
Abstract
Abstract
Recently, we have demonstrated the expression of the polycomb group protein Ezh2 in embryonic and adult neural stem cells. Although Ezh2 remained highly expressed when neural stem cells differentiate into oligodendrocyte precursor cells, it is downregulated during the differentiation into neurons or astrocytes. This is in accordance with the differentiation repressive role Ezh2 is thought to play in the maintenance and self-renewal of stem cells. To establish the importance of downregulation of Ezh2 for becoming astrocytes, we have studied the effect of forced Ezh2 expression in postnatal mouse astrocytes. Upon forced expression of this polycomb group protein, cultured astrocytes retracted their cell extensions and became proliferating round/bipolar cells that occasionally formed small neurosphere-like clusters. Analysis of the expression profile of these Ezh2-expressing astrocytes reveal downregulation of typical astrocytic genes, like GFAP and S100, and upregulation of genes that are generally expressed in neural stem cells, like nestin, Sox2, musashi, and CD133. However, these neural stem cell-like cells lack a differentiation potential, indicating that overexpression of Ezh2 alone is insufficient for a complete dedifferentiation.
Introduction
Central players regulating the accessibility for transcription of lineage specific genes during stem cell differentiation are the polycomb group (PcG) proteins. PcG proteins are transcription repressors that function by modulating the chromatin structure. They act in complexes, so-called polycomb repressive complexes (PRC), of which PRC1 and PRC2 are the most prominent ones (for a recent review, see Simon and Kingston, 2009). Besides the histone binding domain RbAp48, the core components of mouse PRC2 are Eed, Suz12, and the methyltransferase Ezh2, which catalyzes histone H3 lysine 27 trimethylation. Upon this H3 lysine 27 trimethylation, PRC1 is recruited, which contains the ubiquitin ligase Ring1, an essential component for longterm PcG-mediated repression. The PcG proteins play a critical role in the maintenance of pluripotent embryonic stem (ES) cells. However, it has become clear that they also may be involved in the maintenance and fate choice of multipotent stem cells, like haematopoietic stem cells.
A number of recent articles have demonstrated the involvement of PcG proteins in the maintenance of the multipotent neural stem cells and their fate choice during differentiation (Hirabayashi et al., 2009; Mohn et al., 2008; Sher et al., 2008). We were the first to demonstrate the presence of the PRC2 protein Ezh2 in embryonic and adult NSCs (Sher et al., 2008). In accordance with the role Ezh2 is thought to play in the maintenance and selfrenewal of ES cells and the repression of lineage-determining genes, Ezh2 was downregulated when NSCs differentiated into neurons or astrocytes. Remarkably, Ezh2 remained highly expressed in differentiating oligodendrocytes, suggesting a different function of Ezh2 in this cell lineage (Sher et al., 2008). To further examine the mechanisms of neural fate determination and subsequent stabilisation, we have induced the expression of Ezh2 in postnatal mouse astrocytes and investigated the consequences of this epigenetic modulation for the maintenance of their differentiated state.
Materials and Methods
Cell cultures
Primary murine neural stem cell culture
NSCs were isolated from the telencephalon of C57/BL6 mouse embryos at embryonic day 14 (E14). Briefly, the telencephalon was cut into small pieces at room temperature and after mechanical trituration in ice-cold phosphate-buffered saline; the tissue was incubated with Accutase (Sigma-Aldrich, Zwijndrecht, The Netherlands; http://www.sigmaaldrich.com) for 10 to 15 min at 37°C. After repeated trituration, the cell suspension was passed through a cell strainer (70-mm pore size; BD Falcon, Franklin Lakes, NJ, USA; http://www.bdbiosciences.com) and seeded (1–1.5 million cells) in T25 (Nunc, Roskilde, Denmark; http://www.nuncbrand.com) tissue culture flasks containing proliferation medium, which consisted of Neurobasal medium (Invitrogen, Breda, The Netherlands; http://www.invitrogen.com) supplemented with B27 (2%, Invitrogen), human recombinant epidermal growth factor (EGF) (20 ng/mL, Invitrogen), basic fibroblast growth factor (bFGF) (20 ng/mL; Invitrogen), GlutaMAX (1%; GIBCO, Grand Island, NY, USA; http://www.invitrogen.com), Primocin (100 mg/mL), and heparin (5 mg/mL; Sigma-Aldrich) in a humidified 5% CO2/95% air incubator at 37°C. Within 3–5 days the cells grew as free-floating neurospheres and were dissociated and passaged with the use of Accutase. To induce the differentiation of NSCs into the three neural cell types, neurospheres were exposed to Accutase for 5–10 min and dissociated by triturating. Subsequently, the cells were plated in poly-L-lysine and laminin-coated chamber slides (Nunc), at approximately 15,000 cells per well in Neurobasal medium supplemented with B27 for 5 days at 5% CO2 and 37°C.
