
Abstract
Abstract
Developmental signaling molecules involved in dorsal patterning of the spinal cord have been identified in vivo; however, studies have not produced specific functional dorsal spinal cord neurons in vitro. We present here differentiation of R1 embryonic stem (ES) cells into GABAergic dorsal spinal cord neurons by sequential treatment with developmental signaling molecules. We found that retinoic acid, Bmp4 altered the specification of neural progenitors and instructed neural fate when applied at distinct stages of development. High concentration of retinoic acid initiated caudal patterning during early differentiation; Bmp4 induced dorsal development. The combination of retinoic acid and different concentration Bmp4 controlled the differing regional progenitors of spinal cord. Low-concentration Bmp4 and high concentration of retinoic acid-treated embryoid bodies resulted in the differentiation of GABAergic neurons. In summary, we demonstrate this simple treatment paradigm produced simple dorsal spinal cord neurons, which could be utilized for developmental and preclinical studies.
Introduction
In vitro differentiation of ES cells into neural cells closely follows genetic programming that is similar to processes employed during embryonic development The differentiation methods typically induce cell aggregation, resulting in embryoid bodies (EBs). Various inducing conditions during embryonic body formation can drastically affect the proportions of differentiating cell types. Neural progenitor differentiation has been shown to be greatly influenced by retinoic acid (RA) (Schlett and Madarasz, 1997). In vivo studies have demonstrated that RA induces differentiation of neural progenitors along the rostral/caudal axis in a concentration-dependent manner (Kim et al., 2009; Ribes et al., 2009). However, the overall effects of RA remain poorly understood, and precise analysis of gene expressional changes due to RA treatment should be useful for establishing proper culture protocols for ES cell differentiation.
The appearance of distinct cell types at defined positions in the spinal cord is dependent on inductive signals that are derived from axial midline cells (Jessell, 2000). Bone morphogenetic protein (BMP)-family members, which play a prominent role in dorsal neural tube cell-type specification, also appear to influence patterning in ventral regions of the neural tube (Busch et al., 2006; Stottmann et al., 2006). Shh is necessary and sufficient in vivo and in vitro to induce differentiation of most ventral neuronal subtypes (Hu and Zhang, 2009; Li et al., 2008). Results imply that several signaling pathways influence the response of progenitor cells to ventrodorsal activity. When neuronal differentiation is initiated in the ventral neural tube, the combined expression of a group of homeodomain proteins have been used to identify five domains of progenitors (Caspary and Anderson, 2003). The ventral expression boundaries of Class I proteins delineate progenitor domains. In contrast, the dorsal expression limits of Class II proteins serve to define progenitor domains. A critical role of Class I and Class II proteins in cell-fate specification is to control activation of specific neuronal subtype determining genes (Helms and Johnson, 2003). These selective interactions between pairs of Class I and Class II proteins function to refine boundaries, which establish discrete patterns of gene expression. The present study analyzed extracellular signals and transcriptional mediators that direct generation of distinct ES cell-derived neuronal subtypes. The present results provided a framework to better understand spinal cord patterning and to fulfill their clinical transplantation.
Results
High-concentration RA initiated caudalization of neural progenitor cells in EBs
RA is thought to be one of the most important extrinsic inductive signals required for position identity specification during in vivo embryonic development (Maden, 2006). To initially determine whether high-concentration RA promoted terminal differentiation of ES cell-derived neural progenitors, 2 and 5 μM concentration of RA were added at different time points or cells were exposed for different culture periods (Supplementary Fig. 1; see online Supplementary Data at www.liebertonline/cell.com). As showed in Figure 1A, the high number of nestin-positive cells in RA-treated EBs compared with untreated EBs. ES cell neural differentiation was promoted by 4 days of exposure to high concentration of RA in EBs (Fig. 1A and Supplementary Fig. 1B). In contrast, Nestin-positive cells were detected between 2 and 5 μM RA treatments, and results further confirmed that 5 μM RA strongly induced neural differentiation of ES cells (Fig. 1B). Subsequently, rostral–caudal identity of neural cells was altered by varying concentrations of RA (Okada et al., 2004). High-RA treatment (>2 μM) suppressed the forebrain marker Otx2, but induced the hindbrain marker Gbx2 and the spinal cord marker Hoxd4, Hoxc4, Hoxc5, and Hoxc6 (Fig. 1C) in a concentration-dependent manner. These results demonstrated that high-concentration RA did not limit differentiation to a neural fate, but also modified caudalization in EBs.
