
Abstract
Abstract
Low efficiency of somatic cell nuclear transfer (SCNT) is attributed to incomplete reprogramming of transferred nuclei into oocytes. Trichostatin A (TSA), a histone deacetylase inhibitor, has been used to enhance nuclear reprogramming following SCNT. However, the molecular mechanism of TSA for the improvement of the preimplantation embryo and fetal development following SCNT is not known. The present study investigates embryo viability and gene expression of cloned bovine preimplantation embryos in the presence and absence of TSA compared to embryos produced by in vitro fertilization or parthenogenetic activation. Our results indicated that TSA treatment significantly improved total and inner cell mass (ICM) cell number and ratio of ICM:trophectoderm (TE) and also decreased the apoptotic index including total, ICM, and ratio of ICM:TE. Four apoptotic-related genes, Bcl-xL, survivin, Bcl2-associated X protein (Bax), and caspase 3 (Casp3), and four pluripotency/differentiation related genes, Oct4, SRY (sex determining region Y)-box 2 (Sox2), Cdx2, and colony-stimulating factor 1 receptor (Csf1r), were measured by real-time RT-PCR. TSA treatment resulted in the high expression of antiapoptotic gene Bcl-xL and low expression of pro-apoptotic gene Bax compared to untreated NT embryos, fertilized embryos, or parthenotes. Furthermore, mRNA expression of Cdx2 was higher in NT-TSA embryos than in NT and in vitro fertilization (IVF) counterparts. Additionally, low expression of microRNA (mir)-21 in NT embryos was enhanced following TSA treatment. These results suggest that TSA positively regulates nuclear reprogramming, and TSA may increased resistance or reduced signal for induction of apoptosis.
Introduction
For the successful development of cloned embryos, the donor cell nucleus must be epigenetically reprogrammed by the cytoplasm of the oocyte during early development. However, it is possible that cloned embryos may show abnormalities at the molecular level, including abnormal gene expression (Daniels et al., 2000; Inoue. et al., 2006, Li et al., 2005; Winger et al., 2000; Wrenzycki et al., 2001), abnormal X-chromosome inactivation (Yang et al., 2007), and abnormal genome imprinting (Yang et al., 2007). Moreover, the low success rate is believed to be associated with epigenetic errors, including abnormal histone modification. In early-stage bovine embryos, the pattern of histone H3 lysine 9 (H3K9) methylation closely mimics that of DNA methylation. At the blastocyst stage, both DNA and H3K9 were hypermethylated in the inner cell mass (ICM) but hypomethylated in the trophectoderm (TE).
Trichostatin A (TSA) is a well-known histone deacetylase inhibitor, which enhances the pool of acetylated histones (Yoshida et al., 1990) and DNA demethylation (Hattori et al., 2004). TSA can remarkably reduce abnormal DNA hypermethylation depending on the origins of transferred nuclei and their genomic regions (Kishigami et al., 2006b). Recent studies have shown that TSA has increased the rate of development to the blastocyst stage and improved blastocyst quality through induced hyperacetylation in cloned mouse (Kishigami et al., 2006b; Rybouchkin et al., 2006), bovine (Ding et al., 2008), porcine (Cervera et al., 2009; Zhang et al., 2007), and rabbit (Shi et al., 2008) embryos. In contrast, Iager et al. (2008) has reported that TSA does not affect the developmental rate in bovine somatic cell nuclear transfer (SCNT) embryos. However, it is not known how TSA facilitates nuclear reprogramming and influences the preimplantation embryo and fetal development following SCNT.
In NT embryos, the percentage of embryos with DNA-fragmented nuclei is apparently higher than in in vitro fertilization (IVF) embryos for cows (Fahrudin et al., 2002; Feugang et al., 2003) and pigs (Hao et al., 2003). During mouse embryonic development, multiple caspases and anti- and pro-apoptotic genes are expressed (Cohen, 1997; Cryns and Yuan, 1999; Salveson and Dixit, 1997), and in cows, anti- and pro-apoptotic genes such as Bcl2, Bcl-xL, and Bax are involved in regulation of apoptosis in preimplantation embryos (Augustin et al., 2003; Kolle et al., 2002; Lonergan et al., 2003; Yang et al., 2002). Survivin has been shown to directly inhibit the activity of the cysteine protease caspase-3 and likely acts to inhibit apoptosis by directly binding to caspases (Conway et al., 2000; Shin et al., 2001).
miRNAs (microRNA) have recently been discovered as a class of small RNAs that regulate gene expression epigenetically at the posttranscriptional or translational level. miRNAs are expressed in a tissue-specific manner and are thought to play important roles in cell proliferation, apoptosis, and differentiation during mammalian development (Ambros, 2004; Sempere et al., 2004). Several apoptosis-related miRNAs are involved in apoptosis. For example, miR-15, miR-16, and mir-34 induce apoptosis (Cimmino et al., 2005; Hermeking, 2009; Yamakuchi et al., 2008), and miRNA-21 is an oncogenic miRNA exhibiting antiapoptotic activity in various carcinoma cell lines (Chan et al., 2005; Meng et al., 2006). Recently, we reported that developmentally important miRNAs, such as mir-127, are aberrantly expressed in cloned mouse embryos (Cui et al., 2009). To our knowledge, bovine embryonic apoptosis and its relation to miRNA expression, especially in embryos derived by NT, have not been reported previously.
