
Abstract
Abstract
Tetraploid embryos are a useful model for postimplantation development of polyploidy cells, and tetraploid cells are an advantage in studies for chimeras yielding offspring completely derived from embryo stem cells or induced pluripotent cells. This study was designed to investigate the effects of cytochalasin B (CB) on bovine oocyte meiosis, and to induce the formation of diploid oocytes and tetraploid embryos. The results showed that: (1) incubation of oocytes in CB at ≥2.0 μg/mL concentrations for 24 h significantly decreased oocyte maturation and the matured oocytes' haploid composition. Over 50% of the CB-treated oocytes did not expel PB1 (non-PB1), and most of the non-PB1 oocytes contained 2n (60) chromosomes. (2) Pretreatment of oocytes with CB at concentrations of 7.5 and 15 μg/mL for 10 h significantly decreased oocyte maturation. Posttreatment of oocytes with CB resulted in most of the oocytes containing 2n chromosomes. (3) The parthenogenetic blastocysts (25–28%) derived from the non-PB1 oocytes of posttreatment group was significantly higher than that from pretreatment, whole period treatment, and the control oocytes (12–16%). (4) Cytogenetic analysis of the embryos derived from CB-treated non-PB1 oocytes resulted in 74% of the one-cell stage embryos being 4n = 120 chromosomes, 82% of two-cell stage embryos contained 4n chromosomes in each blastomere, and 75% of the blastocysts were tetraploidy (4n = 120). (6) The stopped uncleaved one-cell embryos showed an amazing phenomenon of over 15% of them containing extra chromosomes, which suggested multiple DNA duplication occurred within 40 h after activation. In conclusion, CB inhibits PB1 extrusion, disfigures spindle structure, decreases oocyte maturation, and results in formation of diploid (2n or 4c) oocytes. The diploid oocytes resulted in a higher development of tetraploid embryos, which would be a unique approach for the production of tetraploid embryos in bovine.
Introduction
Disruption or inhibition of MF assembly by cytochalasin B (CB) or cytochalasin D (CD) during meiosis did not prevent oocyte GVBD, but resulted in failure of PB extrusion in the mouse (Kubiak et al., 1991; Wassarman et al., 1976; Zhu et al., 2003) and pig (Somfai et al., 2006). When mouse oocytes were exposed to CD, segregation of homologous chromosomes occurred, resulting in two meiotic spindles that merged into one single spindle in the oocytes (Kubiak et al., 1991). In porcine, CB caused similar disturbances as occurred in mouse segregation of homologous chromosomes (Somfai et al., 2006). A similar phenomenon was observed in nicotine-treated oocytes in bovine and resulted in the occurrence of diploid oocytes (Liu et al., 2007).
Tetraploid embryos have proven to be a useful model for postimplantation development of polyploidy cells (Kaufman and Webb, 1990). The tetraploid component readily contributes to extra-embryonic tissues but rarely to the fetus itself (James et al., 1995). In a diploid and tetraploid chimera, the tetraploid cells display a nonrandom distribution in the developing conceptus (Everett and West, 1998). Tetraploid cells are an advantage in studies, for chimeras yield offspring completely derived from embryo stem cells (ESs) (Nagy et al., 1993; Ueda et al., 1995; Wang et al., 1997). In very recent studies, tetraploid blastocysts were successfully used as intermediate recipients for the production of viable pups derived from induced pluripotent cells (iPS) in mice (Kang et al., 2009; Zhao et al., 2009).
Tetraploid embryos can result from inhibition of cleavage at an early stage (Snow, 1973), blastomere fusion (Eglitis, 1980; O'Neill et al., 1990; Spindle, 1981), or electrofusion (Curnow et al., 2000; Iwasaki et al., 1989; Ozil and Modlinski, 1986) in mice, rabbits, pigs, and cattle. The formation of diploid oocytes makes it possible to produce parthenogenetic tetraploid embryos. This study attempted to produce tetraploid embryos by the inhibition of both PB1 and PB2 during oocyte maturation in vitro. Dynamic changes of meiotic spindle formation, chromosomal composition of the resultant oocytes, subsequent parthenogenetic embryo development, and their cytogenetic composition were investigated, respectively, after treatment of bovine oocytes with different concentrations of cytochalasin B.