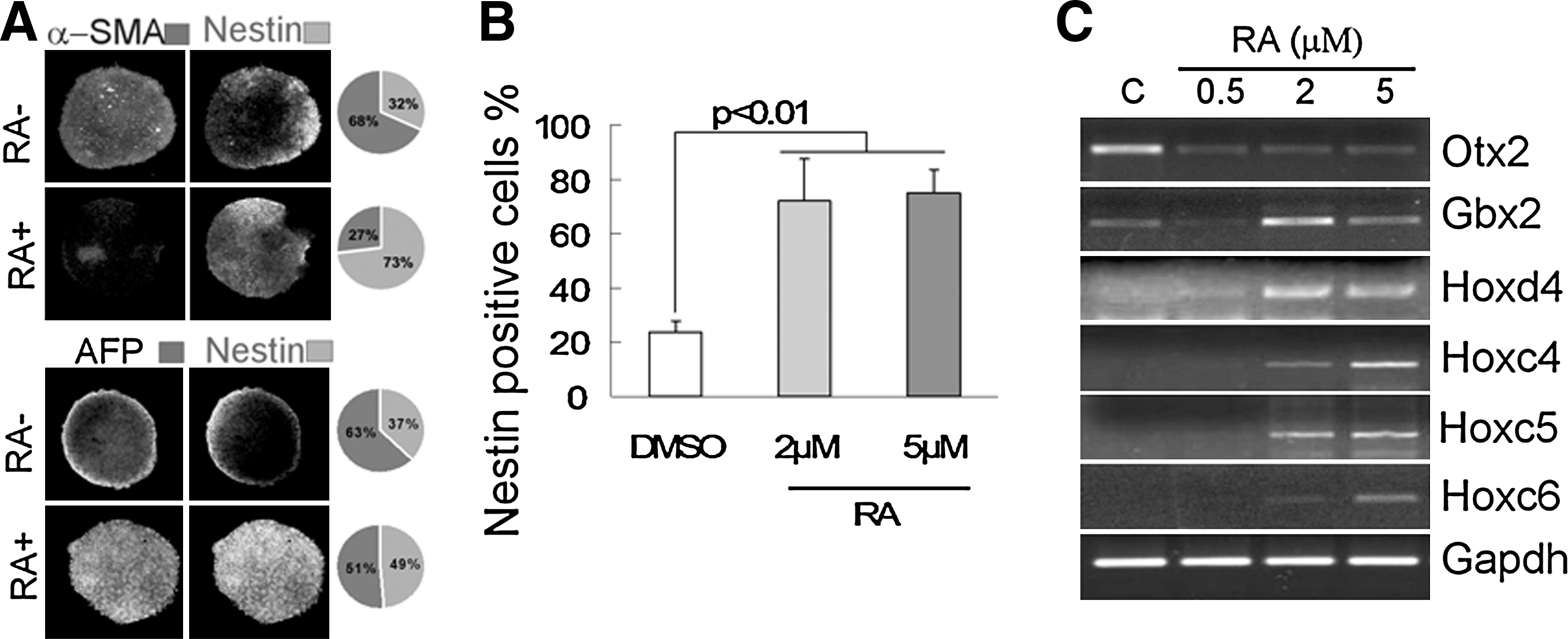
Neural induction of mouse ES by high concentration RA. (
Interestingly, Noggin has been shown to induce neural differentiation of human ES cells (Lamb and Harland, 1995). However, Noggin was not found to increase expression of the neural progenitor marker Pax6 or the caudal gene markers Gbx2 and Hoxc4 through contrast specifically defined programs (Supplementary Fig. 2A–C) in mouse ES cells.