The objectives of this research were to investigate the effect of TSA on bovine NT embryo development, total and ICM cell number, and apoptosis as compared to untreated NT embryos, fertilized embryos, or parthenotes. Furthermore, we quantified the expression of apoptotic-related genes, including mRNAs, miRNAs and pluripotency/differentiation-related genes, in these embryos.
Materials and Methods
All chemicals were purchased from Sigma Chemical Company (St. Louis, MO) unless otherwise noted.
Maturation and preparation of oocytes
Bovine ovaries were collected at the abattoir, washed several times with fresh saline, and transported in sterile PBS at 33°C to the laboratory within 3 h of collection. Cumulus oocyte complexes (COCs) were aspirated from 2–7-mm follicles, washed, and selected morphologically for in vitro maturation. Groups of 30–40 COCs were incubated in TCM 199 (Gibco, Grand Island, NY) supplemented with 10% (v/v) fetal bovine serum (FBS) (Gibco), 1 μg/mL FSH (Stimufol; Merial, Lyon, France), 0.2 mM sodium pyruvate, 1% antibiotic, and 1 μg/mL estradiol-17β for 19–24 h at 39°C in a humidified atmosphere of 5% CO2 in air. At the end of the maturation period, oocytes from the same batch were used either to provide a recipient cytoplast for cloning, for parthenogenetic activation, or for IVF.
IVF
IVF was performed according to the standard procedures previously described (Rosenkrans et al., 1993) with some modification. Following maturation, COCs were placed in fertilization microdrops of Fert-TALP without glucose (Parrish et al., 1988) containing 4 IU/mL heparin and PHE (20 μM penicillamine, 10 μM hypotaurine, 1 μM epinephrine). The frozen–thawed semen was thawed, and then motile spermatozoa were obtained for fertilization by using a gradient Percoll (Pharmacia, diluted into 45% and 90%). The viable sperm pellet was recovered and resuspended, and sperm concentration was determined by using a hemocytometer. Sperm were produced at 1 × 106 spermatozoa/mL at the final concentration. Spermatozoa and maturated oocytes were coincubated for approximately 9 h. Following coculture, zygotes were washed and cultured in CR1aa (Rosenkrans et al., 1993) medium. The next day, the cumulus cells were removed and culturing was continued in CR1aa.
Donor ear fibroblast culture and preparation
Ear fibroblast cells were cultured from the ear biopsy of an adult female Holstein. The ear biopsy was washed several times in phosphate-buffered saline (PBS) and diced into 1-mm pieces. Explants were transferred into a 100-mm tissue culture dish containing Dulbecco's modified Eagle's medium (DMEM)+10% FBS and then cultured in the incubator under the same conditions as for in vitro maturation. When confluence was achieved after approximately 10 days in culture, cells were trypsinized, and the resulting pellet was resuspended in the above culture medium and subcultured until use.
Enucleation
Between 19 and 21 h after IVM, surrounding cumulus cells were removed by vortexing in the presence of 2 mg/mL hyaluronidase, and oocytes with an extruded first polar body were selected for enucleation. Prior to enucleation, oocytes were incubated for 20 min in 4 μg/mL demicolcine diluted in IVM medium. After demecolcine treatment, each group of oocytes was washed three times in demicolcine-free medium and transferred for enucleation into a manipulation medium drop containing 10% FBS overlaid with warm mineral oil. The zona pellucidae were partially dissected with a fine glass needle, and a small amount of cytoplasm containing metaphase chromosomes was pushed out by squeezing the oocyte with a holding pipette and glass needle.
Donor cell injection and activation of reconstructed embryos
Donor cell injection was performed using direct injection methods (Kishigami et al., 2006c). The donor membrane was removed from cells by gentle aspiration in and out of the injection pipette until each nucleus was clearly separate from any visible cytoplasmic material. The donor nucleus was injected using a piezoelectric device (Prime Tech, Ibaraki, Japan). Embryos were activated by 5 μM ionomycin calcium salt for 5 min and then cultured in CR1aa (Rosenkrans et al., 1993) medium with 2.0 mM 6-(dimethylamino)purune (6-DMAP) for 4 h at 39°C in a humidified atmosphere of 5% CO2 in air.
TSA treatment
For TSA treatment, chemically activated reconstructed oocytes were cultured in CR1aa medium containing both 2.0 mM 6-DMAP and 0, 50, 500, and 1000 nM TSA for 4 h and then further cultured in CR1aa medium containing 50 nM TSA for another 6 or 16 h at 39°C in a humidified atmosphere of 5% CO2 in air.
Parthenogenetic activation
For parthenogenetic activation, denuded oocytes were exposed to 5 μM ionomycin calcium salt for 5 min and then cultured in CR1aa medium with 2.0 mM 6-DMAP for 4 h (Campbell et al., 2000) at 39°C in a humidified atmosphere of 5% CO2 in air.
Embryo culture
Fertilized and activated embryos were thoroughly washed in CR1aa medium containing 0.3% BSA, transferred into 30-μL droplets (13–15 embryos/droplet) of medium under mineral oil, and cultured for 3 days. After 3 days, eight-cell stage embryos were collected and then cultured in CR1aa medium containing 10% FBS for another 4 days at 39°C in a humidified atmosphere of 5% CO2 in air.