Materials and Methods
Chemicals
Unless otherwise noted, all chemicals used in this study were purchased from Sigma Chemical Co. (St. Louis, MO). Each experiment included at least three replicates.
Maturation of oocytes in vitro (IVM)
Maturation of bovine oocytes was in accordance to the procedure of Li et al. (2004). Briefly, bovine cumulus oocyte complexes (COC) were aspirated from 3–8-mm diameter follicles on ovaries collected from a local abattoir. Only COC with a compact and a homogenous ooplasm were selected. The oocyte maturation medium was TCM 199 with Earle's salts, L-glutamine, and sodium bicarbonate (Gibco Inc., Grand Island, NY), supplemented with 10% fetal bovine serum (FBS) (HyClone, Logan, UT), 25 μg/mL gentamycin, 0.01 U/mL FSH (NIH-FSH-S17), 0.01 U/mL LH (USDA-bLH-6). The oocytes were cultured in four-well plates (30–50 oocytes/per well) in 0.5 mL maturation medium in the presence of CB at different concentrations and for different times according to the experimental design.
Parthenogenetic activation
After maturation, the oocytes with PB1 (PB1 oocytes) and without PB1 (non-PB1 oocytes) were activated by 5 μM ionomycin for 5 min, then treated with 10 μg/mL cycloheximide (CHX) and 5 μg/mL CB in CR1aa plus 3% FBS for 5 h at 39°C in 5% CO2 in air. The addition of CB in CHX medium was to inhibit PB1 and PB2 extrusion and produce tetraploid embryos. After activation, the oocytes were cultured in 30 μL droplets of CR1aa + 0.3% bovine serum albumin (BSA) for 40 h. The cleaved embryos were transferred to CR1aa + 4% FBS medium onto a monolayer of bovine cumulus cells and incubated in 5% CO2 in air at 39°C for 7 days.
Chromosomal analysis and cell number count
One- and two-cell embryos, and day 7 blastocysts were prepared, respectively, and then examined for their cytogenetic composition and the number of nuclei contained within each embryo (Li et al., 2004, 2005a). One- and two-cell embryos were cultured in 0.05 μg/mL of colcemid (Gibco) medium for around 20 h, whereas the blastocysts were within the colcemid medium for 5 h. The embryos were then treated in 1% trisodium citrate for 10–15 min, and transferred onto a clean glass slide and fixed. Slides were stained with 1% Giemsa for 10 min. Chromosomes were examined at 1000 × under oil, and the chromosome composition was determined for each embryo. Images were captured by digital camera with PIXERA Viewfinder Program (Pixera Corporation, San Jose, CA) under a Nikon microscope. Embryos that did not show an interpretable metaphase spread due to gross overspreading or clumped chromosomes were not classified. Simultaneously, blastocyst cell number was counted.
Fluorescent immunostaining of oocytes
Microtubule immunofluorescent staining was described in our previous reports (Li et al., 2005b; Liu et al., 2007). Briefly, oocytes were fixed and treated with a mouse monoclonal antibody against α-tubulin (Sigma, T-5168). Then they were stained in fluorescein isothiocyanate-labeled goat-antimouse Ig G (Southern Biotechnology Associate, Inc., Birmingham, AL, Cat No. 1030-02). Chromatin was stained with 10 μg/mL of propidium iodide. The samples were examined under a Zeiss epifluenent microscope (Carl Zeiss Optical, Inc., Chester, VA). Images were captured by digital camera with PIXERA Viewfinder Program (Pixera Corporation).
Oocytes maturing in 5.0 μg/mL CB-containing medium for 16, 20, 22, and 24 h were denuded, respectively, and then fixed for immunofluorescent staining. Simultaneously, the same stage control counterparts were treated.
