes (enucleated or not) from 1 h after activation but persisted in MG132-treated oocytes. Two hours after activation with and without MG132, cyclin B was no longer revealed.
Effect of MG132 on MPF activity
MPF activity was assayed in SCNT embryos derived from recipient oocytes with or without MG132 treatment, just after fusion and 1 h after fusion. As shown in Figure 3, MPF activity dropped rapidly to a basal level just after fusion in control SCNT embryos but was maintained at a high level in SCNT embryos derived from MG132-treated oocytes. One hour after fusion, MPF activity decreased and reached a basal level in both groups (with or without MG132; Fig. 3). This shows that MPF activity decreases rapidly after the electric pulses inducing fusion and that MG132 is able to inhibit this reduction in MPF activity.
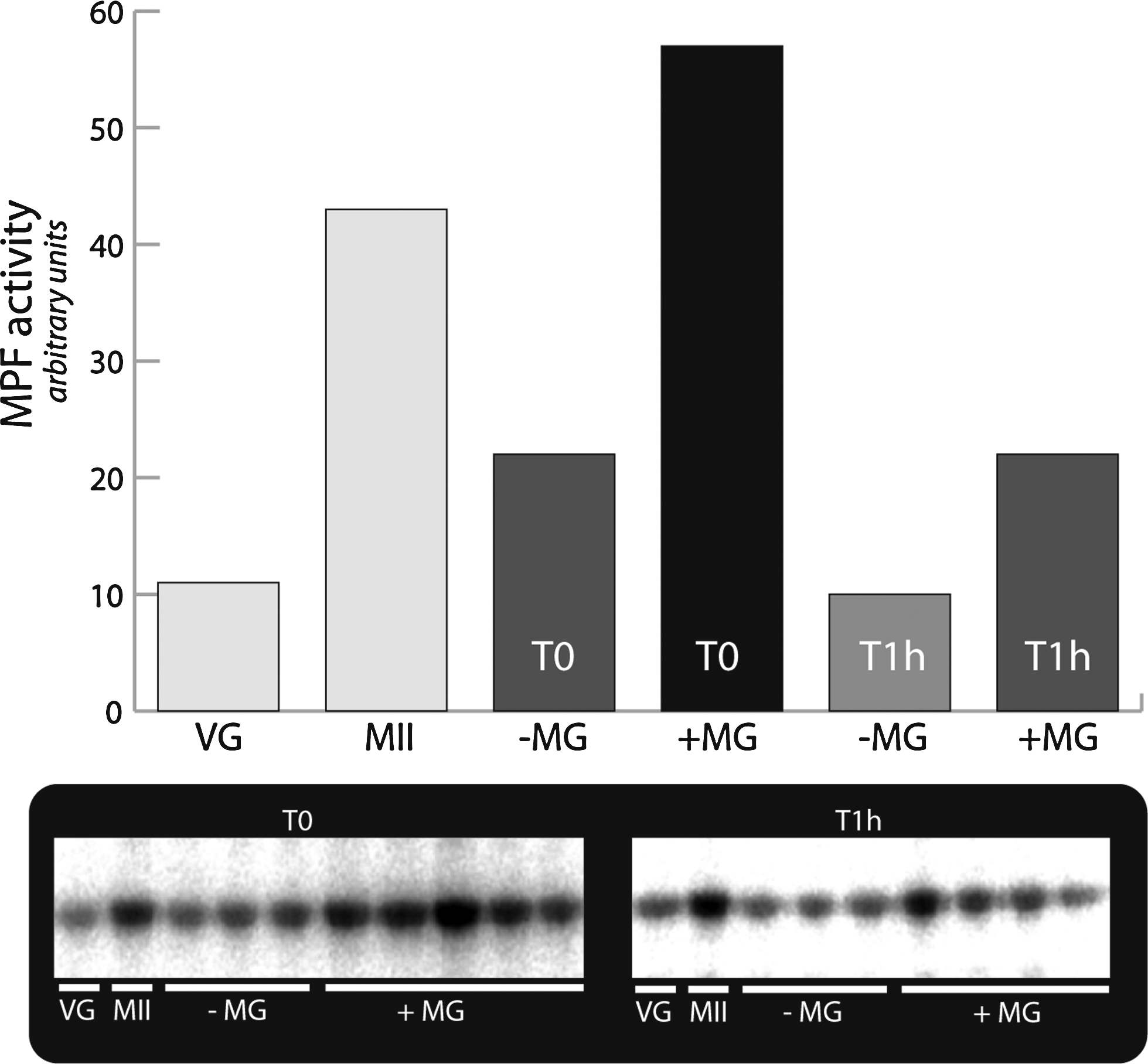
MPF activity. In embryos without MG132 (−MG), just after fusion (T0), MPF activity dropped rapidly to a basal level, while in MG132-treated embryos (+MG), MPF activity was maintained at a high level. One hour after fusion, in both groups (+MG, −MG), MPF activity was low. Oocytes at the germinal vesicle stage (VG) and at the MII stage were used as controls. In SDS gels, each sample corresponds to a single reconstituted embryo. The histogram represents the average intensity of the bands (arbitrary units) assessed by densitometric analysis (Lecphor Biocom).