Immunofluorescence and quantification of signal intensity
To determine the expression and distribution of proteins, bovine embryos were fixed with 4% formaldehyde for 20 min, and after fixation, they were permeabilized with 0.2% Triton X-100 for 10 min. The embryos were incubated with anti-di-methyl histone H3 lysine 9 (H3-m2K9, Cell Signaling) and antihistone H3 acetyl lysine 9 (H3-aceK9, Cambridge, Abcam, UK) for 1 h and then with FITC-labeled secondary antibodies. Hoechst 33343 was used to stain the nuclei. Slides were examined by laser-scanning confocal microscopy using a Leica DM IRB equipped with a krypton–argon ion laser for the simultaneous excitation of fluorescence for proteins and chromatin. The identity of the slides was concealed, and histone H3-m2K9- and H3-aceK9-specific green fluorescence and UV intensity for chromatin was scored using MetaMorph soft ware (Version 6.1; Universal Imaging Corporation) (Cui et al., 2006) in each embryo, and about 15 embryos were used in each experiment. The chromatin of untreated embryos was used as a control in each embryo, and the intensity level in the IVF embryo was arbitrarily set to onefold.
Cell number detection and TUNEL assay
Bovine blastocysts were fixed with 3.7% formaldehyde for 30 min at room temperature and then permeabilized with 0.3% Triton X-100 in 0.1% PVA-PBS for 1 h under the same conditions. The numbers of ICM and TE in blastocysts were examined for the presence or absence of ICM-specific Oct4 (Santa Cruz, Santa Crua, CA, USA) protein (Palmieri et al., 1994). Terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) was used to determine the proportion of cells undergoing apoptosis. The total cell number of blastocysts was measured using the fluorochrome nuclear stain bisbenzimide Hoechst No.33342. The apoptotic index was calculated by the percentage of nuclei displaying apoptotic morphology concurrent with TUNEL labeling, that is, the percentage of apoptotic nuclei number/total nuclei number. The blastocysts were 60, 68, 61, and 62 in IVF, parthenote, NT, and NT-TSA groups, respectively, and performed four separate experiments.
Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
Messenger RNA was extracted with the Dynabeads mRNA Direct Kit (Dynal Asa, Oslo, Norway), according to the manufacturer's instructions. Initially, standard cDNA synthesis was achieved by reverse transcription of RNA using the Oligo (dT)12-18 primer and superscript reverse transcriptase (Invitrogen Co., Grand Island, NY).
Real-time RT-PCR was performed using the nine primer sets shown in Table 1 with DNA Engine OPTICOJ 3 (MJ Research, Waltham, MA, USA) (Cui et al., 2006). The relative quantification of gene expression was conducted using the 2-ΔΔCt method (Livak and Schmittgen, 2001). In all experiments, glyceraldehyde-3-phosphate dehydrogenase (Gapdh) mRNA was employed as an internal standard for the analysis of relative transcript levels of each gene in various embryos. Ten embryos were used in each experiment and performed three separate experiments with three replicates.
TaqMan RT-PCR
All primers, including for specific miRNA cDNA synthesis and for PCR amplification, and the kit used in miRNA analysis was purchased from Applied Biosystems (Bedford, MA, USA).
For relative quantification of miRNA-15, -16, -21, and -34 expression, reverse transcriptase (RT) reactions contained 5-μL RNA samples, 3 μL of stem–loop RT primer, 1 × RT buffer (P/N: 4319981), 0.25 mM each of dNTPs, 3.33 U/μL MultiScribe reverse transcriptase (P/N: 4319983), and 0.25 U/μL RNase inhibitor (P/N: N8080119). The 15-μL reactions were incubated for 30 min at 16°C, 30 min at 42°C, 5 min at 85°C, and then held at 4°C to convert miRNA into cDNA. Real-time PCR was performed on a 20-μL volume and included 4 μL of RT product, 1 × TaqMan Universal PCR Master Mix (P/N: 4324018), and 1 μL of 20 × real-time solution containing TaqMan probe and primers. The amplification parameters for real-time PCR were employed according to the manufacturer's protocol. The relative quantification of miRNA expression was analyzed using the same method as for mRNA expression (Livak and Schmittgen, 2001). Because no internal control miRNA has been used in bovine studies, Gapdh mRNA was employed as an internal reference.
Statistical Analysis
The general linear models (GLM) procedure in the SAS program (SAS, 1985) was used to analyze the data from all experiments. Significant differences were determined using Tukey's multiple range test (Steel and Torrie, 1980), and p < 0.05 was considered significant.
Results
Effect of TSA on in vitro preimplantation embryo development
As a preliminary experiment, NT embryos treated with 50 nM TSA for 10 h (TSA 50–10 h) or for 20 h (TSA 50–20 h), 500 nM for 10 h (TSA 500–10 h), and 1000 nM for 10 h (TSA 1000–10 h) postactivation were cultured and compared with untreated (TSA 0) embryos (Fig. 1A). Concentrations of 500 and 1000 nM TSA significantly inhibited embryo development; however, no differences were observed in TSA 50–10 h and TSA 50–20 h group embryos when compared to untreated embryos (TSA 0; described below as NT). We therefore chose 50 nM for 10 h (described below as NT-TSA) as our TSA treatment experiment in the present study and compared it to IVF, parthenote, NT, and NT-TSA.