Experimental design
Experiment 1 examined the effects of CB on oocyte maturation and chromosomal composition after incubation in CB at 0.1, 0.2, 0.5, 1.0, 2.0, 5.0, 7.5, and 10.0 μg/mL concentrations for 24 h, respectively. The matured oocytes were examined for the presence of the PB1. Both PB1 oocytes and non-PB1 oocytes were prepared for chromosome analysis.
Experiment 2 tested the potential development of CB-incubated oocytes. Oocytes with or without PB1 were parthenogenetically activated. The resultant blastocysts were analyzed for their chromosomal composition.
Experiment 3 was designed to determine whether pretreatment of oocytes with CB affected their maturation and subsequent embryo development. Oocytes were treated with CB at 7.5 and 15 μg/mL-containing media for 6, 8, and 10 h, respectively. The oocytes were intensively washed and then incubated in maturation medium to 25 h. Both PB1 oocytes and non-PB1 oocytes were activated and cultured in CR1aa for 7 days.
Experiment 4, data from Experiment 3 showed that pretreatment of oocytes with CB negative affected oocyte maturation and subsequent embryo development. In Experiment 4, COC were cultured for the first 10 h and then transferred to 1.0, 3.0, 5.0, and 7.5 μg/mL CB-containing medium and cultured to 24 h. The oocyte maturation and their subsequent development were examined.
Table 1 summarized the experimental design described above.
IVM, in vitro maturation.
Statistical analysis
Data were pooled from at least three replicates per group for the in vitro maturation and in vitro development studies. Differences in blastocyst cell numbers between groups were analyzed by one-way analysis of variance (ANOVA). Differences in rates of blastocyst development and chromosome composition between experimental groups were analyzed using Student's t-test. A P-value of < 0.05 was considered statistically significant.
Results
CB affected oocyte maturation and haploid composition in a dose-dependent manner
Oocytes incubated in CB at 0.1, 0.2, 0.5, and 1.0 μg/mL concentrations resulted in similar maturation rates when compared to the control (83–88%), but significantly decreased in the 2.0 μg/mL (44.8%) and 5.0 μg/mL (29%) groups. None of the oocytes matured in 7.5 and 10.0 μg/mL groups expelled PB1 (Table 2).
Values with different superscripts within each column are significantly different. a,b and b,c: p < 0.05; a,c: p < 0.01. Each treatment was repeated at least for three times.
Analysis of the PB1 oocytes showed that 88 to 95% oocytes contained a haploid composition (n = 30) when the oocytes were incubated in 0.1, 0.2, 0.5, and 1.0 μg/mL CB-containing media for 24 h. Significantly lower (74%, p < 0.05) haploid complements were observed when CB concentration increased to 2.0 μg/mL, and only around 50% of the oocytes were haploid in the 5.0 μg/mL treatment group (Table 2).
As shown in Table 2, more than half of the oocytes incubated at or over 2.0 μg/mL CB media failed to expel PB1, and none of the oocytes showed a PB1 extrusion in 7.5 and 10.0 μg/mL CB groups (Table 3). Analyses of these non-PB1 oocytes showed that over 50% of the oocytes contained diploid (2n = 60) chromosomes either in two 30-chromosome plates (Fig. 1D–G) or in a 60-chromosome plate (Fig. 1H–L).

Chromosomal state after the oocytes incubated in CB-containing media.
Values with different superscripts within each column are significantly different. a,b and b,c: P < 0.05; a,c: p < 0.01.
Fluorescent immunostatining of oocytes maturing for 16, 20, 22, and 24 h showed that CB caused the disfiguration of the spindle microtubules compared to the controls (Fig. 2A–E). The spindles were noticeably expanded with irregular shapes (Fig. 2F). CB did not inhibit homologous chromosomes separation but stopped the movement from the central to the cortical area, and arrested at the AI stage. The arrested two chromosome groups formed two spindles (Fig. 2G–J), which occurred as early as 16 h after IVM. A large spindle that was larger than a normal spindle (Fig. 2K–L) appeared at 20 h of IVM. The number of two-spindle oocytes reached the highest incidence at 20 h of IVM and then decreased from IVM 22 to 24 h. The number of one large-spindle oocytes began to appear at 20 h and the incidence reached the highest from IVM 22 to 24 h. The two-spindle and the one large-spindle oocytes correspond to the 30-chromosome-plate and the 60-chromosome-plate in Figure 1, respectively, and both types were diploid.