Dorsoventral specification of BMP4 and Shh-induced EBs
Mouse embryonic developmental studies have demonstrated that Bmp4 and Bmp7 are required for initial differentiation of dorsal neural cell types at the neural tube stage. To determine the role of Bmps in EB dorsal–ventral axis formation, EBs were treated with Bmp4 or Bmp7 for 2 days following RA induction. RT-PCR revealed that Bmp4 treatment raised expression levels of class I genes Pax6, Pax7, and Dbx1, and significantly suppressed class II genes Nkx2.2 and Nkx6.1 (Supplementary Fig. 3A), whereas Bmp7 exhibited no obvious effect on dorsoventral gene expression (Supplementary Fig. 3A). Furthermore, studies were designed to determine whether Bmp4 could alter RA-induced effect in EBs. Cell cultures were treated with Bmp4 for 2 days in combination with RA. Caudal gene expression and the number of Nestin-positive cells remained unchanged compared to the control (Supplementary Fig. 3B and C). In contrast to Bmps activity, Shh treatment suppressed class I gene expression and induced expression of class II genes in a dose-dependent manner (Supplementary Fig. 3D). Cyclopamine, an inhibitor of Shh, was used in combination with Shh, which inhibited ventral decision activity of Shh, as determined by RT-PCR analysis (Supplementary Fig. 3D). These results indicated that Bmp4 and Shh treatment influenced dorsoventral positional identity of EBs in vitro.
Induction of dorsal spinal cord neural progenitor differentiation
Progenitor cell identity generated by Bmp4 and Shh was analyzed by immunofluorescence. Results showed that 5 nM Bmp4 significantly decreased the number of Mash1-positive cells. Mash1-positive cells were significantly increased at 0.5 nM, but not at 5 nM, Bmp4 (Fig. 2A and C). Moreover, Bmp4 treatment resulted in the decreased the number of Olig2-positive cells, which was consistent with gene expression patterns for dorsal progenitor cells in the spinal cord. In contrast, Shh treatment suppressed expression of Math1 and increased the number of Ngn2/Mash1- and Olig2-positive cells (Fig. 2B and D). A total of 300 nM Shh increased the number of Olig2/Tuj1-positive cells. However, following 7 days of 30 nM Shh treatment, the number of Olig2-positive cells among the Tuj1-positive neuronal population increased. Consistent with these results, a high proportion of HB9 neurons (Supplementary Fig. 3E) were detected among the Tuj1-positive neurons, which exhibited choline acetyltransferase (ChAT) expression following 12 days of 300 nM Shh treatment (Supplementary Fig. 3E). In contrast, 0.5 nM Shh increased the number of oligodendrocytes (O4+, MBP+) (data not shown).