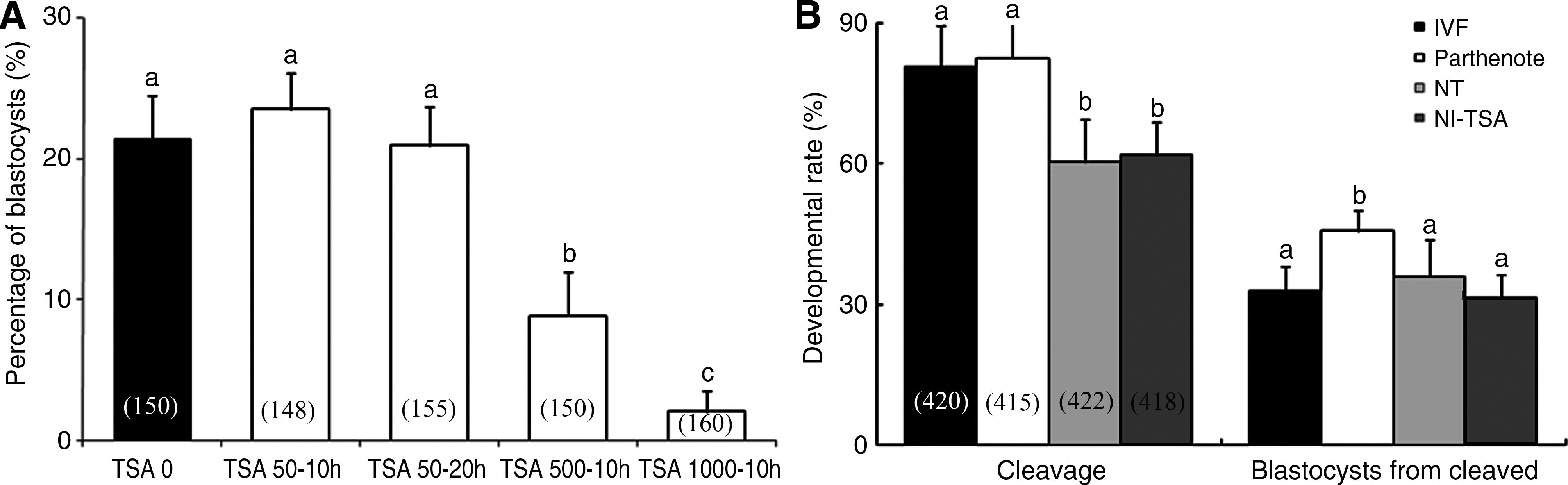
The percentage of embryos to develop to blastocysts after treatment with various concentrations of TSA (
The developmental rates to the cleavage and day 7 (168 h postactivation) blastocyst stages of embryos were determined from eight replicates in four treatment groups (Fig. 1B). Development from the one- to two-cell stage is often impeded by the failure of pronuclear formation and by improper cell-cycle stages of donor cells in cloned (NT and NT-TSA) embryos (Kishigami et al., 2006a). Then, we calculated the percentage of blastocysts from two-cell-stage embryos. TSA treatment did not influence NT embryo (NT-TSA) development to the blastocyst from the two-cell stage when compared to untreated control NT embryos. There were also no differences compared to IVF embryos, with rates ranging from 31 to 36%. However, the percentage of blastocysts was significantly higher in the parthenotes group (47%) than in other groups.
Effect of TSA on methylation and acetylation of histone H3K9
Fluorescence-stained two-cell and blastocyst stage embryos were used to assess the levels of acetylation and methylation of histone H3K9 (Fig. 2). Histone H3-m2K9- and H3-aceK9-specific green fluorescence intensity and UV intensity for chromatin were scored using MetaMorph soft ware (Version 6.1; Universal Imaging Corporation, Buckinghamshire, UK) (Cui et al., 2006). To normalize the high density of small nuclei, the intensity of chromatin (UV) was used as a control in each embryo, and to describe the relative intensity, the level of the IVF embryo was arbitrarily set to onefold. Experiments were repeated three times independently, and about 15 embryos were used in each experiment. Overall, treatment with 50 nM TSA for 10 h did not affect acetylation and methlylation of H3K9 in two-cell (Fig. 2A and D) and blastocyst (Fig. 2B and E) stage embryos (NT-TSA) compared to IVF, parthenote, and NT embryos (Fig. 2). However, 1 μM TSA caused hyperacetylation in two-cell stage NT embryos (NT-TSA 1 μM) when compared to those of the other three groups (Fig. 2C and F).