Primary murine astrocyte culture
Astrocytes were obtained from mixed glia cultures established from postnatal mouse brains. Brains from postnatal C57/BL6 mice were rapidly isolated, and after removal of meninges the cortices were dissected in icecold medium A (Hanks' balanced salt solution supplemented with 1.5% glucose and 1.5% HEPES). Cortices were dissociated mechanically by trituration in medium A and subsequently by trypsinization for 20 min at 37°C in medium A supplemented with 0.25% trypsin and 1,000 U/mL DNase I. After inhibition of trypsin with medium A containing 0.1 mg/mL trypsin inhibitor (Sigma-Aldrich), 20% fetal calf serum (FCS), and 1000 U/mL DNase I, the cells were centrifuged for 10 min at 800 rpm. Single-cell suspensions were seeded in T75 cm2 culture flasks (Greiner Bio-One, Alphen a/d Rijn, The Netherlands; http://www.gbo.com/en) containing Dulbecco's modified Eagle's medium (DMEM) containing 10% FCS, 1% glutamine, 1% sodium pyrupyruvate, and Primocin. After 7–10 days in vitro, the mixed glial cultures were placed overnight on a shaker (150 rpm) at 37°C, and on the following day the nonadherent glial cells were removed. Next, astrocytic monolayers were carefully separated from the underlying adherent microglia by a mild trypsinization step and plated in new T75 cm2 tissue culture flasks. In the following weeks, the overnight shaking and trypsinization steps were repeated until pure astrocyte cultures (approximately 98%) were obtained.
Primary murine oligodendrocyte culture
The above T75 cm2 flasks containing mixed glial culture were shaken overnight at 37°C. Subsequently, the supernatant-containing detached cells were centrifuged at 500 rpm and the pellet was resuspended in culture medium and plated in uncoated culture dishes for 30 min. Microglia and astrocytes attached to the bottom of the culture dish, whereas oligodendrocyte precursor cells (OPCs) remained floating. In consecutive turns the floating cells were transferred from one culture dish to another until a pure (approximately 95%) OPC culture was obtained. After centrifugation, the primary OPCs were cultured in defined SATO medium (oligodendrocytes specific).
Primary murine cortical neuronal culture
Cultures of cortical neurons were established from postnatal brains. In brief, cortices of C57/BL6 embryonic day 17 embryos were excised and collected in ice-cold Hank's Buffered Salt Solution (HBSS) supplemented with 30% glucose. Meninges were removed, and brain tissue was dissociated by trituration in Neurobasal-B27 medium (supplemented with 0.4% glucose, 2 mM L-glutamine, and 0.01% penicillin/streptomycin) and filtered through a cell strainer. After one washing step, cells were seeded on poly-D-lysine (10 mg/mL)-coated six-well plates and maintained in Neurobasal/B27 medium for 7–10 days in a humidified atmosphere (5% CO2/95% air) at 37°C.
Ezh2-gene transfection
Ezh2-containing plasmid was constructed by cloning the Ezh2 gene in a pCDNA3.1 vector. This plasmid was used for the transient forced expression of Ezh2 in astrocytes. Gene transfections were performed by using an electroporesis-based transfection protocol (Amaxa GmbH, Cologne, Germany; http://amaxa.com) specifically designed for the transfection of mouse astrocytes by Amaxa. By using Accutase, 2–3 million cells were collected from T75 tissue culture flasks and were transfected with 5–7 μg of plasmid DNA containing Ezh2. For control transfections, the same vector containing DNA-encoding green fluorescent protein (eGFP) was used. After transfection, all cells were cultured up to 10 days in a humidified 5% CO2-95% air incubator at 37°C in Dulbecco's modified Eagle's medium (DMEM) containing 10% FCS, 1% glutamine, 1% sodium pyrupyruvate, EGF (20 ng/mL) bFGF (20 ng/mL), and Primocin. After 10 days of culture another boost of overexpression was given using the same procedure. After 15 days of culturing from the day of first transfection samples were collected for qPCR and Western blot analysis.