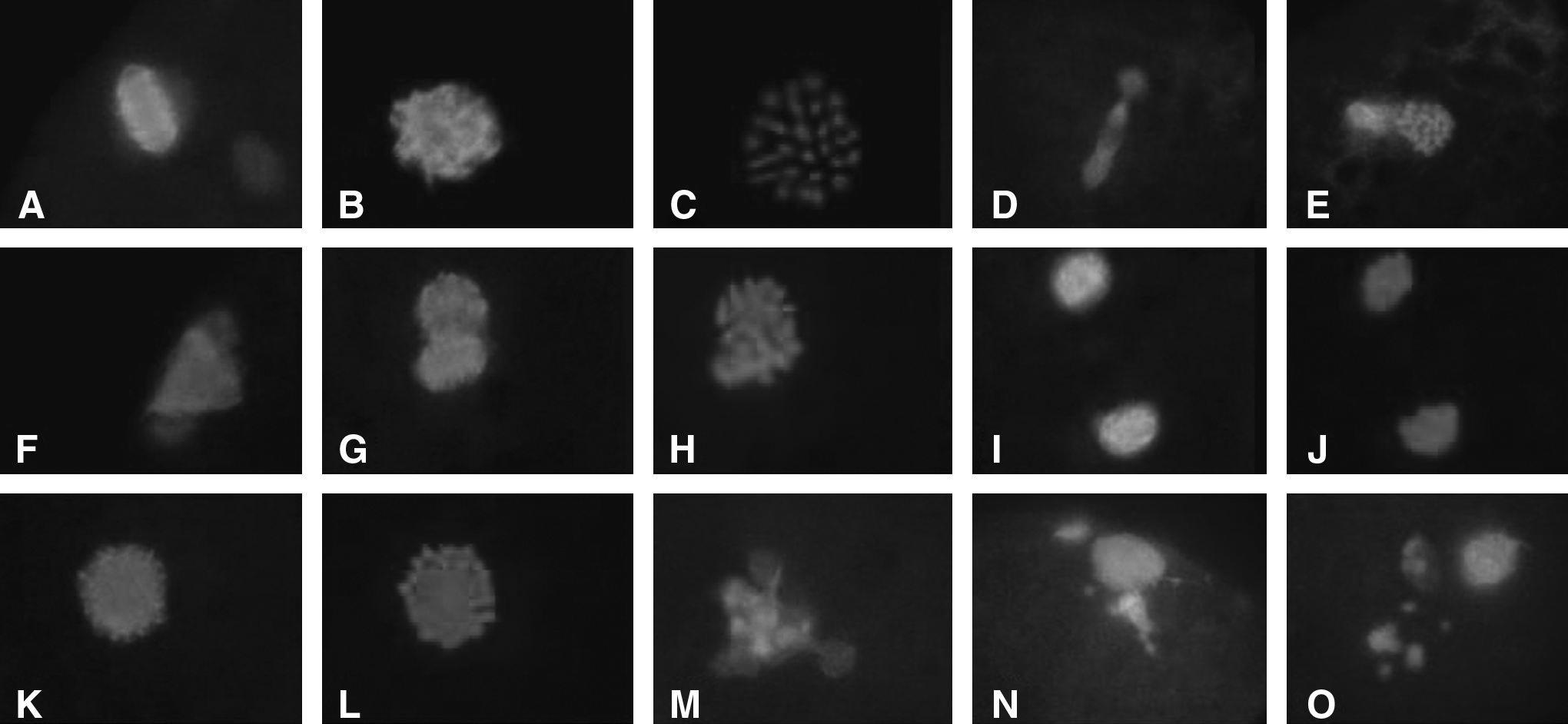
Meiotic spindle morphology of oocytes treated by CB. Oocytes at MI
Tetraploid embryo formation after activation of oocytes treated with CB
To test the developmental potential of CB-treated oocytes, both PB1 and non-PB1 oocytes were parthenogenetically activated. The PB1 oocytes derived from the 0.1 μg/mL group resulted in similar blastocyst development compared to the control, wherea the embryo development in 0.5 and 1.0 μg/mL groups significantly decreased. Non-PB1 oocytes developed to blastocyst stage with similar proportions to the 0.5 and 1.0 μg/mL yielded PB1 oocytes (Table 4). The blastocyst cell number in CB at and over 0.5 μg/mL was significantly lower than that from the control and 0.1 μg/mL groups (Table 4).