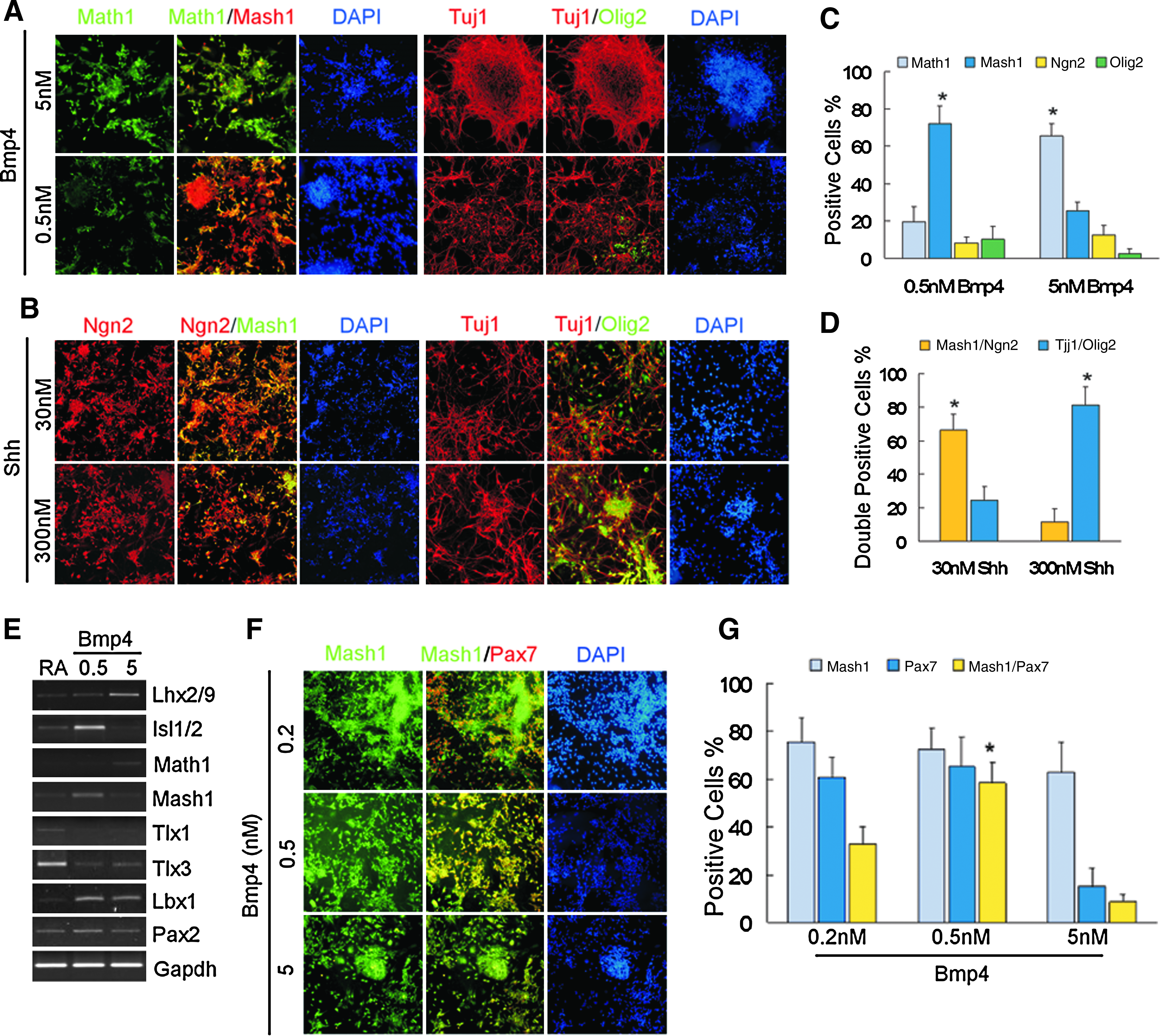
BMP4 and Shh induced differentiation of ES cells. (
Collectively, these results demonstrated that Bmp4 treatment, similar to the in vivo situation, initialized dorsalization of ES cell-derived neural tissues and induced the generation of dorsal spinal cord progenitor cells when combined with RA in vitro. In contrast, Shh promoted the differentiation of a spinal cord ventral progenitor fate, and cells differentiated into motor neurons and oligodendrocyte.
Generation of GABAergic neurons of the dorsal spinal cord
It is well known that progenitor cells expressing Math1 give rise to dI1 neurons, and Mash1-positive progenitors are likely to give rise to dI3–dI5 and dIL neurons during spinal cord development. To further investigate neural cell fate specification and differentiation induced by RA and Bmp4, RT-PCR analysis of transcription factors specific to neural cell fate was performed on day 12. Results showed that 5 nM Bmp4 induced gene expression in the posterior regions of spinal cord (Lhx2/9, Math1) (Fig. 2E), whereas 0.5 nM Bmp4 increased gene expression in anterior regions (Isl1, Mash1, Pax2, and Lbx1) and suppressed Tlx1 and Tlx3 (Fig. 2E). Moreover, 0.5 nM Bmp4 significantly increased the number of Pax7/Mash1-positive cells, compared to 5 nM Bmp4 and RA (Fig. 2F and G). Tlx1 and Tlx2 are postmitotic selector genes, which inhibit GABAergic neuronal cell fate in the embryonic dorsal spinal cord (Cheng et al., 2004). Therefore, transmitter phenotypes of the neural cells were examined. Isolated GABA-positive neurons, and networks of GABA-positive neurons (up to 90%), were observed following RA and 0.5 nM Bmp4 treatment (Fig. 3A). In addition, the Tuj1-positive neurons expressed the synaptic protein synaptophysin (Fig. 3C) and Map2 (data not shown). However, expression of GABAergic neurons was less in RA and 5 nM Bmp4-treated cells (Fig. 3B). Interestingly, Gad65 expression was detected in Mash1-positive progenitor cells earlier than GABA was expressed in Tuj1-positive neurons (Fig. 3D).