RT-PCR, primers
The primers used in this study are listed in Table 1.
Western blot
Sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis and Western blot analysis were used to detect Ezh2 and other protein expressions in NSCs preoligodendrocytes (pOLs) astrocytes and neurons. Cell lysates were made in Laemmli buffer and boiled for 5 min. Proteins were separated for 1.5 h on a 10% SDS-polyacrylamide gel electrophoresis apparatus (Bio-Rad, Hercules, CA, USA). Subsequently, proteins were transferred to nitrocellulose membranes by using semidry transfer buffer [25 mM Tris, 150 mM glycine, and 10% (v/v) methanol] and 3 mA/cm2 current for 45–50 min. Nitrocellulose membranes were blocked with Odyssey Blocking Buffer (www.licor.com3% milk in TBS-T for 1 h at room temperature. Membranes were probed with antibodies overnight at 4°C. The antibodies against housekeeping proteins were used for normalization of proteins of intrest expression levels. Membranes were washed four times (5 min each) with TBS-T. Primary antibodies were detected using fluorescent secondary antibodies incubated at room temperature for 1 h. After four washes in TBS-T, protein signals were detected by using Odyssey Infrared Imaging System (LI-COR Biosciences, Lincoln, NE, USA) The intensities of the protein signals were quantified by using ImageJ software (NIH Image, Bethesda, MD, USA).
The following primary antibodies were used for Western blotting: anti-Ezh2 (BD Transduction Laboratories, Sparks, MD, USA; 612666), anti-beta-actin (ab6276; Abcam, Cambridge, MA, USA) anti- proliferation cell nuclear antigen (PCNA), a proliferation marker (Abcam; ab29), anti-EED (Millipore, Bedford, MA, USA; 09-774), and anti-Suz12 (Diagenode, Denville, NJ; CS-029-100). Primary antibodies were detected using the following secondary antibodies: IRDye 680 (LI-COR Biosciences; 926-32222) and IRDye 800CW (LI-COR Biosciences; 926-32213).
Immunohistochemistry
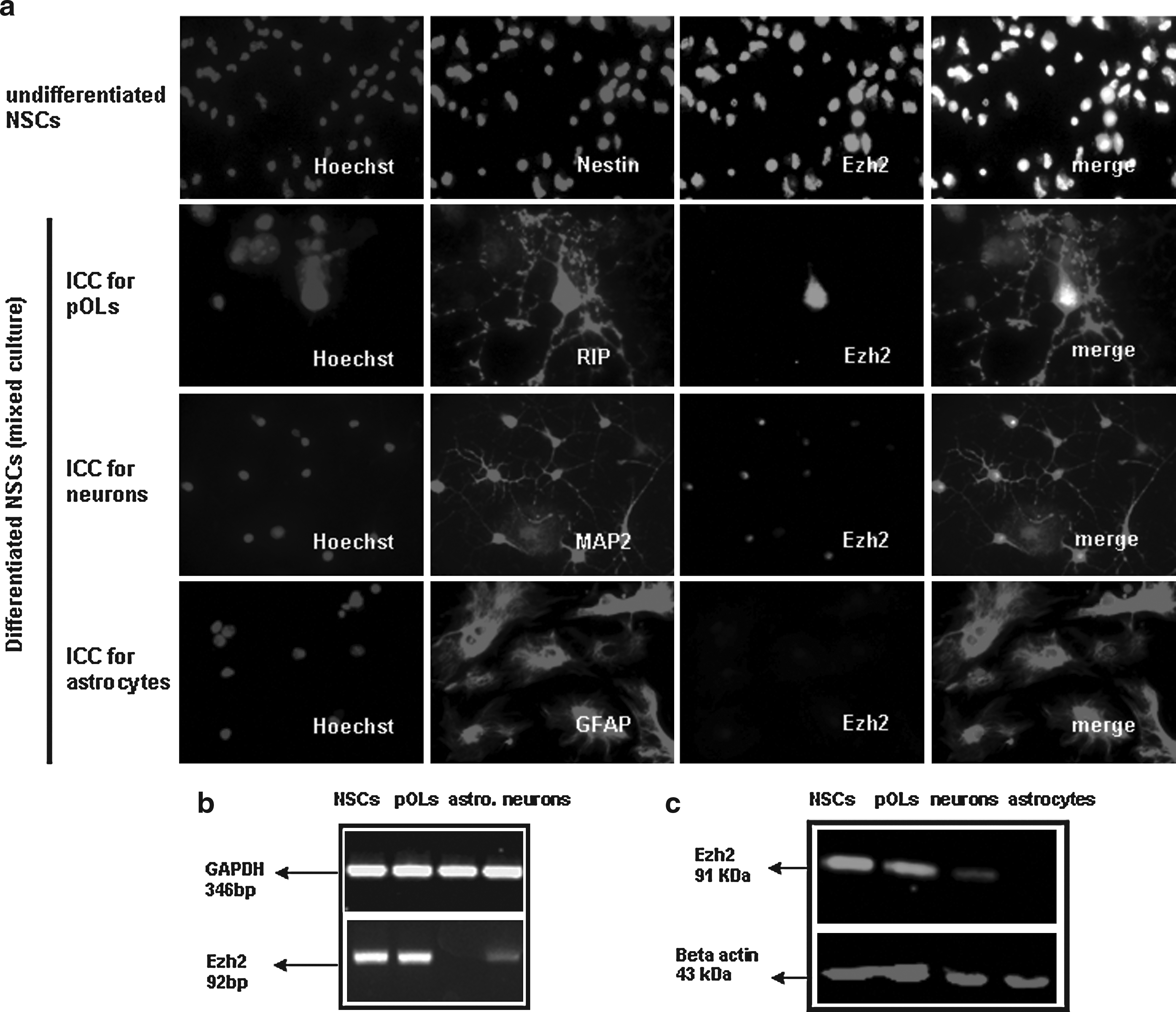