Values with different superscripts within each column are significantly different. a,b and b,c: p < 0.05; a,c: p < 0.01.
Chromosomal analyses showed that the majority of the blastocysts derived from PB1 oocytes were diploid. However, almost all of the blastocysts derived from CB-treated non-PB1 oocytes were tetraploid (Table 4).
Pretreatment of oocytes with CB affect oocyte maturation and embryo development in a time-dependent manner
Pretreatment of oocytes with CB at 7.5 and 15 μg/mL for 6 h resulted in similar maturation rates (around 80%) to the controls (83%). The maturation rate (65.9%) significantly decreased when the oocytes were pretreated with 15 μg/mL CB for 8 h. Prolonging CB pretreatment to 10 h in both 7.5 and 15 μg/mL groups resulted in significantly lower oocyte maturation rates (60%). After activation, there were no differences in cleavage, blastocyst rate, and blastocyst cell number between 6-h pretreated oocytes and the control. In the 8-h pretreated oocytes, 15 μg/mL CB caused significantly lower blastocyst development compared to the 7.5 μg/mL group and the control. Pretreatment to 10 h caused significantly lower embryo development and blastocyst cell number (Table 5).
Values with different superscripts within each column are significantly different. a,b and b,c: p < 0.05; a,c: p < 0.01.
Posttreatment of oocytes with CB improved embryo development
After maturation for 10 h, the oocytes were then incubated in CB-containing medium to 24 h. The maturation rates dramatically decreased (p < .05, or p < 0.01) in all experimental groups, even at 1 μg/mL treatment (69.6%) when compared to the control (84.9%). About 60 to 90% of the oocytes did not expel PB1 after treatment with 3.0, 5.0, and 7.5 μg/mL CB, which were significantly higher than the control (Table 6).
The oocytes were activated by ionomycin and CHX + CB.
Values with different superscripts within each column are significantly different. a,b and b,c: p < 0.05; a,c: p < 0.01.
After parthenogenetic activation, the cleavage rates were similar among the CB-treated groups and the control in both PB1 oocytes and non-PB1 oocytes. The blastocyst development, however, was significantly higher in CB-treated (25–28%) than the control oocytes (15%) (Table 6).
Chromosomal analysis showed that approximately 60% of the blastocysts derived from PB1 oocytes in both CB-treated and control groups were diploid. The non-PB1 oocytes treated by CB at 3.0, 5.0, and 7.5 μg/mL resulted in 73, 75, and 76% tetraploid (4n = 120) blastocysts, respectively, and none of the embryos were diploid (Table 7). The blastocyst cell number of the CB-treated embryos was significantly lower than the controls.
Including 3n + 4n, 2n + 3n + 4n, and 4n + 6n + 8n embryos, respectively. pb + : oocytes with PB1; pb-: non-PB1 oocytes. Values with different superscripts within each column are significantly different (p < 0.05).
To better understand the derivation of 4n chromosomes in blastocysts we examined the cytogenetic composition of very early developing embryos such as one-, and two-cell embryos from posttreated oocytes. The results showed that 74% (31/42) of the one-cell embryos were 4n = 120, and 12% (5/42) were in a 2n state. Eighty-two percent (95/116) blastomeres of the two-cell embryos contained 4n chromosomes, and about 3% (4/116) of the embryos were a 2n + 4n chimera.