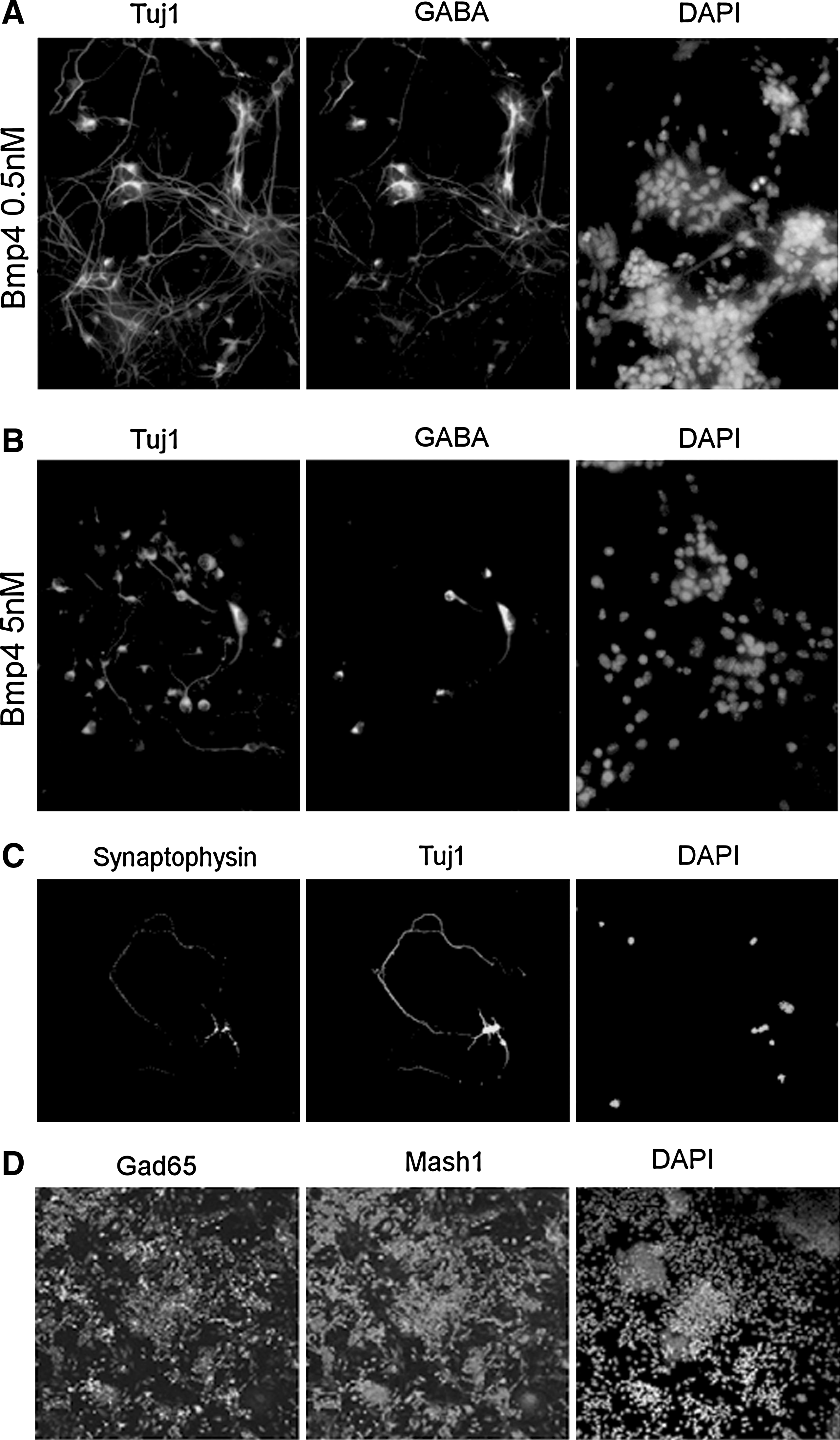
Directed generation of GABAergic neurons. (
Discussion
Previous in vitro and in vivo studies have shown that ES cells are capable of neuronal differentiation. However, systematic derivation of dorsal specific spinal cord neurons from ES cells has not been reported. The present study demonstrated that RA and Bmp4 induced dorsal GABAergic neuronal differentiation from mouse ES cells through the manipulation of timing, concentration, and combination of exogenous factors. The location of cell fate in the CNS depends on two signaling systems that are activated in combination with the more basic program of neural induction in vivo (Melton et al., 2004). These two signaling systems intersect along the rostral–caudal and dorsoventral axes of the neural tube to establish a grid-like set of positional cues. RA is well known as a biologically active form of vitamin A, which plays an important role in patterning of the anteroposterior and dorsoventral axis and is thought to be involved in the generation of the entire anteroposterior CNS. The present study showed that EBs treated with high concentrations of RA expressed genes specific to caudalization (hoxc4, hoxc5, and hox6). In addition, induced expression of more posterior genes required an even higher concentration of RA, which was consistent with physiological concentrations of RA that lead to posterior rhombomere development in vivo.