Undifferentiated and differentiated NSCs cultures were fixed with 4% paraformaldehyde and immunostained for markers of various neural cell types. The following antibodies were used: undifferentiated NSCs: antinestin (ab22035, Abcam) to identify neurons, antimicrotubule-associated protein-2 (AB5622, Millipore) astrocytes: antiglial fibrillary acidic protein (MAB3402, Millipore), and pOLs: antioligodendrocytes clone NS-1 (MAB1580, Millipore).
Fluorescein isothiocyanate- or Cy3-conjugated secondary antibodies and Hoechst nuclear staining were used to identify the labeled and unlabeled cells. Zeiss (Axioskope 2) fluorescent microscope equipped with a Leica DFC300FX camera and the Leica microsystem LAS program was used to make pictures.
Results
Immunohistochemical staining shows prominent expression of the polycomb group protein Ezh2 in cultured NSCs isolated from E14 mouse embryo's (Fig. 1a). Differentiation of the NSCs into an astrocytic phenotype is accompanied by a complete downregulation of Ezh2 expression, in contrast to a remnant weak expression upon differentiation into neurons and a high expression in freshly differentiated pOLs.The immunohistochemical data are confirmed by qPCR and Western Blot analysis (Fig. 1b and c).
Forcing expression of Ezh2 in a culture of pure mature astrocytes (Fig. 2a) results in a complete retraction of their astrocytic extensions and a switch to a round cell type within 5 days (Fig. 2b). These cells started to divide when exposed to EGF and FGF2, and occasionally the onset of small neurosphere-like aggregate formation was observed. No significant cell death due to the forced Ezh2 overexpression was detected (data not shown).

Comparing the expression profiles of astrocytes 5 days after the forced overexpression of Ezh2 with control astrocytes (transfected with GFP or noncoding DNA), revealed a significant switch from an astrocytic profile into a more neural stem cell like phenotype: a steep downregulation of typical astrocytic genes, like GFAP and S100b, was accompanied by a clear induction of the expression of nestin, vimentin, Sox2, musashi 1/2, and CD133 (Fig. 3a).