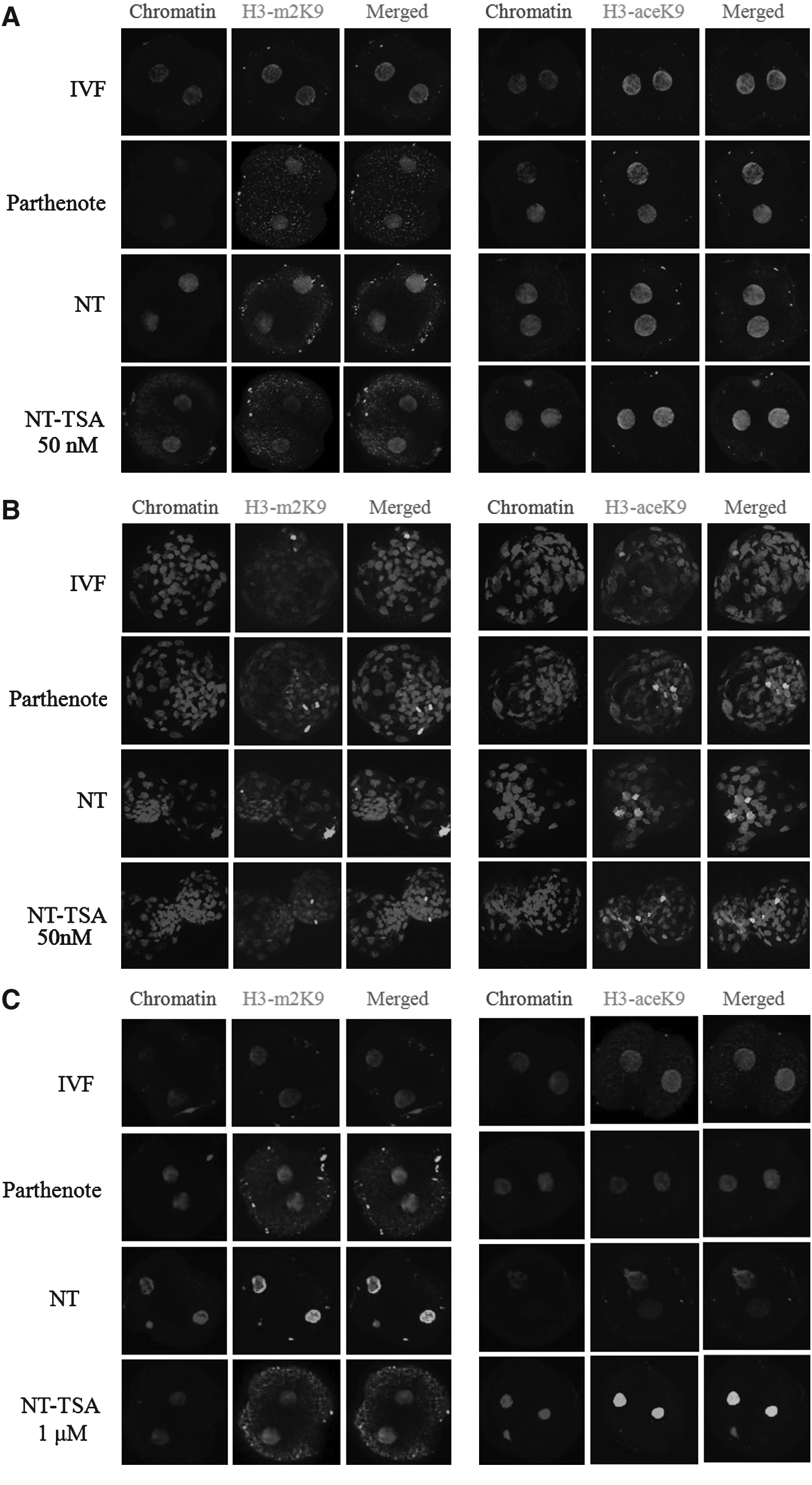
Laser scanning confocal microscopy images of methylation (left) and acetylation (right) of histone H3K9 in two-cell (treatment of NT embryos with 50 nM (
Effect of TSA on cell number and apoptosis
The mean total and ICM cell number and ratio of ICM:TE in blastocysts reconstructed with bovine ear cells and those without TSA treatment were not statistically different compared with the parthenote and IVF groups, with a number ranging from 76 to 81 (total cell; Fig. 3A) and 20 to 25 (ICM cell, Fig. 3B). However, TSA treatment (NT-TSA) significantly increased the total (22.3%) and ICM (47.6%) cell number (total cell 101 and ICM cells 31, respectively) and the ratio of ICM:TE compared to the non-TSA-treated control NT group, higher than those of the IVF counterpart (Fig. 3A). Additionally, the NT group showed a high apoptotic index (i.e., percentage of nuclei displaying apoptotic morphology concurrent with TUNEL labeling), including in total (Fig. 3D) and ICM (Fig. 3E) cells compared to in IVF and parthenote embryos. Fortunately, addition of TSA to the culture medium resulted in a significant reduction of apoptosis in total (Fig. 3D) and ICM (Fig. 3E) cells compared to non-TSA-treated cultures.
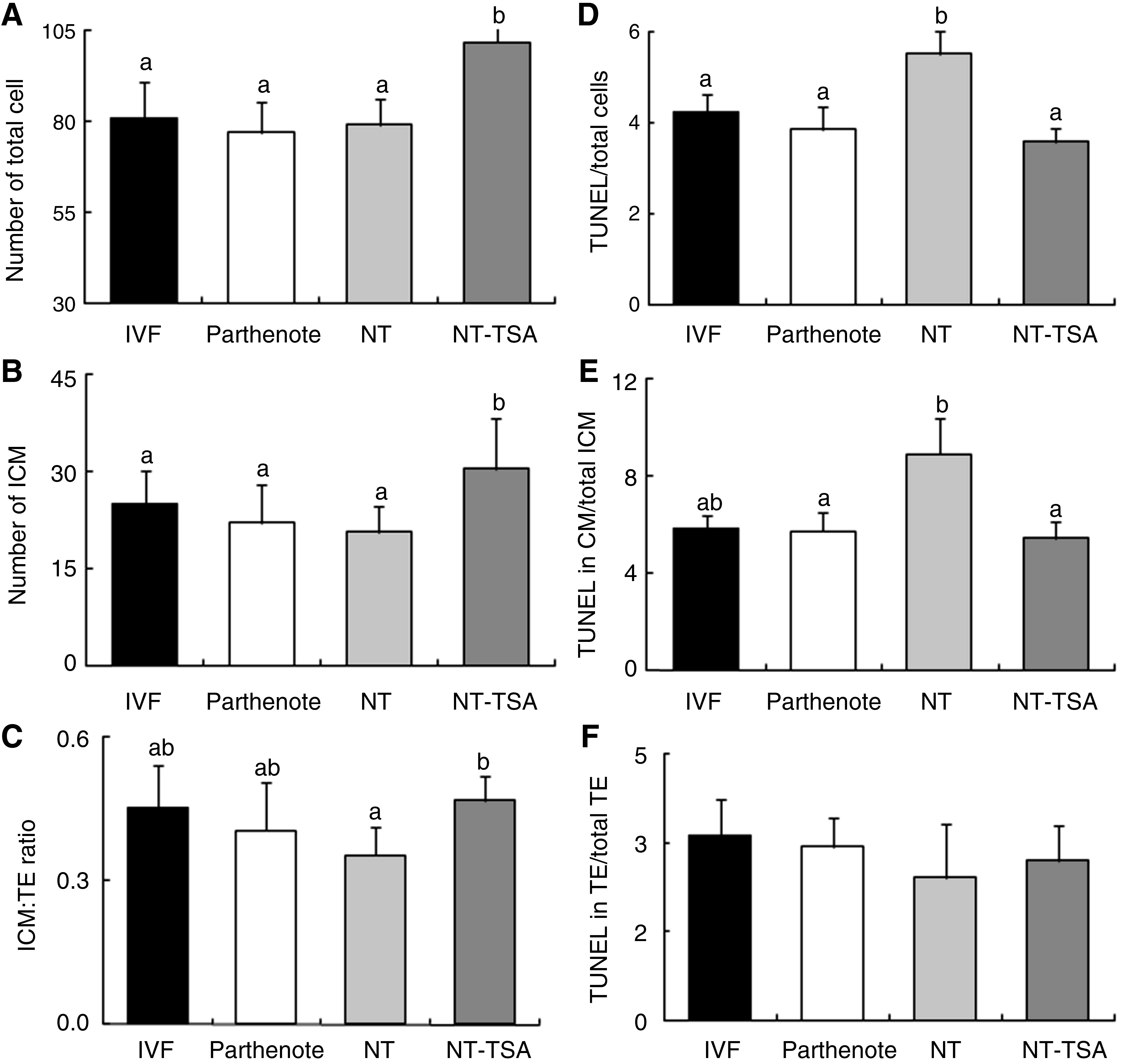
Total cell (
Effect of TSA on mRNA and microRNA expression
Messenger RNA expression of apoptotic-related and ICM- or TE-related genes was observed by real-time RT-PCR. Experiments were replicated four times independently with three observations for each time. In all experiments, Gapdh was used as the internal reference for normalization of relative mRNA expression. Expression levels of survivin, Oct4, and Csfr1 were not statistically different among the groups (Fig. 4A and B). TSA treatment increased Cdx2 expression when compared with non-TSA-treated control NT embryos, and Sox2 and Cdx2 expression was higher in parthenotes than in IVF embryos (Fig. 4A). Antiapoptotic gene Bcl-xL mRNA expression was higher and pro-apoptotic gene Bax mRNA expression was lower in the NT-TSA group than in the other three groups. Another pro-apoptotic gene, Casp3, exhibited lower expression in parthenotes compared to other groups (Fig. 4B).