In contrast to acquisition of rostrocaudal identity, dorsoventral identity was analyzed in terms of expression of transcriptional control of the homeodomain and basic helix-loop-helix (bHLH) proteins. Previous studies had emphasized that Shh and Bmps had emerged as the preeminent inductive signals involved in dorsoventral patterning of the neural tube (Guillemot, 2007; Pierani et al., 1999). The present results demonstrated that high concentration of RA and Bmp4 caused dorsal positioning of the spinal cord, which was partly different from the conclusion reached by Murashov et al. (2005); we have provided additional analyses of different dose of Bmp4 in the cell culture system. Shh induced formation of the ventral position, as determined by gene expression analysis of various classes of transcription factors that specify neural cell fate. Therefore, following acquisition of rostrocaudal identity, addition of Bmp promoted differentiation of dorsal spinal cord progenitor cells, including expression of Mash1 or Math1. Shh induced differentiation of ventral spinal cord progenitor cells, which were Olig2- or Ngn2-positive. In particular, high Shh concentration induced efficient differentiation of HB9-positive motor neurons. According to these results, higher RA concentrations, in combination with Bmp4 or Shh, facilitated EBs to acquire distinct characteristics and different neural fates depending on their spatial patterning function in vitro.
In the spinal cord, the bHLH family transcription factors initiate neurogenesis in neural progenitors. Mash1 appears to mark progenitors that give rise to dI3–dI5 and dILA/B neurons (Helms et al., 2005). The present results indicated that 0.5 nM Bmp induced Mash1 expression, where the majority of cells were also Pax7-positive, as well as expression of progenitor cell markers of dl4 and dlA neurons, such as Isl1, Pax2, and Lbx1. The same conditions inhibited expression of Tlx1/3. Pax2 is specifically required for dorsal spinal cord GABAergic differentiation, and Tlx1/3 has been shown to suppress GABAergic differentiation. The GABA system followed a rostrocaudal gradient of maturation, which initially spread from a ventral–medial to a ventrolateral position, and subsequently faded in ventral areas of the cord while invading dorsal areas (Wilson and Maden, 2005). From E15.5 to P0 GABA-positive fibers are primarily located in dorsal areas of the mouse spinal cord (Allain et al., 2004, 2006) The present studies suggested that in vitro spinal cord GABAergic neurons were derived from proliferating progenitors, which subsequently expressed Mash1 and Pax7, and upon exiting the cell cycle expressed GAD65. These results were similar to mouse in vivo development, because the position of progenitor cells along these two axes is thought to influence cell fate by defining the identity and concentration of inductive signals in vivo.
Future studies are needed to map expression patterns of regional-specific markers observed in ES-derived neural cells to in vivo markers. The combination of high RA concentration-dependent neural induction and Bmp4 signaling was shown to control-specific dorsal neuronal differentiation from ES cells. These results provide a model for spinal cord development research and validate the use of ES cells that carry disease-causing genes to further understand disease mechanisms.
Methods
ES cell culture
Experiments were performed using R1 mouse ES cells (purchased from ATCC, Rockville, MD). ES cells were seeded on mitomycin C-inhibited (10 μg/mL for 2.5 h; Sigma, St. Louis, MO) embryonic fibroblasts in Dulbecco's-modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA) containing 10% fetal bovine serum (Invitrogen), 2 mM L-glutamine, 100 units/mL penicillin, 100 μg/mL streptomycin, 1000 unit/mL recombinant mouse LIF (ESGRO; Chemicon, Temecula, CA), and 0.1 mM β-mercaptoethanol (Sigma) (ES medium).