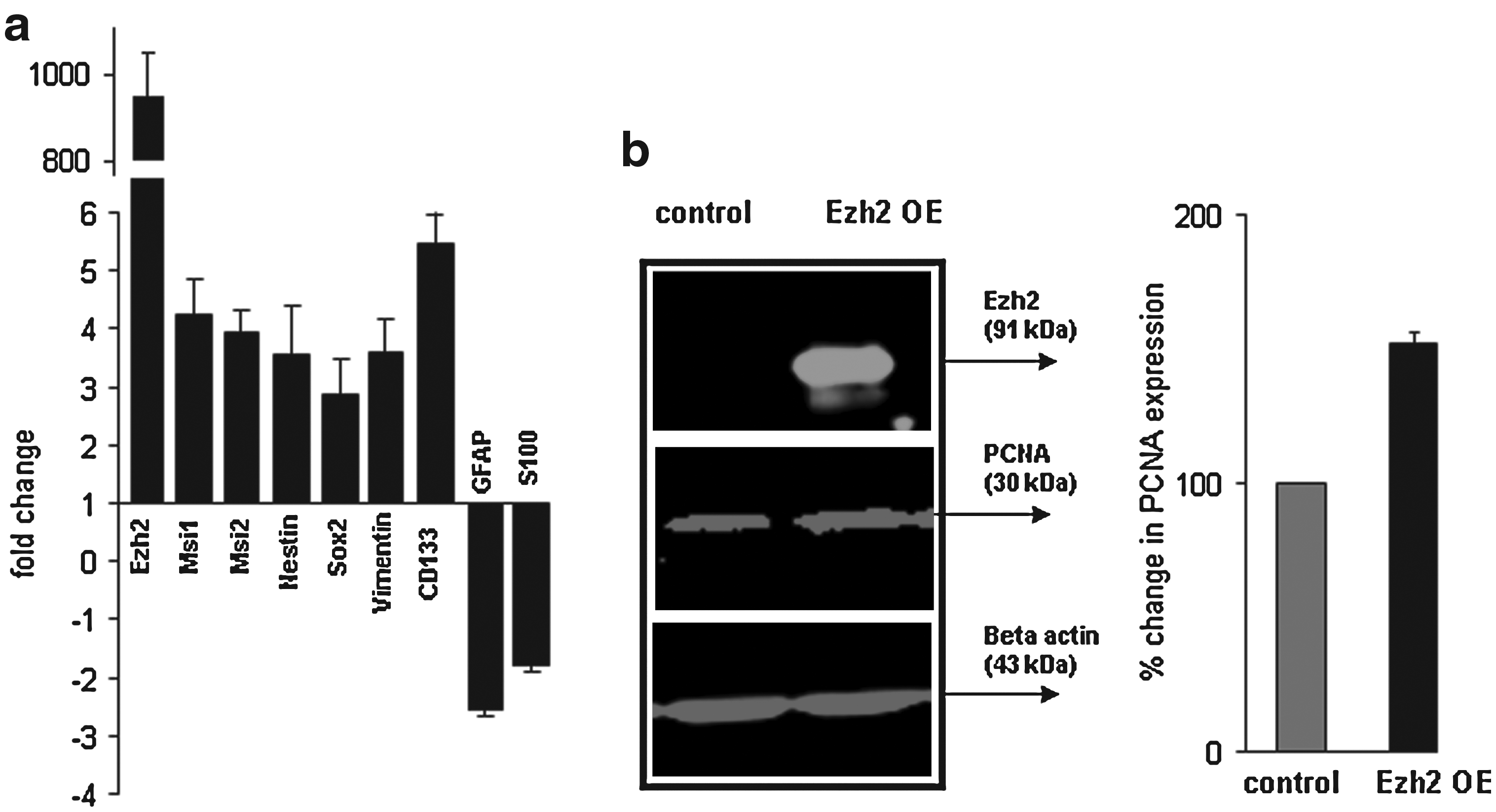
Western Blot analysis of PCNA, a proliferation marker (Fig. 3b), shows an increase of approximately 50% in proliferation activity in the dedifferentiated astrocytes after induced Ezh2-overexpression in medium containing the proliferation factors EGF and FGF2. However, withdrawal of these factors did not induce the differentiation of these neural stem cell-like cells.