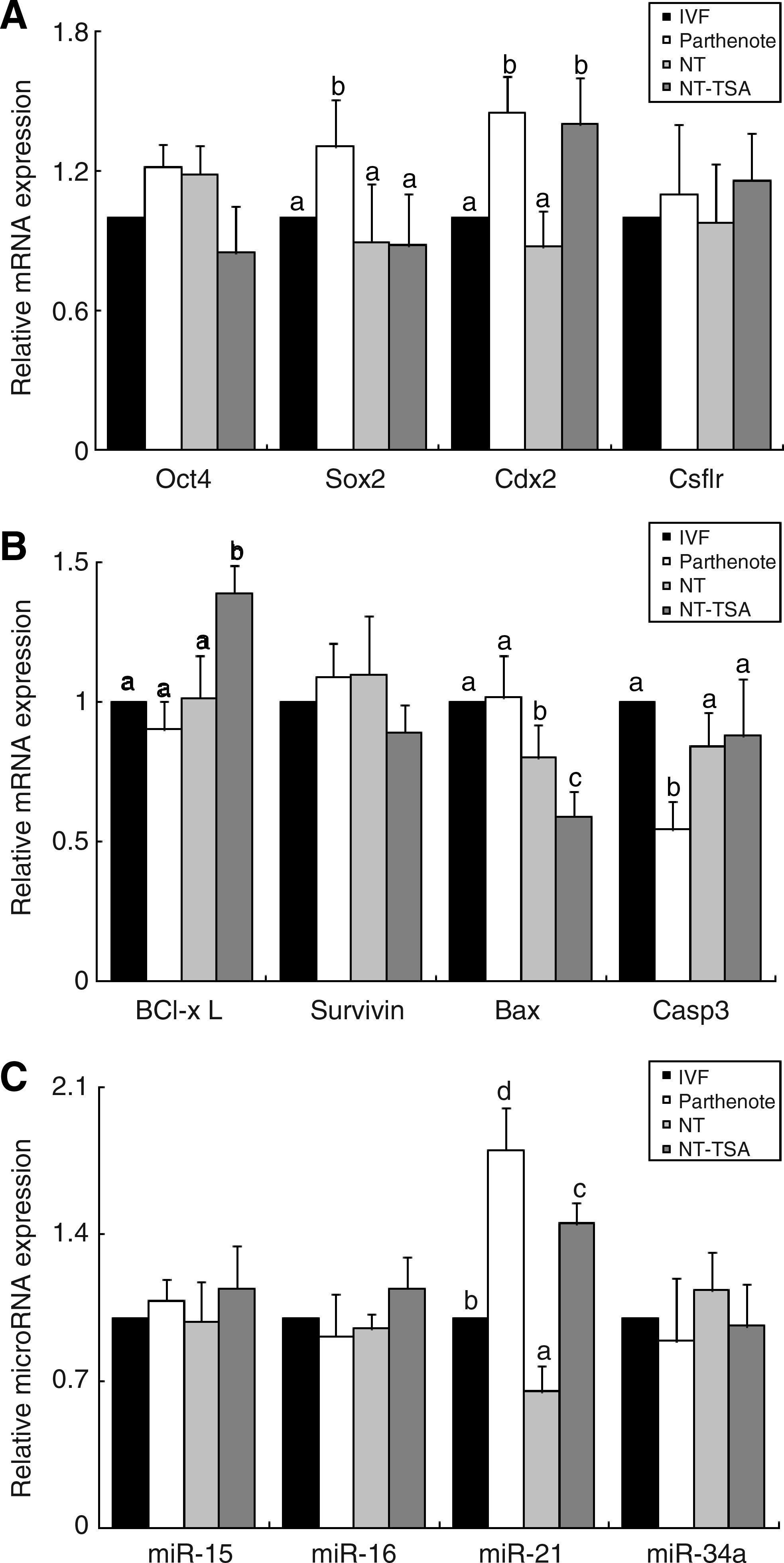
Real-time RT-PCR determination of the relative expression levels of pluripotency or differentiation (
TaqMan real-time RT-PCR relative quantification analysis of four different miRNAs was performed on five blastocysts in each separate experiment. Because the use of miRNA as an internal control has not yet been reported in bovine studies, Gapdh mRNA expression was used as the internal control and miRNA expression of the IVF group was arbitrarily set to onefold (Fig. 4C). There was no difference in miRNA expression of mir-15, −16, and −34a among the group. mir-21 expression was significantly lower in the NT group than in the other groups, but was enhanced by TSA treatment compared to non-TSA-treated embryos. Additionally, mir-21 was expressed at greater levels in the embryos that were derived from parthenogenetic activation.
Discussion
Reduced histone acetylation was observed in the cloned bovine embryos (Iager et al. 2008; Wee et al., 2006), which could cause an aberrant transcription of cloned embryos and fetuses. Therefore, the histone deacetylase inhibitor TSA has been suggested to be useful for recovery of aberrant nuclear reprogramming following nuclear transfer. In fact, TSA increases the rate of development to the blastocyst stage and results in improved blastocyst quality through induced hyperacetylation in cloned mouse (Kishigami et al., 2006b; Rybouchkin et al., 2006), bovine (Iager et al., 2008), porcine (Cervera et al., 2009; Zhang et al., 2007), and rabbit (Shi et al., 2008) embryos. In the present study, we treated cloned bovine embryos with 0, 50, 500, and 1000 nM TSA, consistent with previous reports (Enright et al., 2003; Kishigami et al., 2006a). High concentrations (500 or 1000 nM) of TSA were toxic to the embryo development (Fig. 1A) even though high concentration (1000 nM TSA) hyperacetylated two-cell stage NT embryos (Fig. 2C and F). Results suggest that treatment of NT embryos with sufficient TSA can modify their histone acetylating status. Treatment with 50 nM TSA for 10 h did not influence the percentage of cleavage and development to blastocysts from the two-cell stage, when compared to control NT or the fertilized (IVF) counterpart (Fig. 1B), a result consistent with a previous study (Iager et al., 2008). The blastocyst formation rate is not necessarily indicative of a positive developmental capacity in bovine clones (Rybouchkin et al., 2006). Cloned blastocyst formation rates can reach nearly 80% and sometimes surpass IVF rates, while the proportion of developmentally competent embryos is greater following IVF (Cibelli et al., 2006).
One criterion for evaluation of blastocyst quality is the cell number, especially that of the ratio of ICM:TE cells (Van Soom et al., 1996; Yu et al., 2007). Placental abnormalities or early fetal loss might be due to aberrant allocation of cells in NT embryos to ICM and TE cells at preimplantation stages (Im et al., 2006). The structural integrity of the blastocyst composed of ICM and TE cells might influence development after implantation in mammals (Koo et al., 2002, 2004). We therefore measured the total and ICM cell number and the ratio of ICM:TE in the four treatment groups. Interestingly, TSA treatment increased total cell (22.3%) and ICM (47.6%) cell number compared to the non-TSA-treated NT group (Fig. 3). This result is consistent with an earlier study: the mean total and TE cell numbers were increased by 25% and the ICM cell numbers were increased by 50% in TSA-treated embryos compared to untreated NT embryos (Rybouchkin et al., 2006). This can be explained by improved reprogramming in the TSA group or otherwise more effective DNA replication because prevention of histone deacetylation stimulates some cells (Aoki and Schultz, 1999; Vogelauer et al., 2002).