Tables 4, 5, and 6 showed that high proportion of the CB-treated oocytes did not cleave, and stopped at the one-cell stage 40 h after activation treatment. In the control group, over 80% of the uncleaved embryos stopped at one PN and two PN. However, around 50 to 60% of the one-cell stopped embryos derived from CB-treated oocytes were at one PN, two PN, and three to six PN; 7 to 11% embryos stopped at 4n metaphases (Table 8); and about 15% of the one-cell embryos contained multiple chromosome (over 4n) composition, which were difficult to count as shown in Figure 3D–F.
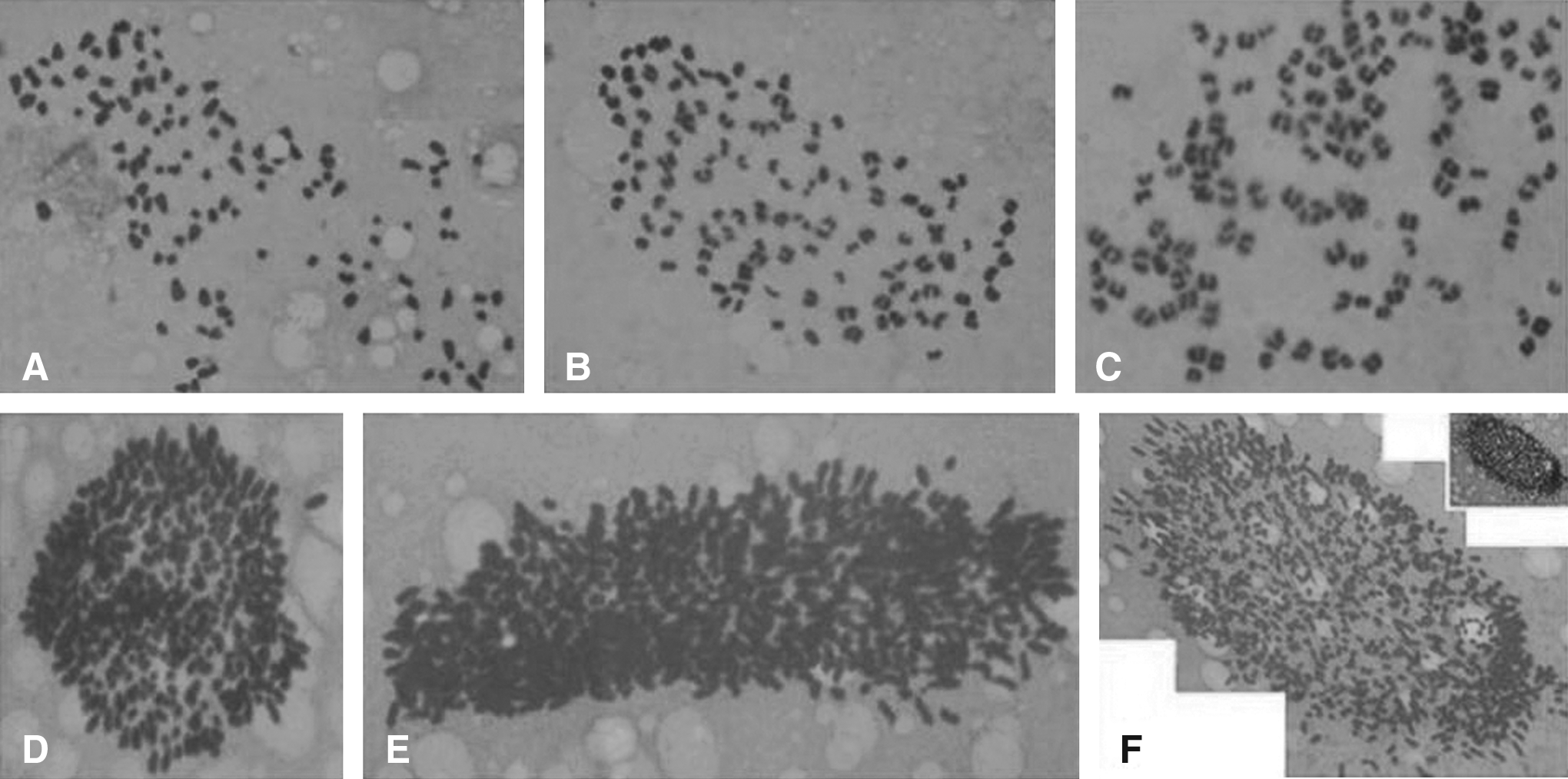
Chromosomal compositions of the blastocysts.