Heterochromatin organization in SCNT embryos
Nuclear distribution of CBX1 protein was investigated at the four-cell stage. As previously described (Pichugin et al., 2010), three types of heterochromatin distribution pattern could be distinguished at that stage: spread, fibrous, or globular. In this study, a total number of 99 nuclei for the control group and 106 nuclei for the treated group were classified based on this morphological criterion (representative examples are shown in Fig. 4). Data show that the spread pattern was less dominant in control SCNT than in embryos derived from MG132-treated oocytes: 27.3% (27/99) vs. 53% (56/106)). The fibrous pattern was equally distributed within the two groups: 46.5% (46/99) vs. 39.6% (42/106). Interestingly, significantly more embryos with globular CBX1 blocks were observed in the control group in comparison to the treated group: 26.2% (26/99) vs. 7.5% (8/106). In our previous study we observed that the spread pattern was also the most prominent one in IVF embryos (70%), whereas the globular pattern was observed in only 12% of the in vitro fertilization (IVF) embryos (Pichugin et al., 2010). Altogether, these results suggest a delayed heterochromatin condensation in SCNT embryos derived from MG132 oocytes and improvement of pericentric heterochromatin reprogramming after nuclear transfer.
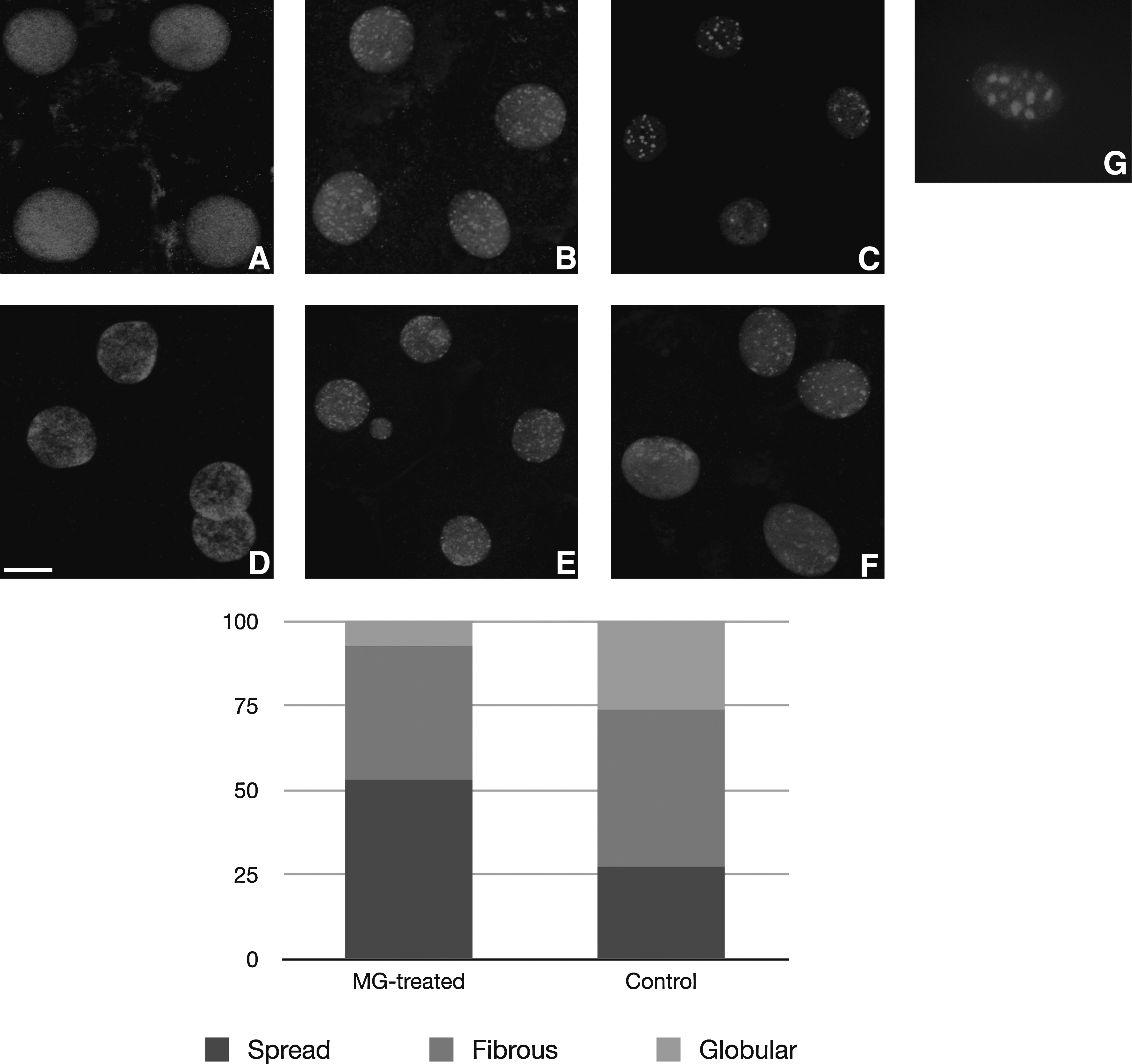
Distribution of CBX1 in four-cell stage SCNT embryos and fibrobalst donor cells. Pattern 1: the spread pattern, characterized by uniform CBX1 staining
In vivo development of treated and untreated SCNT embryos
In order to test in vivo development potential, five grade 1 SCNT blastocysts derived from MG132-treated oocytes and five grade 1 control SCNT blastocysts were transferred into 10 recipient heifers (one embryo per recipient). Because of the small samples size, no statistical analysis could be performed. However, in the MG132 group, four recipients were confirmed as being pregnant by ultrasonography on day 35, whereas only two were confirmed pregnant in the control group. At full term, one calf was delivered from the MG132 group and none from the control group.
Discussion
The present study shows that when SCNT bovine embryos were derived from oocytes treated with MG132 prior to nuclear transfer, a significant increase in nuclear envelope breakdown and PCC occurred after introduction of the donor nucleus into the recipient oocytes. After SCNT, the frequency of development to the blastocyst stage did not differ between MG132-treated and untreated recipient oocytes. However, we observed a significant increase in the total cells number in embryos produced after MG132 treatment, which is an indicator of embryo quality. As it had previously been