Apoptosis is another criterion of quality to eliminate cells with nuclear or chromosomal abnormalities (Hardy, 1997). Apoptotic cell death occurs in preimplantation mammalian embryos and plays an important role in embryo development. Apoptosis has been observed in bovine embryos after the eight-cell stage (Fahrudin et al., 2002; Matwee et al., 2000). The increase of apoptosis in NT blastocysts is also correlated with a decrease in the total cell number (Yu et al., 2007). In the present study, NT embryos showed a higher apoptotic index than that of the IVF or parthenote group (Fig. 3). This observation is similar to that reported previously, where Fahrudin et al. (2002) found that higher apoptotic morphology and/or TUNEL labeling were detected in NT embryos when compared with bovine IVF embryos. Especially, DNA fragmentation may play a role in the regulation of the ICM cell number, which is likely to be important, as an abnormal ICM cell number could result in abnormal development of the fetus (Fahrudin et al., 2002). Our data show that TSA protected NT embryos from high apoptosis especially in ICM cells (Fig. 3). A previous study showed that histone deacetylase inhibitors suppressed apoptosis in rat kidney cells (Dong et al., 2009), which indicates that TSA protects ICM cells from apoptosis, at least partially. Importantly, TSA improves full term development in the mouse (Kishigami et al., 2006a).
In the present study, TSA treatment not only had some positive effect on the proportion of nuclei able to achieve preimplantation development with low apoptosis especially in ICM cells but also had a positive influence on the reprogramming of genes including apoptotic- and pluripotency/differentiation-related gene expression. The propensity of a cell to undergo apoptosis is continuously counterbalanced by genes that stimulate cell survival and proliferation. During embryogenesis, the Bcl-2 family controls programmed cell death. The Bcl-2 gene family is known to include antiapoptotic and pro-apoptotic subgroups, and the Bcl2l1 (Bcl-xL) gene functions to protect against apoptosis (Boise et al., 1993; Shen et al., 2006). In contrast, another group of highly conserved genes are positive regulators of apoptosis, for example, the Bak and Bax proteins lead to the release of cytochrome c and the activation of the caspase cascade (Nutt et al., 2005). TSA affects mRNA expression in cloned mouse (Li et al., 2008; Shao et al., 2009) and bovine (Iager et al., 2008) blastocysts. In the present study, we chose to evaluate the expression of several developmentally important genes in blastocysts. Four ICM pluripotency or TE proliferation and/or differentiation genes Oct4, Sox2, Cdx2 (Iagel et al., 2008), and Cfs1r (Regenstreif and Rossant, 1989) were selected. The expression of four apoptosis-related genes was measured: Bcl-xL, Bax, Casp3, and survivin. Finally, expression of the four apoptotic-related miRNA-15, -16, -21, and -34 was determined. In agreement with previous studies, we found that expression of apoptotic-related genes changed according to TSA treatment (Fig. 4B). High expression of anti-apoptotic gene Bcl-xL and low expression of pro-apoptotic gene Bax may contribute to reduce apoptosis of total and ICM cells in NT-TSA blastocysts compared to those in NT embryos. This may be crucial, as a reduction of apoptosis during preimplantation development may be beneficial for later development. Interestingly, Cdx2, the earliest known marker of the TE lineage, was highly expressed in TSA-treated NT embryos compared to that in the control NT and IVF counterparts (Fig. 4A), perhaps due to a high proportion of cell number, including TE, in NT-TSA blastocysts (Iager et al., 2008).
We first employed the TaqMan real-time RT-PCR method to test microRNA expression and its relation to apoptosis, especially in embryos derived by NT (Fig. 4C). Our previous report (Cui et al., 2009) indicated that developmentally important microRNAs, such as mir-127, are aberrantly expressed in cloned mouse embryos. miRNA-21 has been described as an oncogenic miRNA that exhibits antiapoptotic activity in various carcinomas (Chan et al., 2005; Meng et al., 2006; Si et al., 2007). Here, we found that the level of antiapoptotic miRNA-21 was lower in NT embryos than in other groups and was upregulated uniquely after TSA treatment in cloned bovine embryos, whereas there were no changes to levels of pro-apoptotic microRNAs, such as mir-15, -16, and -34.
Most previous studies about embryo apoptosis evaluated the effect of culture conditions on fertilized, parthenogenetic, or nuclear transfer embryos (Brison and Schultz, 1997; Fabian et al., 2004; Herrler et al., 1998; Kawamura et al., 2003; Park et al., 2004a, 2004b). In the current study, as a control, another counterpart was parthenogenetically activated embryos. Delayed development and high apoptosis were observed in parthenote-derived haploidy embryos in the mouse (Cui et al., 2007) and pig (Hao et al., 2004). In contrast, there was no difference in the percentage of diploidy parthenotes to develop into blastocysts and in total cell number when compared to in vivo fertilized embryos in the mouse (Cui et al., 2007). Similarly, development of porcine parthenogenetic diploids to the blastocyst stage appeared normal (Kure-Bayashi et al., 1996). In the present study, the developmental rate was higher in putative diploidy parthenotes treated with 6-DMAP than in IVF embryos, possibly because polyspermy caused failure of development up to the blastocyst stage in the IVF group. Additionally, high Sox2 and Cdx2 mRNA expression and high mir-21 expression were detected in parthenotes compared to that in IVF embryos. This observation may reflect the ability of mRNA/miRNA to support embryo quality at the molecular level in preimplantation embryos.
In conclusion, in this study, we demonstrated that histone deacetylase inhibitor TSA improved the viability of cloned bovine somatic cell embryos by increasing cell numbers and may increased resistance or reduced signal for induction of apoptosis. The antiapoptotic effect of TSA seemed to be positively linked to antiapoptotic mRNA Bcl-xL and antiapoptotic miRNA-21 expression. However, evaluation of development to full term is necessary to ascertain whether this effect reflects a true enhancement in developmental potential.
Footnotes
Acknowledgments
This work is supported by Grant No. 109029-03-2-CG000 from the Ministry of Agricultural and Forestry, Republic of Korea.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.