Including chromatin fragmented, degenerated, and dead. Values with different superscripts within each column are significantly different (p < 0.05).
Discussion
Microfilament, a member of the cytoskeleton frame work, takes part in many cytokinetic events and is very important in oocyte maturation and fertilization (Connors et al., 1998; Dimaggio et al., 1997; Maro et al., 1984; Schatten et al., 1986; Sun and Schatten, 2006; Zhu et al., 2003). In bovine, our previous study showed that MF are distributed in the cortex area beneath the oocyte membrane at the GV, GVBD, pre-MI, and MI stages and did not appear in the chromatin area until the early anaphase stage. MF become apparent from AI to the TI stage. As the phenomenon occurs in spindle MT, spindle MF move toward one of the two rows of chromatin, which is destined to promote PB1. During PB1 emission, almost all of MF are expelled along with PB1 chromatin, which also occurs during meiosis II (Li et al., 2005b). In the present study, CB treatment did not affect the homologous chromosomes segregation, but the spindle MT was disfigured and prohibited the movement of chromosomes from the central to the cortical area, and consequently, inhibited PB1 extrusion.
Inhibitors of microfilament assembly and aggregation such as CB, cytochalasin D, and latrunculins are usually used in cell cytoskeleton studies, especially microfilament formation and architecture. In the mouse, treatment of oocytes with CB resulted in the arrest of mouse oocytes at the MI stage without any effect on GVBD, chromatin condensation, and spindle formation (Wassarman et al., 1976), but disrupted microfilaments and inhibited spindle rotation and cytoplasmic division and polar body extrusion (Zhu et al., 2003). In the pig, around 78 to 91% of the oocytes were arrested at the MI stage when oocytes were cultured in CB at concentrations of 1, 3, and 5 μg/mL for 44 h (Somfai et al., 2006), which suggested that CB at 1 μg/mL concentration is enough to inhibit PB1 extrusion. In bovine, CB at or lower than 1.0 μg/mL did not affect PB1 extrusion, whereas significantly decreased maturation rates were observed with the increase of CB to ≥2.0 μg/mL concentrations (Table 2). These results suggested that the sensitivity of bovine and porcine oocytes to CB be different and species specific.
In bovine, the results in our study indicated that CB at or lower than 1.0 μg/mL concentrations did not affect oocyte haploid. Increasing CB concentration to 2.0 μg/mL not only caused lower oocyte maturation rates but also induced significantly lower oocyte haploid. These data suggest that CB at or over 2.0 μg/mL cause uneven segregation of homogeneous chromosomes and therefore result in the formation of unbalanced chromosomal numbers. The alterations in chromosome numbers can also be due to side effects of long-term CB treatment, which then may lead to abnormal segregation of chromosomes during homogeneous chromosomes segregation during PB extrusion followed by irregular distribution of chromosomes. Analyses of the non-PB1 oocytes derived from CB treatment showed that over 50% contained 60 (4c) chromosomes, either in a single 60-chromosome plate or in two 30-chromosome plates (Fig. 1E–L and Table 3).