ES cell differentiation
ES cells were seeded onto 0.1% gelatin-coated 100-mm dishes at a density of 2 × 106 cells/dish and were cultured for 2–3 days in ES medium for amplification. Cells were then seeded onto 100-mm Petri dishes in suspension for 2 days to allow for formation of embryoid bodies (EBs) using the same above-described medium without LIF. From days 3 to 6, EBs were treated with 2 μM all-trans retinoic acid (RA; Sigma). On day 4, various concentrations of Bmp4 were added to the culture medium. EBs were collected at day 7 of culture and transferred to fibronectin-coated (FN, R&D Systems, Minneapolis, MN) dishes with 80/20 medium (StemCell Technologies, BC, Canada) (80/20 supplemented with N2 and B27, Gibco, Grand Island, NY) for an additional 4 days. Cells were trypsinized, triturated through flame-polished Pasteur pipettes, and reseeded onto poly-L-ornithine/laminin (LPO, Sigma and Invitrogen) substrates for 4 days (Suppl. Fig. 1A). LPO coating included incubation in 140 μL/cm2 poly-L-ornithine solution (15 μg/mL; Sigma) at 37°C for 96 h, followed by washing in Ca2+/Mg2+-free PBS and in DMEM/F12 before cells were plated in the presence of 1 μg/mL laminin (Invitrogen).
RNA extraction and RT- PCR analysis
Total RNA was isolated using Trizol reagent (Invitrogen) and DNase I. Total RNA concentration was measured using an UV spectrophotometer (Amersham, Lake Forrest, IL). Isolated RNA was used to synthesize cDNA, according to instructions from the SuperScript III reverse transcriptase kit (Invitrogen). cDNA samples were normalized based on GAPDH as a reference gene. PCRs were performed using Taq polymerase (Invitrogen). Single bands of amplification products were confirmed in a 2% agarose gel (Invitrogen) containing ethidium bromide for visualization.
Immunocytochemical analysis
Cells were cultivated in a 24-well culture plate with LPO-coated coverslips, washed three times in phosphate-buffered saline (PBS), and fixed in 4% paraformaldehyde for 30 min at 4°C. To reduce nonspecific binding, cells were treated for 30 min with 10% blocking serum after washing three times for 5 min each with PBS. All primary antibodies were applied for 1 h in 10% blocking serum and PBS at room temperature. The following primary antibodies were used for labeling: anti-β-tubulin III (1:100, Chemicon), anti-GABA (1:100, Santa Cruz, Santa Cruz, CA), anti-Pax7 (1:100, Abcam, Cambridge, MA), anti-Mash1 (1:100, Abcam), anti-Math1 (1:100, Abcam), anti-Olig2 (1:100, Abcam), anti-GAD65 (1:100, Abcam), anti-Ngn2 (1:100, Abcam), anti-Nestin (1:100, Santa Cruz), anti-O4 (1:50, Santa Cruz), anti-MBP (1:50, Santa Cruz), anti-NFL (1:50, Santa Cruz), anti-AFP (1:50, Santa Cruz), anti-α-SMA (1:50, Santa Cruz), and anti-CHAT (1:100, Santa Cruz). Cells were then rinsed for 15 min in PBS, and the secondary antibody was administered for 1 h at room temperature. The secondary antibodies used were FITC (1:200, Santa Cruz) and TRITC (1:200, Santa Cruz) labeled. Cell nuclei were labeled for 30 sec with Hoechst 33258 (Sigma), and a laser-scanning confocal microscope was used to identify positively stained cells (Zeiss LSM 510; Zeiss, Thornwood, NY). Cell quantification was performed by counting cells from at least three independent experiments. At least 1000 total cells were evaluated per dish for antigen expression from 20 randomly chosen visual fields.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31000645), the National Natural Science Foundation of China (30900413), the Research Fund for the Doctoral Program of Higher Education of China (20092307120016), and the China Postdoctoral Foundation (20080430939). We thank Dr. Yu Li from Harbin Institute of Technology and Zhi-wei Pan from Harbin Medical University for technical assistance.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.