described that nuclear transfer induces an increase in cell cycle length, resulting in a decrease in total cell numbers (Holm et al., 2003), we can hypothesize that MG132 induced a reduction in cell cycle length, but cells number increase could also have been due to a decrease in apoptosis. At full term, one calf was delivered from the MG132 group and none from the control group. Although these are only preliminary results with few transfers, it shows that MG132 inducing PCC is compatible with embryo survival and full term development.
Depending of the strategy chosen for embryo reconstruction, transferred G1 nuclei retain their initial state or undergo nuclear envelope breakdown, lamina disassembly, and PCC before being remodeled into a one-cell stage nucleus. In this context, MPF activity is known to play a key role in the development of SCNT embryos, particularly by inducing nuclear envelope breakdown and PCC (Campbell and Alberio, 2003; Du et al., 2002; Kwon et al., 2008; Peter et al., 1990). The inactivation of MPF following activation is considered to depend on cyclin B degradation via the ubiquitin/proteasome pathway (Chesnel et al., 2006; Peters, 2002; Sherwood et al., 1993; Tokumoto et al., 1997). In our laboratory, SCNT embryos are subjected to immediate activation (inducing fusion simultaneously) and we observed that thereafter, cyclin B and MPF activity decrease rapidly thus explaining the inability to induce PCC under these experimental conditions (22.2% PCC only). It is important to note that enucleation of oocytes before nuclear transfer could also be implicated in the rapid decrease of MPF activity through the removal of environmental proteins associated to the metaphase plate.
To maintain MPF activity in recipient oocytes, we treated these oocytes prior nuclear transfer with MG132, a proteasome inhibitor known to inhibit cyclin degradation and thus to induce higher MPF activity (Chesnel et al., 2006; Glotzer et al., 1991). We observed that in SCNT embryos derived from MG132-treated oocytes, the level of MPF activity, just after electrostimulation, was higher than in control SCNT embryos and declined to a basal level only 1 h after fusion. Furthermore, in treated recipient oocytes, cyclin B level remained high for at least 1 h after electrostimulation. This shows that MG132 inhibit the cyclin B degradation that normally occurs immediately after electrostimulation and therefore sustains a high level of MPF. These findings are in agreement with those of Huo et al. (2004), who described that cyclin B degradation was also inhibited by MG132 treatment in pig oocyte. Finally, we showed that with addition of MG132, inducing MPF activity maintenance, more than 90% of SCNT embryos presented a characteristic PCC.