Pretreatment of oocytes with CB affected oocyte maturation in a time-dependent manner, and the effect is irreversible. Posttreatment of CB on maturing oocytes at the pre-MI and MI stage caused a significantly decrease in PB1 extrusion even at a 1.0 μg/mL concentration. The majority of oocytes did not expel PB1 and indicates that maturing oocytes at later stages were more sensitive to CB than very early stage oocytes (e.g., GV stage). Both pre- and posttreatments of CB induced at high proportions of oocytes contained 60 or 120 chromosomes (2n = 60 or 4c = 120). Immunostaining also showed that CB induced two groups of anaphase chromatin to form two spindles. The two distinct spindles then move toward each other and merged into a large spindle, which resulted in the formation of diploid oocytes. These results suggest that oocytes incubate in CB-containing media either in the pre-, post-, or whole maturation period inhibit PB1 extrusion and result in the formation of diploid oocytes (2n = 60 or 4c = 120). The similar phenomena were observed in CB-treated porcine oocytes (Somfai et al., 2006) and nicotine-treated bovine oocytes (Liu et al., 2007). The mechanism of this phenomenon needs further clarification.
One of the major aims of this study was to assess the development of CB-treated oocytes. Oocytes pretreated, posttreated, or incubated in CB-containing media for the whole maturation period (IVM for 24 h), and PB1 oocytes and non-PB1 oocytes were parthenogenetically activated. Beyond our anticipation, the highest blastocyst development (25–28%) from both PB1 oocytes and non-PB1 oocytes was obtained from the posttreated oocytes when compared to pretreatment and the control groups. This leads us to believe that posttreated oocytes with CB improve embryo development. The mechanism(s) involved in this phenomenon is not clear and warrants further investigation. Chromosomal analyses showed that blastocysts derived from the CB-treated non-PB1 oocytes were tetraploid. The formation of tetraploid embryos is no doubt from the PB1 and PB2 prohibited oocytes. When oocytes at pre-MI or MI are treated with CB it inhibits the extrusion of PB1 and results in the formation of diploid oocytes (2n, or 4c = 60). During parthenogenetic activation the addition of CB in CHX medium further inhibited the PB2 extrusion, and the tetraploid embryos formed. The present results indicate that the resultant 2n or 4c oocytes have high tetraploid embryo development, which could provide a practical method for the production of tetraploid embryos in the bovine. We also observed that, except for some blastomeres that contained pronuclei, all one-cell and two-cell embryo blastomeres were tetraploid (Table 7). This indicates that 4n nuclei begin to form as early as one-cell and two-cell embryo stages, and that 4n nuclei progress to the blastocyst stage.
In the CB-treated groups, a high proportion of oocytes did not cleave and stopped at the one-cell stage 40 h after activation treatment. Routinely, these uncleaved one-cell embryos are discarded. Consequently, nothing is known on what happens in these activated but uncleaved embryos. We collected and cytogenetically analyzed these uncleaved embryos. In the control group, over 80% of the embryos stopped at one PN and two PN stages. A variety of nuclear state, however, was observed in the CB-treated uncleaved embryos, including three to six PNs, 4n, and over 4n metaphases. It was to our surprise that over 15% of the embryos contained extra number of chromosomes (some reach at least 18n) in single cells. As we have shown in Figure 3, it is very difficult to count the number of chromosomes in these cells. It is incomprehensible how the cell DNA can replicate so many times within 40 h. What happens and how it happens is somewhat of a mystery at this time. However, we did not observe extra high number of chromosomes in the developing embryos and blastocysts, which suggest that the cleaved blastomeres have lost the ability to quickly replicate their DNA. The mechanisms involved in what we have observed in these embryos most likely involve complex biochemical and physiological pathways and should be further investigated.
In conclusion, cytochalasin B, an inhibitor for microfilament assembly, affected bovine oocyte maturation. CB pretreatment and posttreatment did not improve oocyte maturation but induced formation of diploid (2n or 4c) oocytes. The diploid oocytes had significantly highest tetraploid embryo development, which could become an efficient approach for tetraploid embryo production. Around 15% of the uncleaved stopped one-cell embryos contained extra numbers of chromosomes.
Footnotes
Acknowledgments
We thank Dr. Thomas D. Bunch, Department of Animal, Dairy, and Veterinary Sciences, Utah State University, for helpful comments and critical reading of the manuscript. This work was supported by National High-Tech Research and Development (863) Programme (No.2007AA100505; 2008AA10Z159), and Chang Jiang Scholars Program of Ministry of Education, China, to G.P.L.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.