The observed incomplete dedifferentiation of astrocytes may have been due to an aberrant composition and function of the PRC2 complex. Western blot analysis was performed in order to check whether the forced expression of Ezh2 in astrocytes and the subsequent dedifferentiation into NSC-like cells was accompanied by an induction of the expression of the other core PRC2 components, Eed2, and Suz12. Our results show that Suz12 (plus an additional isoform) was expressed but not Eed2, indeed suggesting the presence of an incomplete PRC2 complex (Fig. 4).

Forced Ezh2-overexpression in astrocytes induces an incomplete upregulation of the other PRC2 components normally present in NSCs; Western blot analysis shows induced expression of Suz12 but not of Eed2.
Discussion
The differentiation of NSCs into astrocytes, neurons, or oligodendrocytes is a unidirectional process leading to neural cell types of which the stability is epigenetically assured by chromatine modifications. Previously, we described that upon development of astrocytes the repressive epigenetic factor Ezh2 was completely downregulated. In order to study the importance of this downregulation of Ezh2, we induced the expression of Ezh2 in mature astrocytes. This forced expression of Ezh2 indeed resulted in a (partial) dedifferentiation of the astrocytes towards NSCs.
The formation of an astrocyte requires stable repression of nonastrocytic genes (e.g., genes that support a neuronal or oligodendrocytic cell lineage) and the silencing of neural stem cell genes. Our data indicate that forced expression of the PcG protein Ezh2 results in the repression of genes directly or indirectly involved in determining the astrocytic cell lineage. Apart from this astrogenic repression, a concomitant derepression and/or reactivation of typical neural stem cell genes was observed. It is possible that products of astrocytic genes suppress neural stem cell gene transcription; induced repression of these NSC-gene inhibitors by Ezh2 may have led to the derepression of the NSCs genes.
Forced Ezh2 overexpression in astrocytes also resulted in an increase in proliferative activity. This seems to confirm the role of Ezh2 in modulating genes implicated in cell cycle control as demonstrated in various studies on glioblastoma cancer stem cells. Disruption of Ezh2 signaling in glioblastoma stem cells has been shown to strongly impair self-renewal via c-Myc and prevents glioblastoma stem cell maintenance. Forced (over)expression of Ezh2 activated c-Myc transcription and stimulated self-renewal proliferation and (re)activation of stem cell properties (Suva et al., 2009). As demonstrated by Hirabayashi et al. (2009), during the differentiation of NSCs into astrocytes, the cell fate choice into neurons is prevented by Ezh2 repressing the promotor of the proneural gene neurogenin1. This provisionary repression by Ezh2 in the NSCs during astrocytic differentiation is replaced by a more stable repression in mature astrocytes, presumably via DNA methylation, which no longer requires the action of Ezh2. Apparently, this Ezh2 independent stable neurogenin1 silencing was preserved in the Ezh2 overexpressing astrocytes because no neuronal genes were derepressed or reactivated (data not shown).
The partial switch in astrocytes toward a more neural stem cell state may suggest a role for Ezh2 as a kind of regulator of the differences between GFAP-expressing astrocytes that have neural stem cell potential and those that do not: Imura et al. (2003, 2006) have demonstrated that primary astrocyte cultures prepared from postnatal brain contain substantial subpopulations of GFAP-expressing cells that can easily be turned into multipotent neural stem cells simply by removing serum and adding EGF and FGF to the media. Moreover, it is also well known that there are populations of GFAP-expressing radial cells present in the postnatal and adult brain in vivo that are neural stem cells and characterized by expression of LeX/CD15 (Garcia et al., 2004; Imura et al., 2006). It would be interesting to analyze Ezh2 expression in these GFAP expressing progenitors.
The incomplete dedifferentiation of the astrocytes toward a neural stem cell state by forcing singularly the Ezh2 expression raises several interesting questions for future research: what is the action on transcription of Ezh2 alone or within an incomplete PRC2 complex, what would dedifferentiation be like if the other components and an intact PRC2 complex was induced in the astrocytes, and which are the target genes involved?
Footnotes
Acknowledgments
This research was supported by a grant of the Dutch Foundation MS Research, MS 04- 554MS.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.