It is now generally agreed that nuclear architecture contributes to functional gene regulation, and that higher-order chromatin organization such as heterochromatin clusters is implicated in epigenetic gene expression control. In somatic cells, pericentric regions from several chromosomes have been shown to form blocks of constitutive heterochromatin called chromocenters, which can be detected by CBX1 labeling. This protein, CBX1, therefore appears to be a good marker of heterochromatin organization in a given epigenetic state and in chromatin reprogramming following nuclear transfer. After in vitro fertilization, in cattle, the establishment of chromocenters has been correlated with transcriptional activation of the embryonic genome (EGA) occurring at the eight- to sixteen-cell stage (Martin et al., 2006). Interestingly, it has recently been demonstrated in our laboratory that the kinetics of CBX1 labeling differs between IVF and SCNT bovine embryos. After a dramatic reorganization of heterochromatic blocks following nuclear transfer, embryos were shown to display condensed hetrochromatin blocks at the two- to four-cell stage, that is, sooner than in IVF embryos, which reflected probably an incomplete reprogramming of the donor nucleus (Pichugin et al., 2010). In the present experiments, we investigated nuclear CBX1 distribution at the four-cell stage in both control SCNT embryos (with 22.2% PCC) and MG132-derived SCNT embryos (with 94.1% PCC). In SCNT control embryos, we confirmed that condensed heterochromatin blocks were displayed sooner than in IVF embryos as 26% of the four-cell stage embryos displayed such organization. Interestingly, in SCNT embryos derived from MG132-treated oocytes, the appearance of condensed heterochromatin blocks was delayed when compared to control SCNT embryos: we observed such organization in only 7.5% four-cell stage embryos. This shows that SCNT embryos derived from MG132-treated oocytes displayed a pattern of heterochromatin organization more similar to the one observed in IVF embryos. It suggests that the PCC induced by MG132 treatment enabled the reorganization of the chromatin with a more appropriate timing potentially leading to better reprogramming.
It remains uncertain whether PCC is necessary for the successful reprogramming of an introduced donor nucleus in cattle, and “PCC or not PCC” is still a matter of controversy. Our findings suggest that treatment of the recipient oocytes, with MG132 inducing PCC in all SCNT embryos results and could improve reprogramming following nuclear transfer, in terms of blastocyst quality and CBX1 distribution. Those results are in agreement with some other reports where different strategies have been designed to enhance MPF activity, and hence, the PCC rate. For example, MG132 treatment in mouse SCNT embryos resulted in increased levels of blastocyst formation (Gao et al., 2005; Yu et al., 2005). In pigs, brief exposure to MG132 during the first 2 h after NT resulted in improved embryonic development (Whitworth et al., 2009; You et al., 2010). Interestingly, the first successful cloning of rats was achieved by briefly exposing MII oocytes to MG132 in order to prevent spontaneous activation (Zhou et al., 2003). In ovine oocytes, treatment with caffeine promoted an increased in PCC and caused larger total cell numbers in blastocyst stage embryos (Lee and Campbell, 2006). Alternatively the pretreatment of donor cells with mitotic cell extracts facilitating PCC was associated with improved reprogramming and SCNT embryo development (Sullivan et al., 2004).
Based on all these results, it appears that PCC may be beneficial to SCNT, probably because it enables better access for the oocyte factors involved in the remodeling and reprogramming of the somatic nucleus and also allows a rapid and favorable adequation between the oocyte cytoplasm and the somatic foreign nucleus. This implies that the morphological reorganization of the transferred nucleus could play an important role in the reprogramming process and subsequent development of SCNT embryos. It also suggests that there is a critical window of time just after donor nucleus introduction for this reorganization.
Using this protocol with or without MG132, we were able to reconstruct embryos with two different chromatin behaviors: PCC or not. We thus have an appropriate tool that will enable us to further investigate differences in the molecular/biochemical events involved in chromatin remodeling after nuclear transfer and to understand the mechanisms involved.
Footnotes
Acknowledgments
We thank Pascal Bodinier and Christophe Audouard for their technical help during this work. We also thank Renaud Fleurot and Dr. Pierre Adenot for their advices and the use of the INRA MIMA2 platform. We are grateful to Dr. Yvan Heyman for the IVM/IVF facility, the staff of the UCEA Experimental farm in Bressonsvilliers for helping with oocyte collection and embryo transfers. This work was supported by INRA (Institut National de la Recherche Agronomique) and UNCEIA (Union Nationale des Coopératives agricole d'Elevage et d'Insémination Animale).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.